
The Protective Role of Carbon Monoxide (CO) Produced by Heme Oxygenases and Derived from the CO-Releasing Molecule CORM-2 in the Pathogenesis of Stress-Induced Gastric Lesions: Evidence for Non-Involvement of Nitric Oxide (NO)


 , ,
, , 
Abstract
:1. Introduction
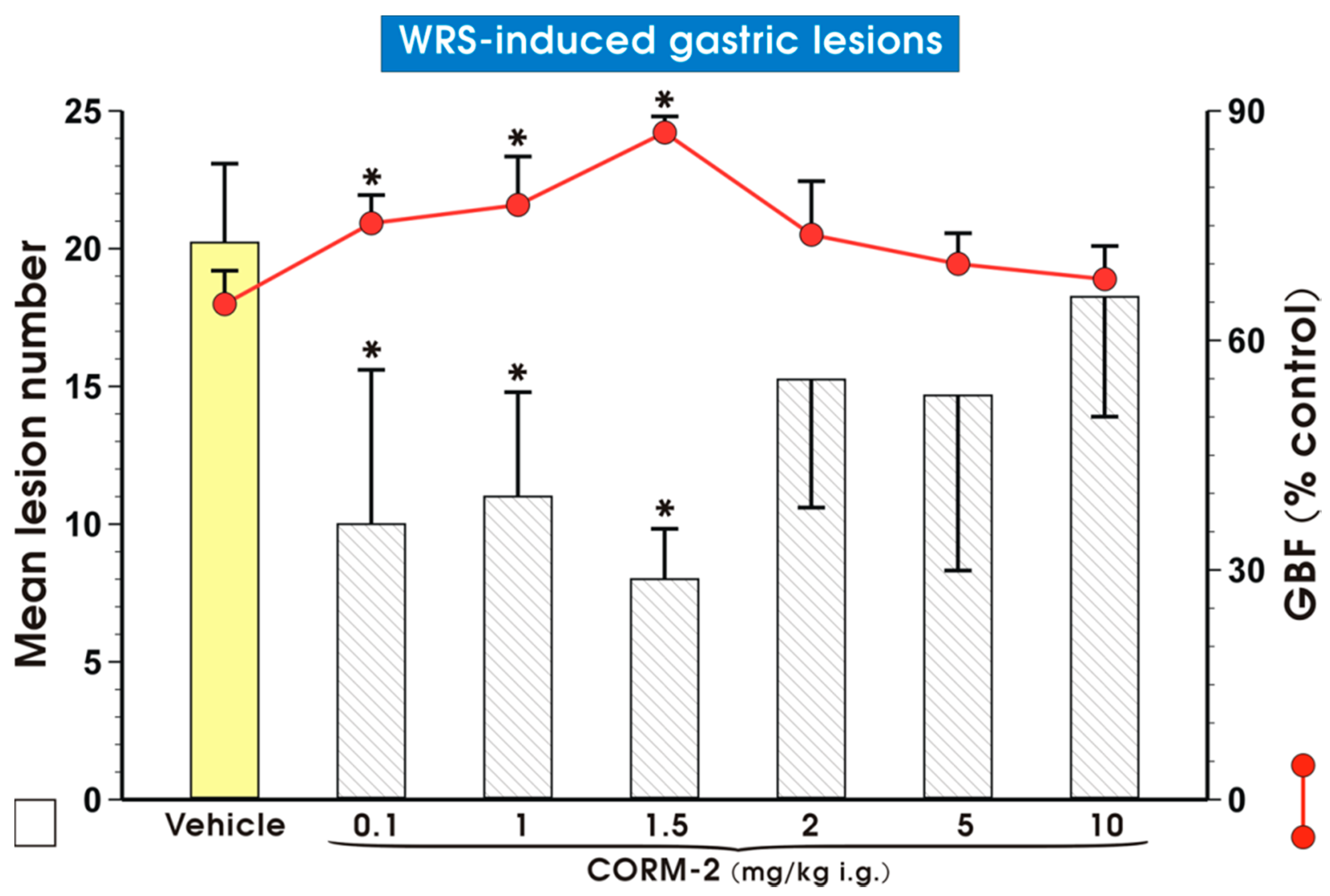
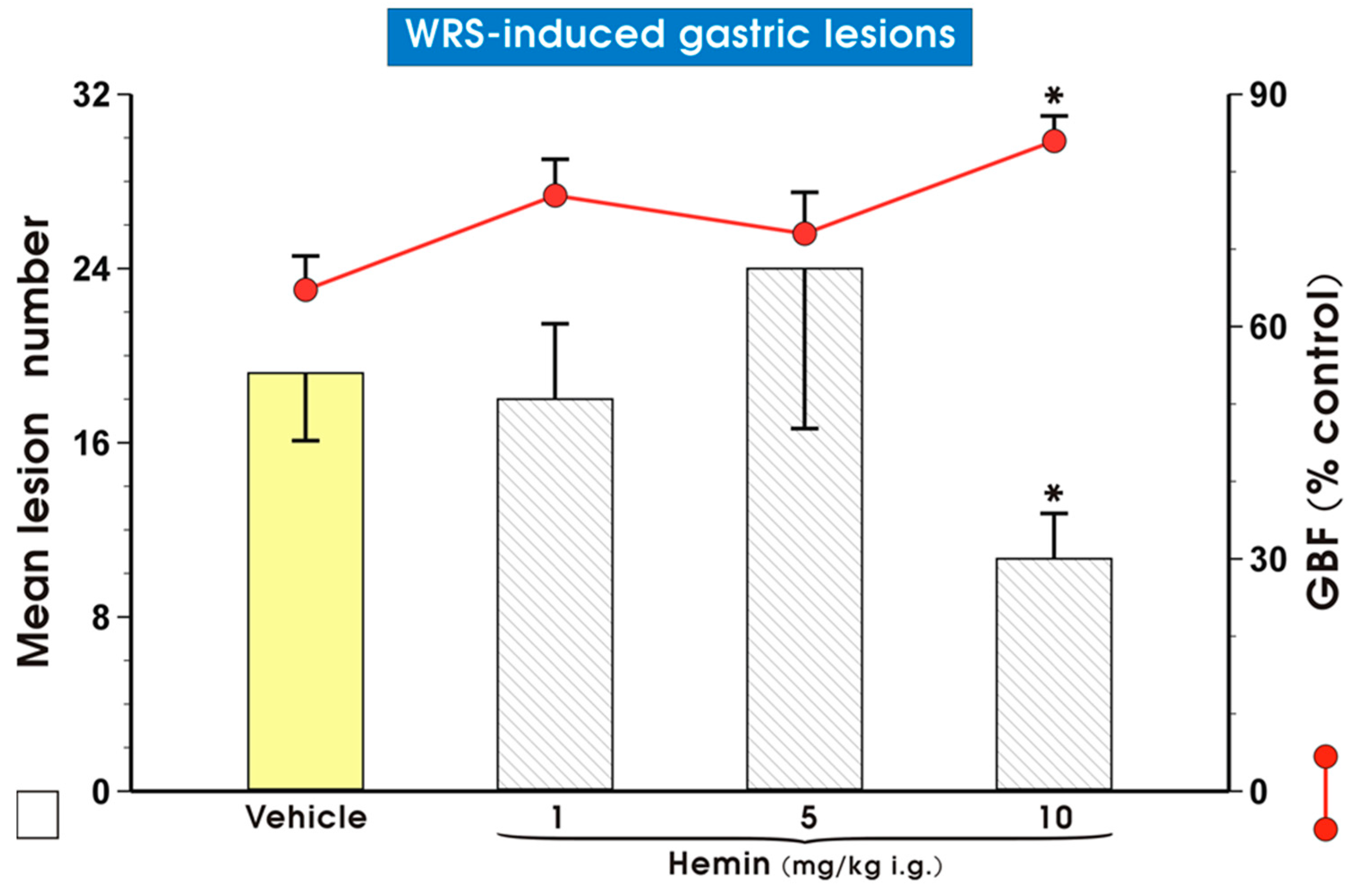
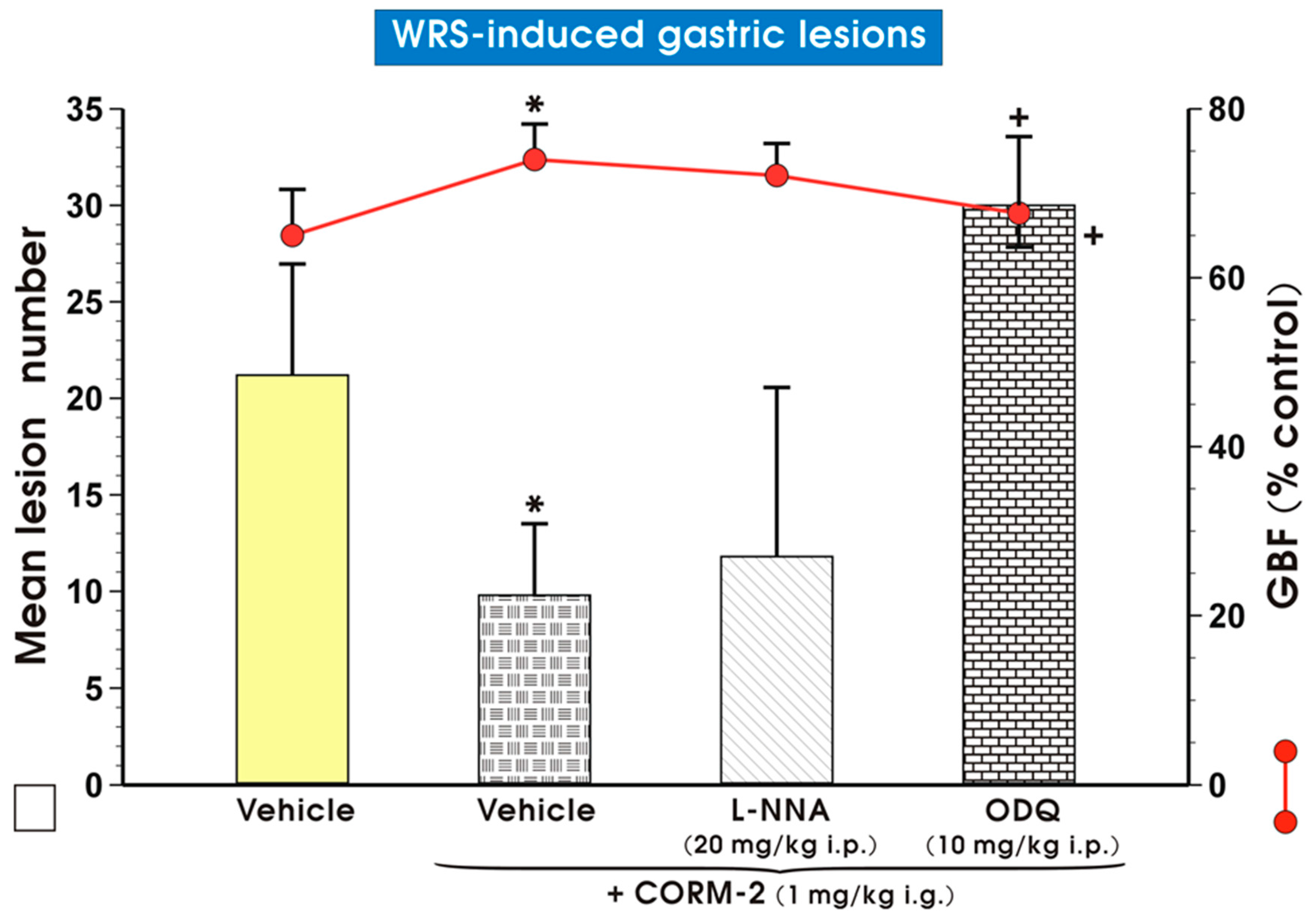
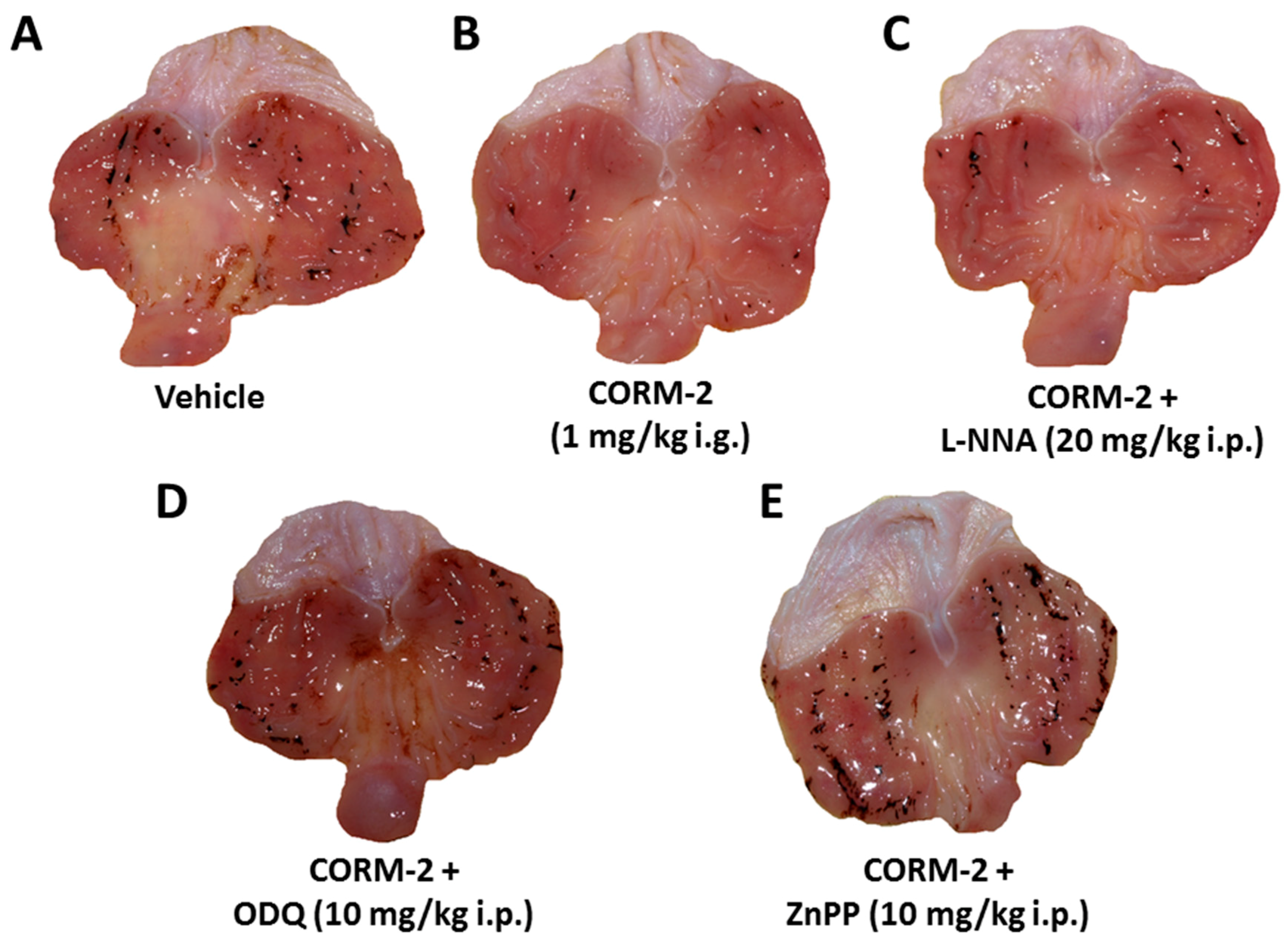
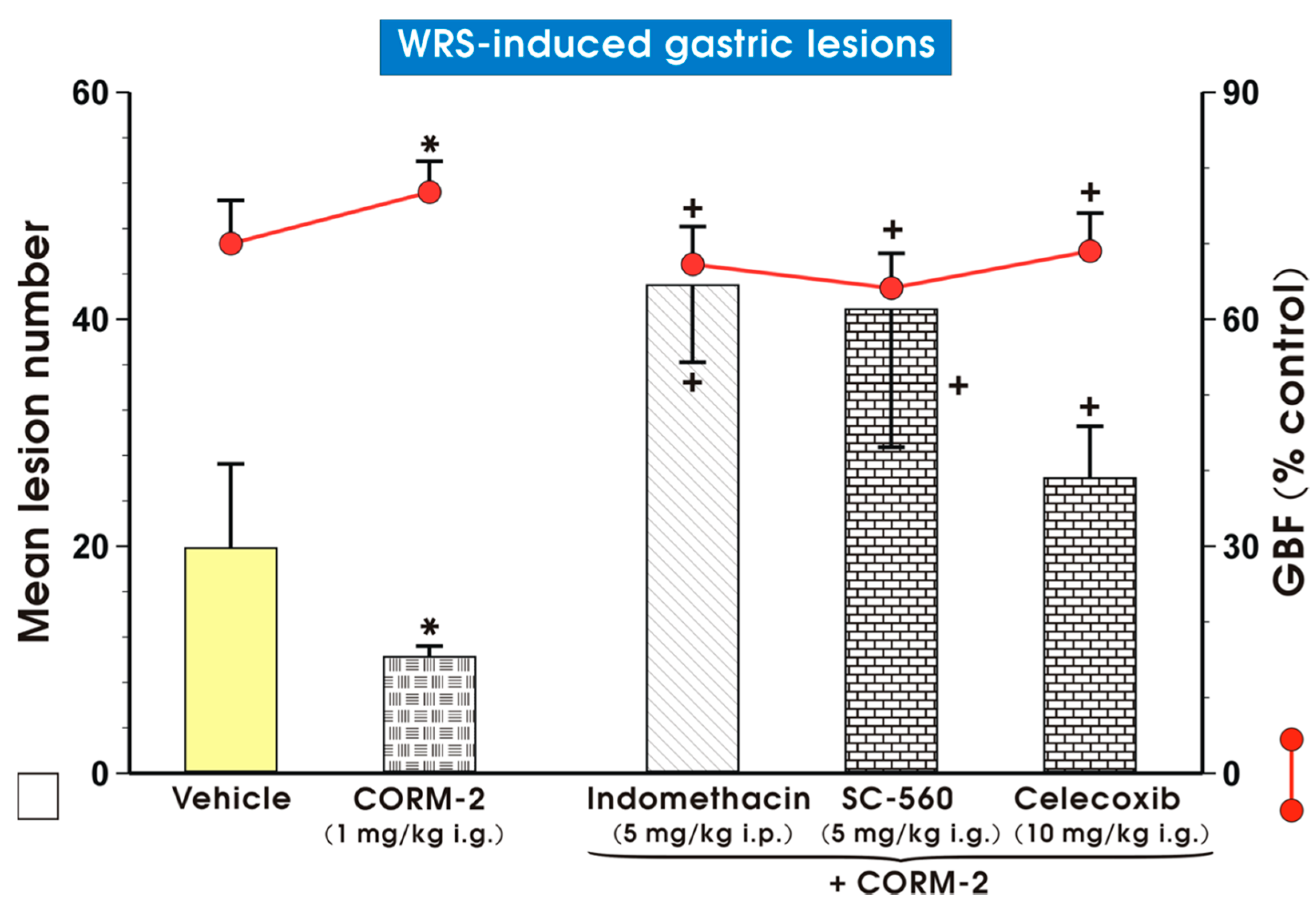
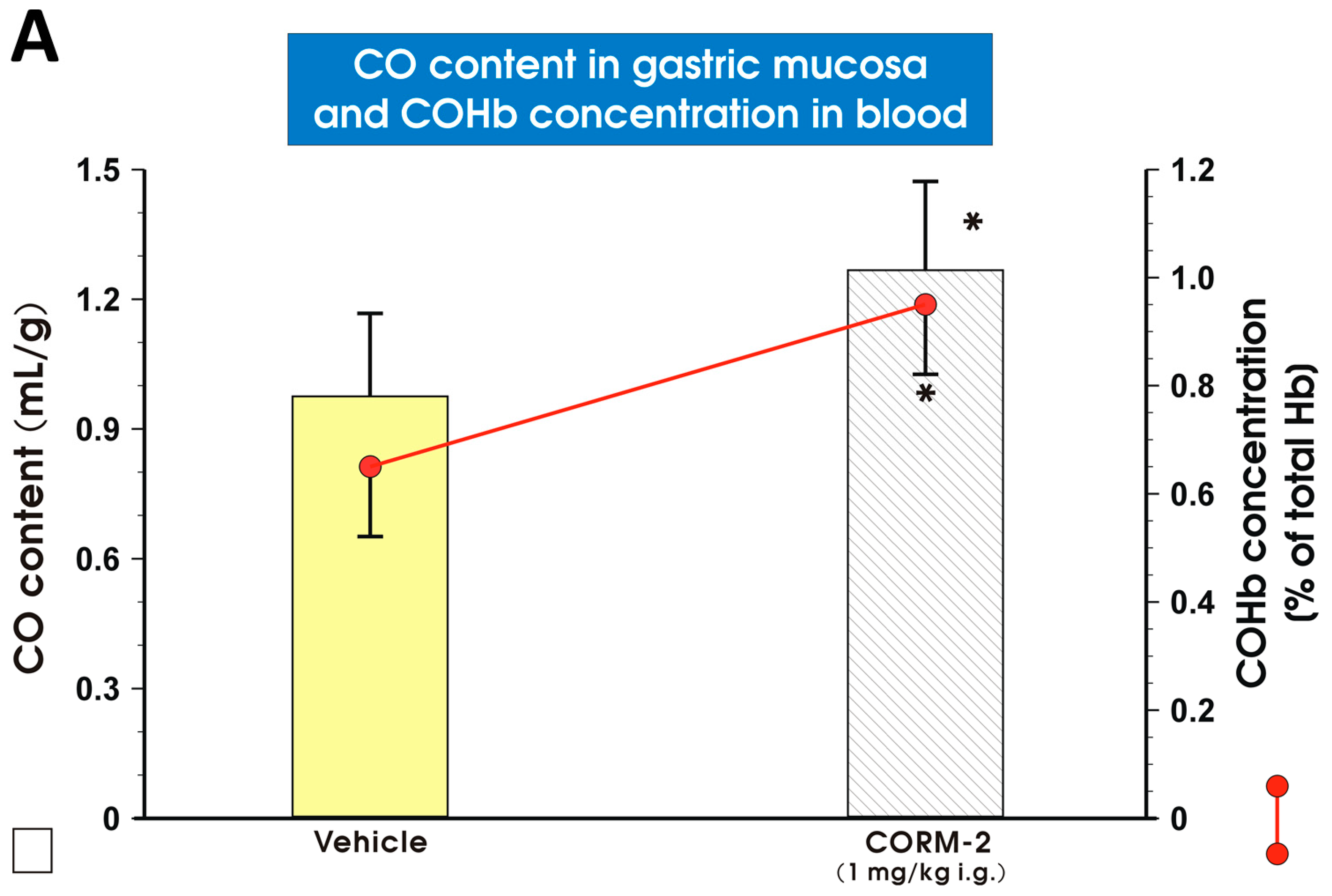
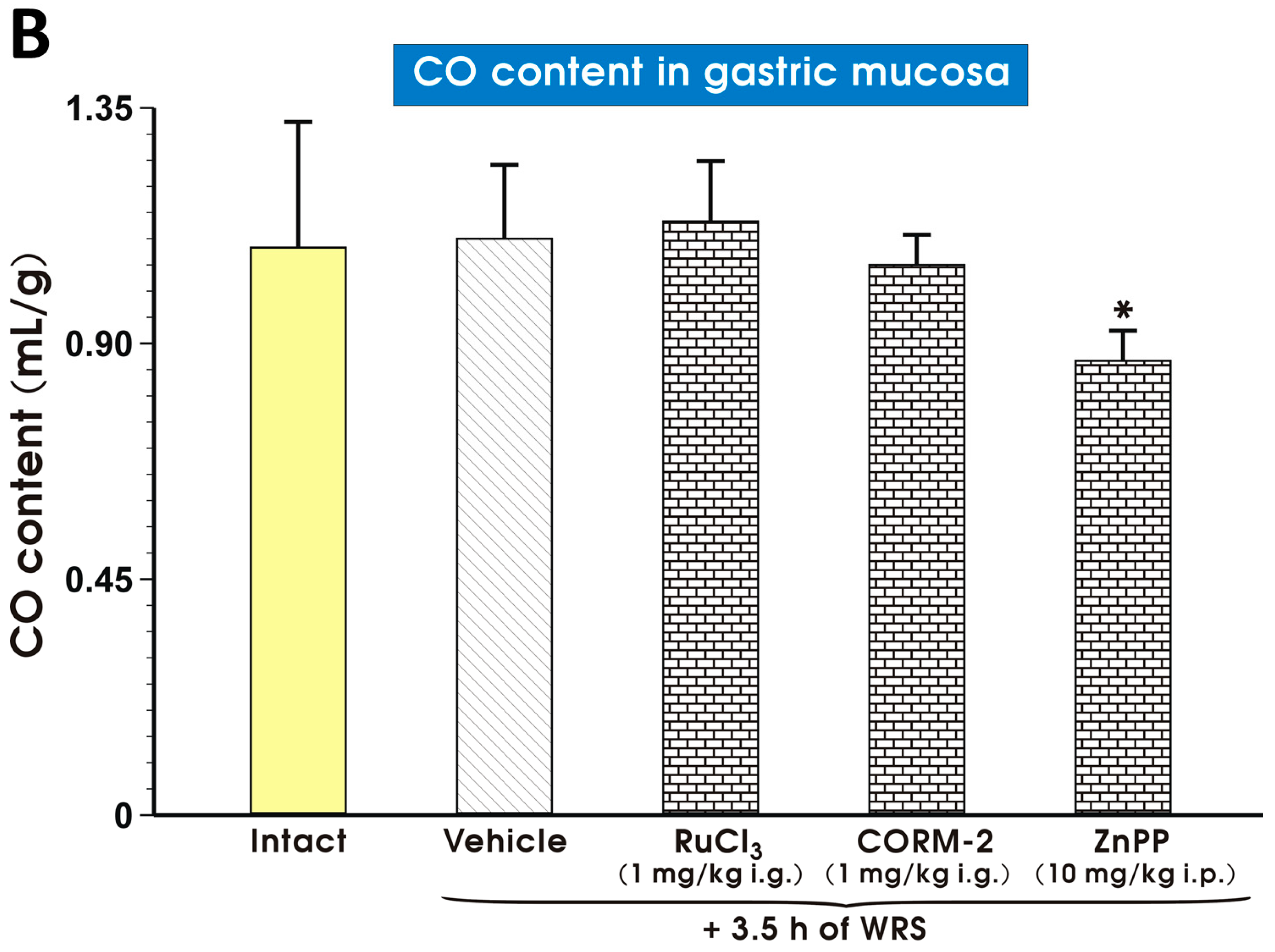
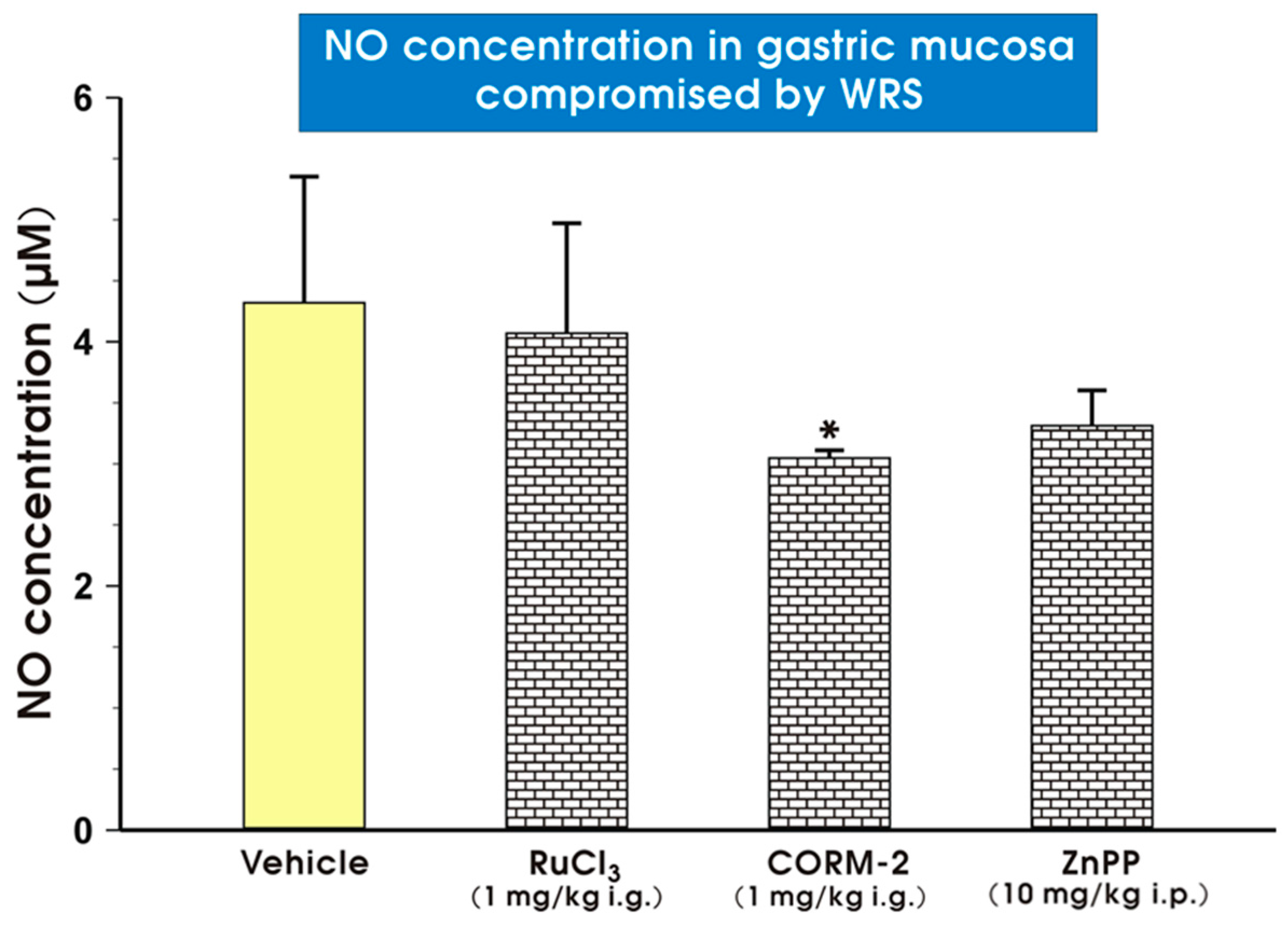
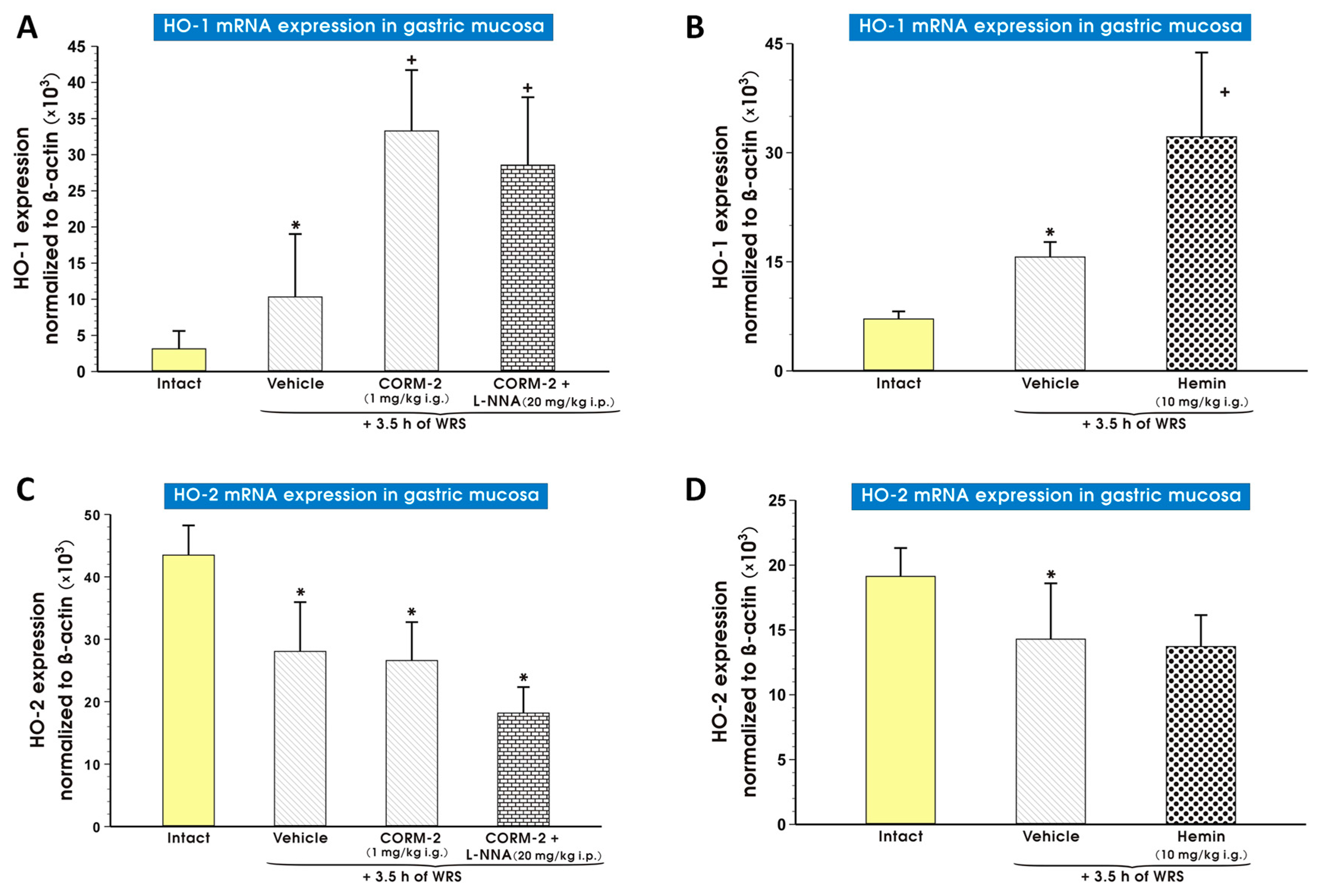
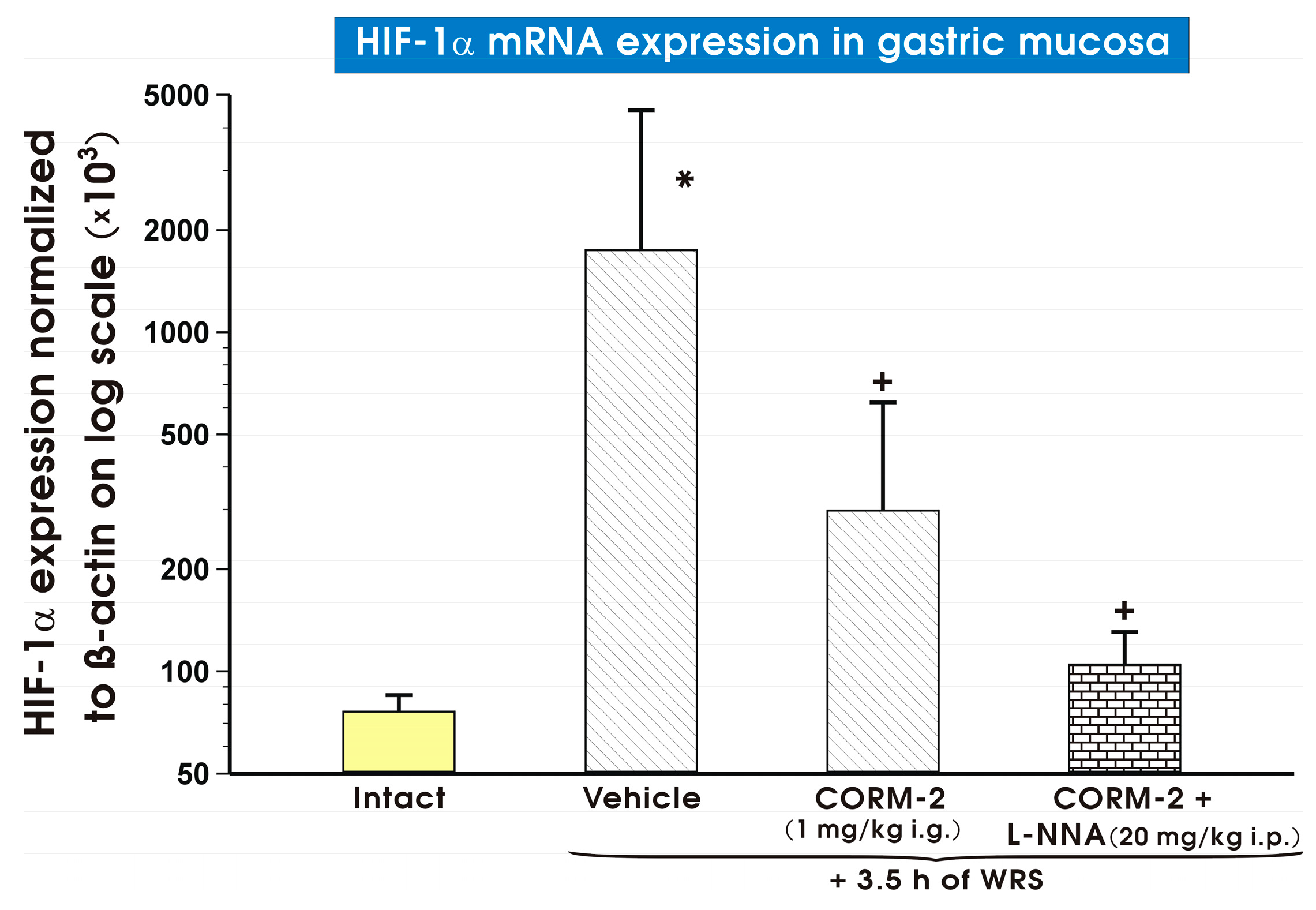
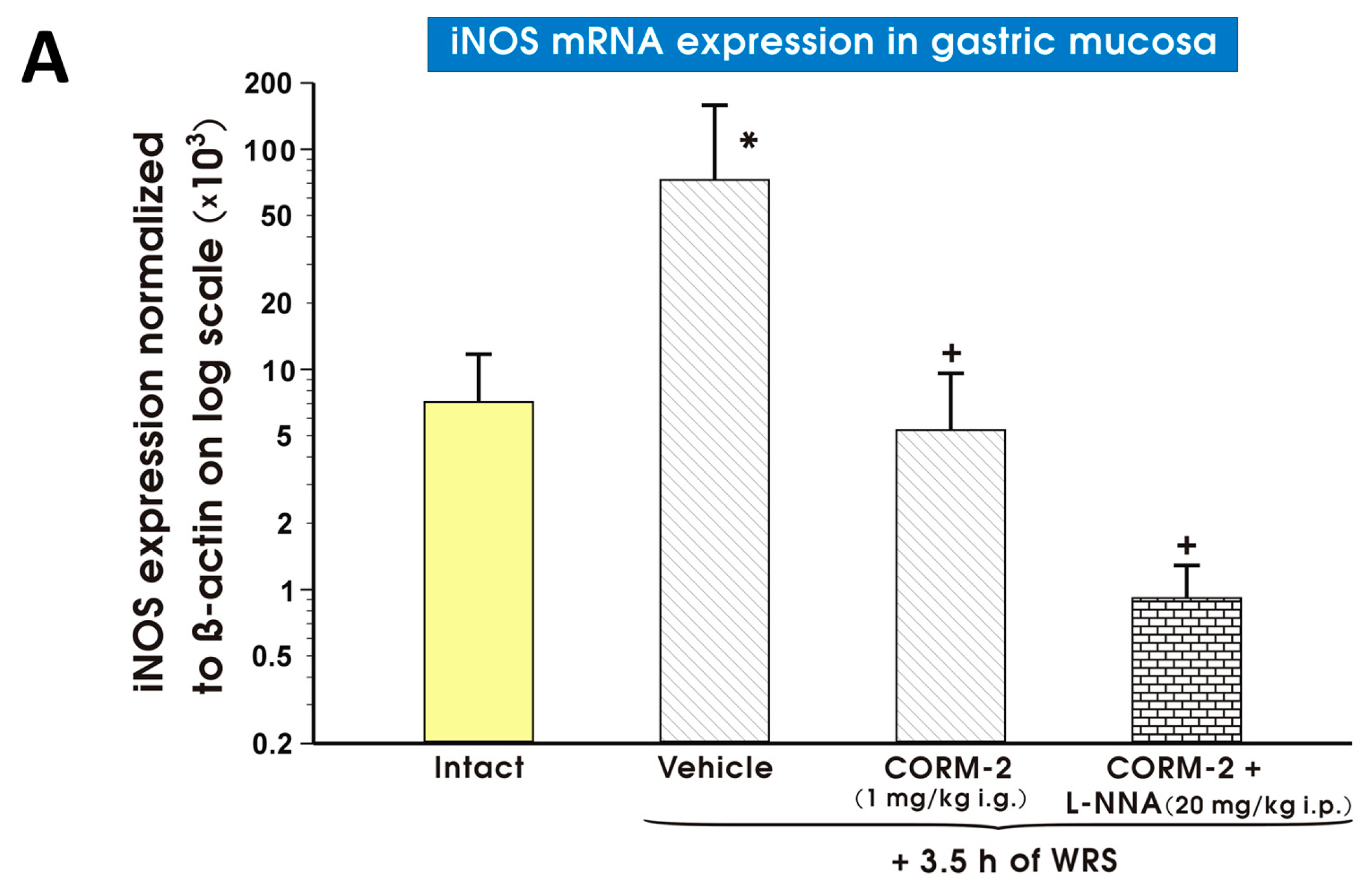
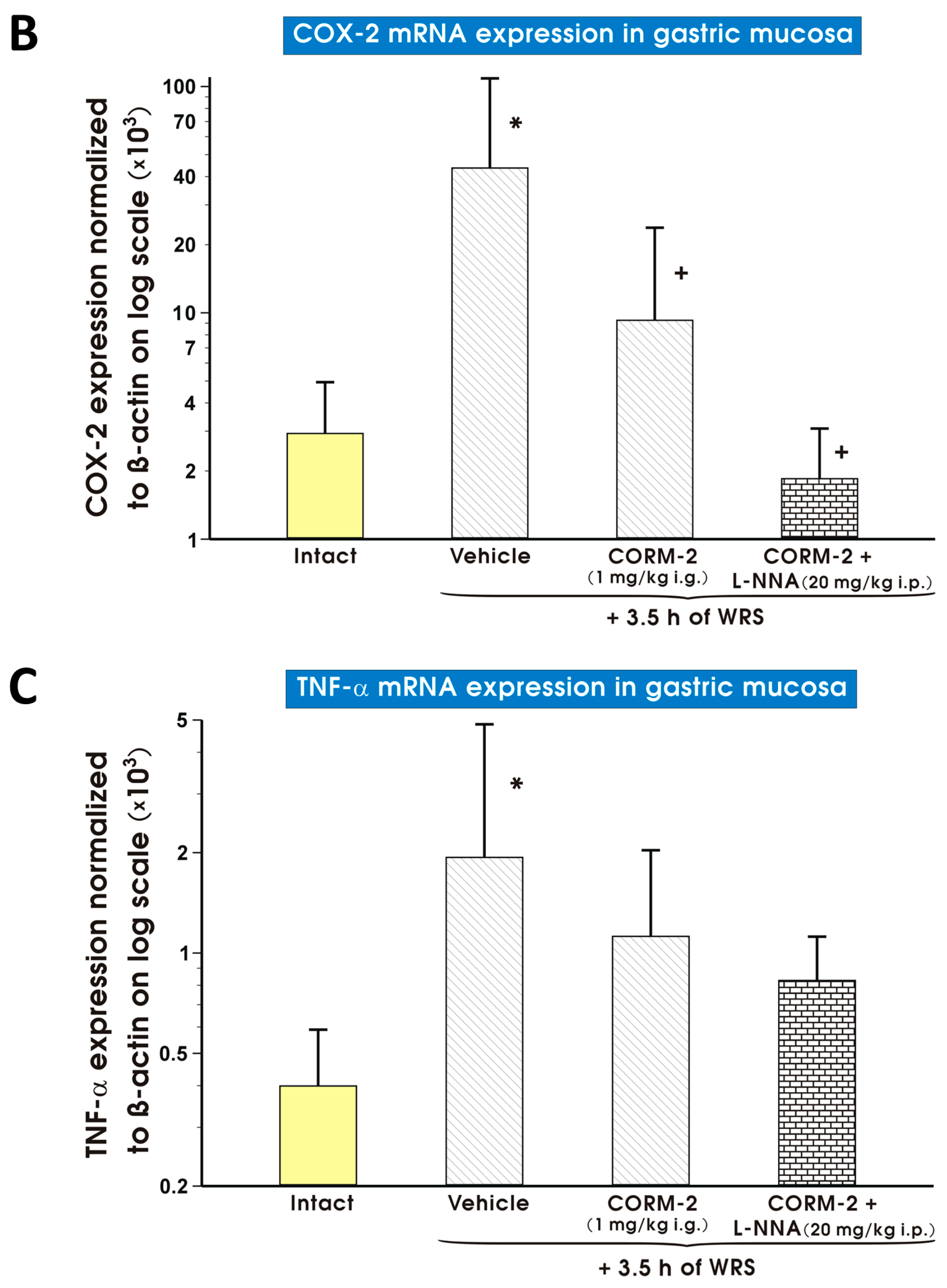
2. Results
3. Discussion
4. Materials and Methods
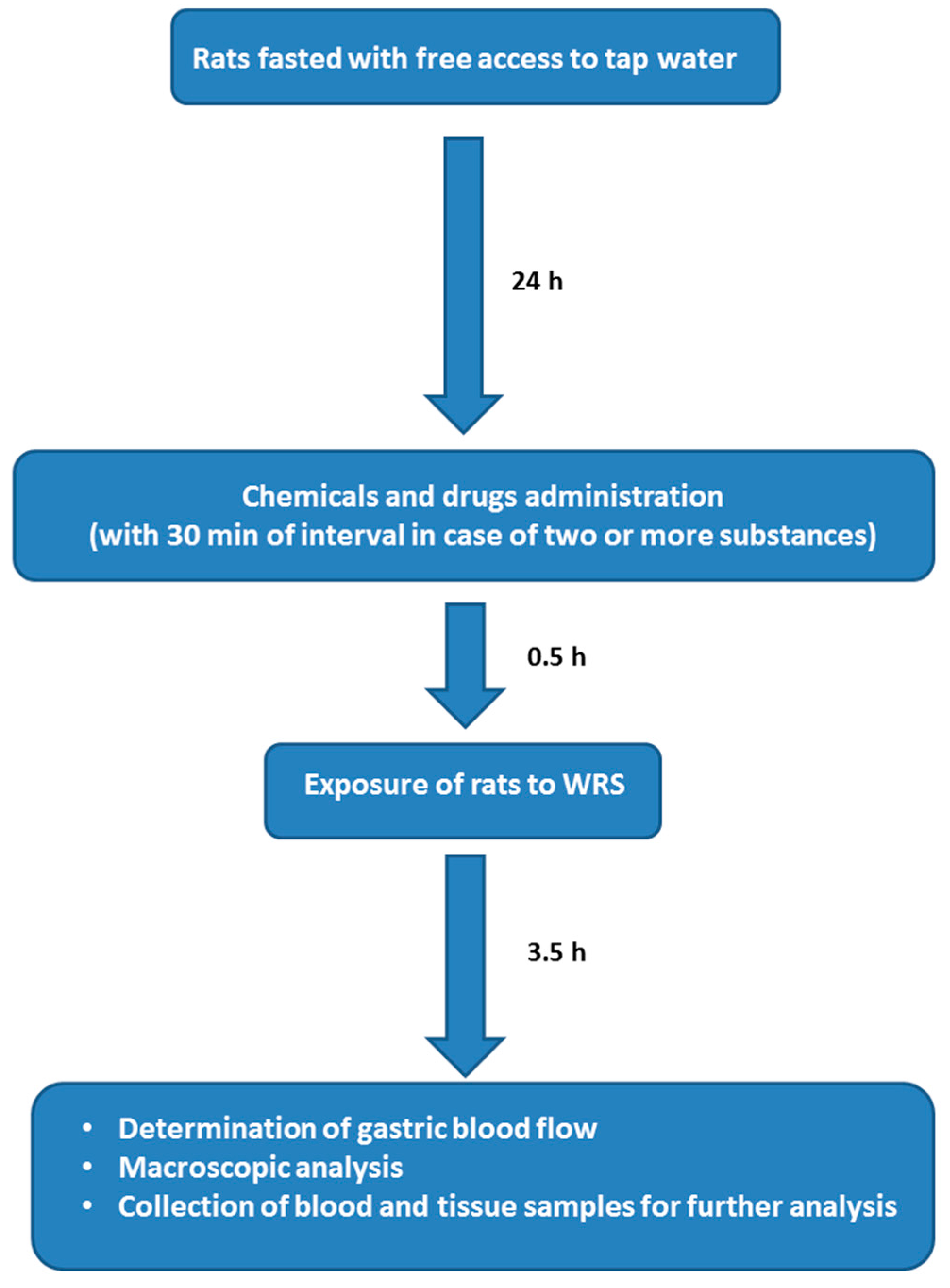
4.1. Animals, Experimental Design, Chemicals and Drugs Treatment
4.2. Determination of Gastric Blood Flow (GBF) and Gastric Lesion Number
4.3. Measurement of Carbon Monoxide (CO) Content in Gastric Mucosa and Blood Carboxyhemoglobin (COHb) Concentration Using Gas Chromatography (GC)
4.4. Determination of Nitric Oxide (NO) Content in Gastric Mucosa
4.5. Determination of mRNA Expression Levels for Heme Oxygenase (HO)-1, HO-2, Hypoxia Inducible Factor 1α (HIF-1α), Cyclooxygenase (COX)-2, Inducible NO Synthase (iNOS) and Tumor Necrosis Factor α (TNF-α) in Gastric Mucosa
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Morse, D.; Choi, A.M. Heme oxygenase-1: The “emerging molecule” has arrived. Am. J. Respir. Cell Mol. Biol. 2002, 27, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Alam, J.; Choi, A.M. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef] [PubMed]
- Tenhunen, R.; Marver, H.S.; Schmid, R. Microsomal heme oxygenase. Characterization of the enzyme. J. Biol. Chem. 1969, 244, 6388–6394. [Google Scholar] [PubMed]
- Rodgers, K.R. Heme-based sensors in biological systems. Curr. Opin. Chem. Biol. 1999, 3, 158–167. [Google Scholar] [CrossRef]
- Volpe, J.A.; O’Toole, M.C.; Caughey, W.S. Quantitative infrared spectroscopy of CO complexes of cytochrome c oxidase, hemoglobin and myoglobin: Evidence for one CO per heme. Biochem. Biophys. Res. Commun. 1975, 62, 48–53. [Google Scholar] [CrossRef]
- Sher, E.A.; Shaklai, M.; Shaklai, N. Carbon monoxide promotes respiratory hemoproteins iron reduction using peroxides as electron donors. PLoS ONE 2012, 7, e33039. [Google Scholar] [CrossRef] [PubMed]
- Caughey, W.S. Carbon monoxide bonding in hemeproteins. Ann. N. Y. Acad. Sci. 1970, 174, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Blomberg, L.M.; Blomberg, M.R.; Siegbahn, P.E. A theoretical study on the binding of O2, NO and CO to heme proteins. J. Inorg. Biochem. 2005, 99, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, R. Carbon monoxide: Endogenous production, physiological functions, and pharmacological applications. Pharmacol. Rev. 2005, 57, 585–630. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, G.; Lam, F.; Hagen, T.; Moncada, S. Inhibition of cellular respiration by endogenously produced carbon monoxide. J. Cell Sci. 2006, 119, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Omura, T.; Sato, R. The carbon monoxide-binding pigment of liver microsomes. I. Evidence for its hemoprotein nature. J. Biol. Chem. 1964, 239, 2370–2378. [Google Scholar] [PubMed]
- Kharitonov, V.G.; Sharma, V.S.; Pilz, R.B.; Magde, D.; Koesling, D. Basis of guanylate cyclase activation by carbon monoxide. Proc. Natl. Acad. Sci. USA 1995, 92, 2568–2571. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.S.; Gadelha, G.G.; Lima, S.J.; Garcia, J.A.; Medeiros, J.V.; Havt, A.; Lima, A.A.; Ribeiro, R.A.; Brito, G.A.; Cunha, F.Q.; et al. Gastroprotective effect of heme-oxygenase 1/biliverdin/CO pathway in ethanol-induced gastric damage in mice. Eur. J. Pharmacol. 2010, 642, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Costa, N.R.; Silva, R.O.; Nicolau, L.A.; Lucetti, L.T.; Santana, A.P.; Aragão, K.S.; Soares, P.M.; Ribeiro, R.A.; Souza, M.H.; Barbosa, A.L.; et al. Role of soluble guanylate cyclase activation in the gastroprotective effect of the HO-1/CO pathway against alendronate-induced gastric damage in rats. Eur. J. Pharmacol. 2013, 700, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Magierowska, K.; Magierowski, M.; Hubalewska-Mazgaj, M.; Adamski, J.; Surmiak, M.; Sliwowski, Z.; Kwiecien, S.; Brzozowski, T. Carbon monoxide (CO) released from tricarbonyldichlororuthenium (II) dimer (CORM-2) in gastroprotection against experimental ethanol-induced gastric damage. PLoS ONE 2015, 10, e0140493. [Google Scholar] [CrossRef] [PubMed]
- Klatt, P.; Schmidt, K.; Mayer, B. Brain nitric oxide synthase is a haemoprotein. Biochem. J. 1992, 288, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Srisook, K.; Han, S.S.; Choi, H.S.; Li, M.H.; Ueda, H.; Kim, C.; Cha, Y.N. CO from enhanced HO activity or from CORM-2 inhibits both O2− and NO production and downregulates HO-1 expression in LPS-stimulated macrophages. Biochem. Pharmacol. 2006, 71, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zou, X.; Chen, Y.; Zhang, P.; Shi, G. Preconditioning of carbon monoxide releasing molecule-derived CO attenuates LPS-induced activation of HUVEC. Int. J. Biol. Sci. 2008, 4, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Van der Ouderaa, F.J.; Buytenhek, M.; Slikkerveer, F.J.; van Dorp, D.A. On the hemoprotein character of prostaglandin endoperoxide synthetase. Biochim. Biophys. Acta 1979, 572, 29–42. [Google Scholar] [CrossRef]
- Percival, M.D.; Ouellet, M.; Vincent, C.J.; Yergey, J.A.; Kennedy, B.P.; O’Neill, G.P. Purification and characterization of recombinant human cyclooxygenase-2. Arch. Biochem. Biophys. 1994, 315, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Sánchez, J.; Chánez-Cárdenas, M.E. A Review on hemeoxygenase-2: Focus on cellular protection and oxygen response. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Migita, C.T.; Matera, K.M.; Ikeda-Saito, M.; Olson, J.S.; Fujii, H.; Yoshimura, T.; Zhou, H.; Yoshida, T. The oxygen and carbon monoxide reactions of heme oxygenase. J. Biol. Chem. 1998, 273, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Wang, J.; Rousseau, D.L.; Ishikawa, K.; Yoshida, T.; Takeuchi, N.; Ikeda-Saito, M. Heme-heme oxygenase complex: Structure and properties of the catalytic site from resonance Raman scattering. Biochemistry 1994, 33, 5531–5538. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, G.; Szurszewski, J.H. Carbon monoxide, hydrogen sulfide, and nitric oxide as signaling molecules in the gastrointestinal tract. Gastroenterology 2014, 147, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Jasnos, K.; Magierowski, M.; Kwiecien, S.; Brzozowski, T. Carbon monoxide in human physiology—Its role in the gastrointestinal tract. Postepy Hig. Med. Dosw. 2014, 68, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Magierowski, M.; Jasnos, K.; Kwiecien, S.; Brzozowski, T. Role of hydrogen sulfide in the physiology of gastrointestinal tract and in the mechanism of gastroprotection. Postepy Hig. Med. Dosw. 2013, 67, 150–156. [Google Scholar] [CrossRef]
- Magierowski, M.; Magierowska, K.; Kwiecien, S.; Brzozowski, T. Gaseous mediators nitric oxide and hydrogen sulfide in the mechanism of gastrointestinal integrity, protection and ulcer healing. Molecules 2015, 20, 9099–9123. [Google Scholar] [CrossRef] [PubMed]
- Tavares, A.F.; Parente, M.R.; Justino, M.C.; Oleastro, M.; Nobre, L.S.; Saraiva, L.M. The bactericidal activity of carbon monoxide-releasing molecules against Helicobacter pylori. PLoS ONE 2013, 8, e83157. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.C.; Choi, K.M.; Dutta, N.; Linden, D.R.; Szurszewski, J.H.; Gibbons, S.J.; Farrugia, G. Carbon monoxide reverses diabetic gastroparesis in NOD mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G1013–G1019. [Google Scholar] [CrossRef] [PubMed]
- Takasuka, H.; Hayashi, S.; Koyama, M.; Yasuda, M.; Aihara, E.; Amagase, K.; Takeuchi, K. Carbon monoxide involved in modulating HCO3− secretion in rat duodenum. J. Pharmacol. Exp. Ther. 2011, 337, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Okabe, S. The effects of drugs on the production and recovery processes of the stress ulcer. Jpn. J. Pharmacol. 1968, 19, 9–19. [Google Scholar] [CrossRef]
- Inoue, Y.; Fujino, Y.; Onodera, M.; Kikuchi, S.; Sato, M.; Sato, H.; Noda, H.; Kkojika, M.; Suzuki, Y.; Endo, S.A. Case of multiple hemorrhagic gastric ulcers developed via a mechanism similar to water-immersion restraint stress. Open J. Clin. Diagn. 2015, 5, 136–140. [Google Scholar] [CrossRef]
- Hirai, K.; Sasahira, T.; Ohmori, H.; Fujii, K.; Kuniyasu, H. Inhibition of heme oxygenase-1 by zinc protoporphyrin IX reduces tumor growth of LL/2 lung cancer in C57BL mice. Int. J. Cancer 2007, 120, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Ju, T.J.; Kim, Z.W.; Dan, J.M.; Kim, J.Y.; Kim, Y.D.; Seo, J.S.; Park, S.Y. Hemin, heme oxygenase-1 inducer, attenuates immobilization induced skeletal muscle atrophy in mice. Life Sci. 2013, 12, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Griffith, O.W.; Kilbourn, R.G. Nitric oxide synthase inhibitors: Amino acids. Methods Enzymol. 1996, 268, 375–392. [Google Scholar] [PubMed]
- Cechova, S.; Pajewski, T.N. The soluble guanylyl cyclase inhibitor ODQ, 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one, dose-dependently reduces the threshold for isoflurane anesthesia in rats. Anesth. Analg. 2004, 99, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Magierowski, M.; Jasnos, K.; Kwiecien, S.; Drozdowicz, D.; Surmiak, M.; Strzalka, M.; Ptak-Belowska, A.; Wallace, J.L.; Brzozowski, T. Endogenous prostaglandins and afferent sensory nerves in gastroprotective effect of hydrogen sulfide against stress-induced gastric lesions. PLoS ONE 2015, 10, e0118972. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, V.; Tandon, R.K. Stress and the gastrointestinal tract. J. Gastroenterol. Hepatol. 2005, 20, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Menguy, R.; Masters, Y.F. Gastric mucosal energy metabolism and “stress ulceration”. Ann. Surg. 1974, 180, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Brzozowski, T.; Konturek, S.J. Stress and the gut: Pathophysiology, clinical consequences, diagnostic approach and treatment options. J. Physiol. Pharmacol. 2011, 62, 591–599. [Google Scholar] [PubMed]
- Kwiecien, S.; Jasnos, K.; Magierowski, M.; Sliwowski, Z.; Pajdo, R.; Brzozowski, B.; Mach, T.; Wojcik, D.; Brzozowski, T. Lipid peroxidation, reactive oxygen species and antioxidative factors in the pathogenesis of gastric mucosal lesions and mechanism of protection against oxidative stress—Induced gastric injury. J. Physiol. Pharmacol. 2014, 65, 613–622. [Google Scholar] [PubMed]
- Ibrahim, I.; El-Sayed, S.; Abdel-Hakim, S.; Hassan, M.; Aziz, N. Inhibition of endogenous CO by ZnPP protects against stress-induced gastric lesion in adult male albino rats. J. Physiol. Biochem. 2012, 68, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Morsy, M.A.; Heeba, G.H.; Abdelwahab, S.A.; Rofaeil, R.R. Protective effects of nebivolol against cold restraint stress-induced gastric ulcer in rats: Role of NO, HO-1, and COX-1,2. Nitric Oxide 2012, 27, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yang, M.; Ma, N.; Shinohara, H.; Semba, R. Contribution of carbon monoxide-producing cells in the gastric mucosa of rat and monkey. Histochem. Cell Biol. 1998, 109, 369–373. [Google Scholar] [PubMed]
- Takeuchi, K.; Aihara, E.; Kimura, M.; Dogishi, K.; Hara, T.; Hayashi, S. Gas mediators involved in modulating duodenal HCO3− secretion. Curr. Med. Chem. 2012, 19, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Ueyama, T.; Yoshida, K.; Kimura, H.; Ito, T.; Shimizu, Y.; Oka, M.; Tsuruo, Y.; Ichinose, M. Adaptive HNENrf2-HO-1 pathway against oxidative stress is associated with acute gastric mucosal lesions. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G460–G469. [Google Scholar] [CrossRef] [PubMed]
- Sawle, P.; Foresti, R.; Mann, B.E.; Johnson, T.R.; Green, C.J.; Motterlini, R. Carbon monoxide-releasing molecules (CO-RMs) attenuate the inflammatory response elicited by lipopolysaccharide in RAW264.7 murine macrophages. Br. J. Pharmacol. 2005, 145, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Oates, P.J.; Hakkinen, J.P. Studies on the mechanism of ethanol-induced gastric damage in rats. Gastroenterology 1988, 94, 10–21. [Google Scholar] [PubMed]
- Mayer, E.A. The neurobiology of stress and gastrointestinal disease. Gut 2000, 47, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Peskar, B.M. Role of cyclooxygenase isoforms in gastric mucosal defence. J. Physiol. Paris 2001, 95, 3–9. [Google Scholar] [CrossRef]
- Brzozowski, T.; Konturek, P.C.; Konturek, S.J.; Drozdowicz, D.; Pajdo, R.; Pawlik, M.; Brzozowska, I.; Hahn, E.G. Expression of cyclooxygenase (COX)-1 and COX-2 in adaptive cytoprotection induced by mild stress. J. Physiol. Paris 2000, 94, 83–91. [Google Scholar] [CrossRef]
- Takeuchi, K. Gastric cytoprotection by prostaglandin E₂ and prostacyclin: Relationship to EP1 and IP receptors. J. Physiol. Pharmacol. 2014, 65, 3–14. [Google Scholar] [PubMed]
- Wang, T.; Leng, Y.F.; Zhang, Y.; Xue, X.; Kang, Y.Q.; Zhang, Y. Oxidative stress and hypoxia-induced factor 1α expression in gastric ischemia. World J. Gastroenterol. 2011, 17, 1915–1922. [Google Scholar] [CrossRef] [PubMed]
- Nakao, A.; Moore, B.A.; Murase, N.; Liu, F.; Zuckerbraun, B.S.; Bach, F.H.; Choi, A.M.; Nalesnik, M.A.; Otterbein, L.E.; Bauer, A.J. Immunomodulatory effects of inhaled carbon monoxide on rat syngeneic small bowel graft motility. Gut 2003, 52, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.A.; Otterbein, L.E.; Türler, A.; Choi, A.M.; Bauer, A.J. Inhaled carbon monoxide suppresses the development of postoperative ileus in the murine small intestine. Gastroenterology 2003, 124, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Kaizu, T.; Nakao, A.; Tsung, A.; Toyokawa, H.; Sahai, R.; Geller, D.A.; Murase, N. Carbon monoxide inhalation ameliorates cold ischemia/reperfusion injury after rat liver transplantation. Surgery 2005, 138, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Kaizu, T.; Ikeda, A.; Nakao, A.; Tsung, A.; Toyokawa, H.; Ueki, S.; Geller, D.A.; Murase, N. Protection of transplant-induced hepatic ischemia/reperfusion injury with carbon monoxide via MEK/ERK1/2 pathway downregulation. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G236–G244. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Brzozowski, T.; Duda, A.; Kwiecien, S.; Löber, S.; Dembinski, A.; Hahn, E.G.; Konturek, S.J. Epidermal growth factor and prostaglandin E2 accelerate mucosal recovery from stress-induced gastric lesions via inhibition of apoptosis. J. Physiol. Paris 2001, 95, 361–367. [Google Scholar] [CrossRef]
- Konturek, S.J.; Brzozowski, T.; Pytko-Polonczyk, J.; Drozdowicz, D. Exogenous and endogenous cholecystokinin protects gastric mucosa against the damage caused by ethanol in rats. Eur. J. Pharmacol. 1995, 273, 57–62. [Google Scholar] [CrossRef]
- Brzozowski, T.; Konturek, P.C.; Sliwowski, Z.; Pajdo, R.; Drozdowicz, D.; Kwiecien, S.; Burnat, G.; Konturek, S.J.; Pawlik, W.W. Prostaglandin/cyclooxygenase pathway in ghrelin-induced gastroprotection against ischemia-reperfusion injury. J. Pharmacol. Exp. Ther. 2006, 319, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Magierowski, M.; Jasnos, K.; Sliwowski, Z.; Surmiak, M.; Krzysiek-Maczka, G.; Ptak-Belowska, A.; Kwiecien, S.; Brzozowski, T. Exogenous asymmetric dimethylarginine (ADMA) in pathogenesis of ischemia-reperfusion-induced gastric lesions: Interaction with protective nitric oxide (NO) and calcitonin gene-related peptide (CGRP). Int. J. Mol. Sci. 2014, 15, 4946–4964. [Google Scholar] [CrossRef] [PubMed]
- Adamski, J.; Zuba, D. Gas chromatographic determination of carboxyhaemoglobin (COHb) in post-mortem blood samples. Probl. Forensic Sci. 2012, 89, 86–98. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Chan, H.; Ning, J.; Lu, K.; Ma, D. The role of hydrogen sulfide in pathologies of the vital organs and its clinical application. J. Physiol. Pharmacol. 2015, 66, 169–179. [Google Scholar] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| β-actin | 5′-GATCAAGATCATTGCTCCTCCTG-3′ | 5′-AGGGTGTAAAACGCAGCTCA-3′ |
| HO-1 | 5′-GTCCCAGGATTTGTCCGAGG-3′ | 5′-GGAGGCCATCACCAGCTTAAA-3′ |
| HO-2 | 5′-CCGGGCAGAAAATACCCAGT-3′ | 5′-ATCAGTGCTTCCTTCCGGTG-3′ |
| HIF-1α | 5′-ATCCATTTTCAGCTCAGGACAC-3′ | 5′-GGTAGGTTTCTGTAACTGGGTCTG-3′ |
| iNOS | 5′-TGGTGAGGGGACTGGACTTT-3′ | 5′-CTCCGTGGGGCTTGTAGTTG-3′ |
| COX-2 | 5′-ATCAGAACCGCATTGCCTCT-3′ | 5′-GCCAGCAATCTGTCTGGTGA-3′ |
| TNF-α | 5′-TGGGCTCCCTCTCATCAGTT-3′ | 5′-TCCGCTTGGTGGTTTGCTAC-3′ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magierowska, K.; Magierowski, M.; Surmiak, M.; Adamski, J.; Mazur-Bialy, A.I.; Pajdo, R.; Sliwowski, Z.; Kwiecien, S.; Brzozowski, T. The Protective Role of Carbon Monoxide (CO) Produced by Heme Oxygenases and Derived from the CO-Releasing Molecule CORM-2 in the Pathogenesis of Stress-Induced Gastric Lesions: Evidence for Non-Involvement of Nitric Oxide (NO). Int. J. Mol. Sci. 2016, 17, 442. https://doi.org/10.3390/ijms17040442
Magierowska K, Magierowski M, Surmiak M, Adamski J, Mazur-Bialy AI, Pajdo R, Sliwowski Z, Kwiecien S, Brzozowski T. The Protective Role of Carbon Monoxide (CO) Produced by Heme Oxygenases and Derived from the CO-Releasing Molecule CORM-2 in the Pathogenesis of Stress-Induced Gastric Lesions: Evidence for Non-Involvement of Nitric Oxide (NO). International Journal of Molecular Sciences. 2016; 17(4):442. https://doi.org/10.3390/ijms17040442
Chicago/Turabian StyleMagierowska, Katarzyna, Marcin Magierowski, Marcin Surmiak, Juliusz Adamski, Agnieszka Irena Mazur-Bialy, Robert Pajdo, Zbigniew Sliwowski, Slawomir Kwiecien, and Tomasz Brzozowski. 2016. "The Protective Role of Carbon Monoxide (CO) Produced by Heme Oxygenases and Derived from the CO-Releasing Molecule CORM-2 in the Pathogenesis of Stress-Induced Gastric Lesions: Evidence for Non-Involvement of Nitric Oxide (NO)" International Journal of Molecular Sciences 17, no. 4: 442. https://doi.org/10.3390/ijms17040442
APA StyleMagierowska, K., Magierowski, M., Surmiak, M., Adamski, J., Mazur-Bialy, A. I., Pajdo, R., Sliwowski, Z., Kwiecien, S., & Brzozowski, T. (2016). The Protective Role of Carbon Monoxide (CO) Produced by Heme Oxygenases and Derived from the CO-Releasing Molecule CORM-2 in the Pathogenesis of Stress-Induced Gastric Lesions: Evidence for Non-Involvement of Nitric Oxide (NO). International Journal of Molecular Sciences, 17(4), 442. https://doi.org/10.3390/ijms17040442