
Metabolic Fingerprinting to Assess the Impact of Salinity on Carotenoid Content in Developing Tomato Fruits


Abstract
:
1. Introduction
2. Results
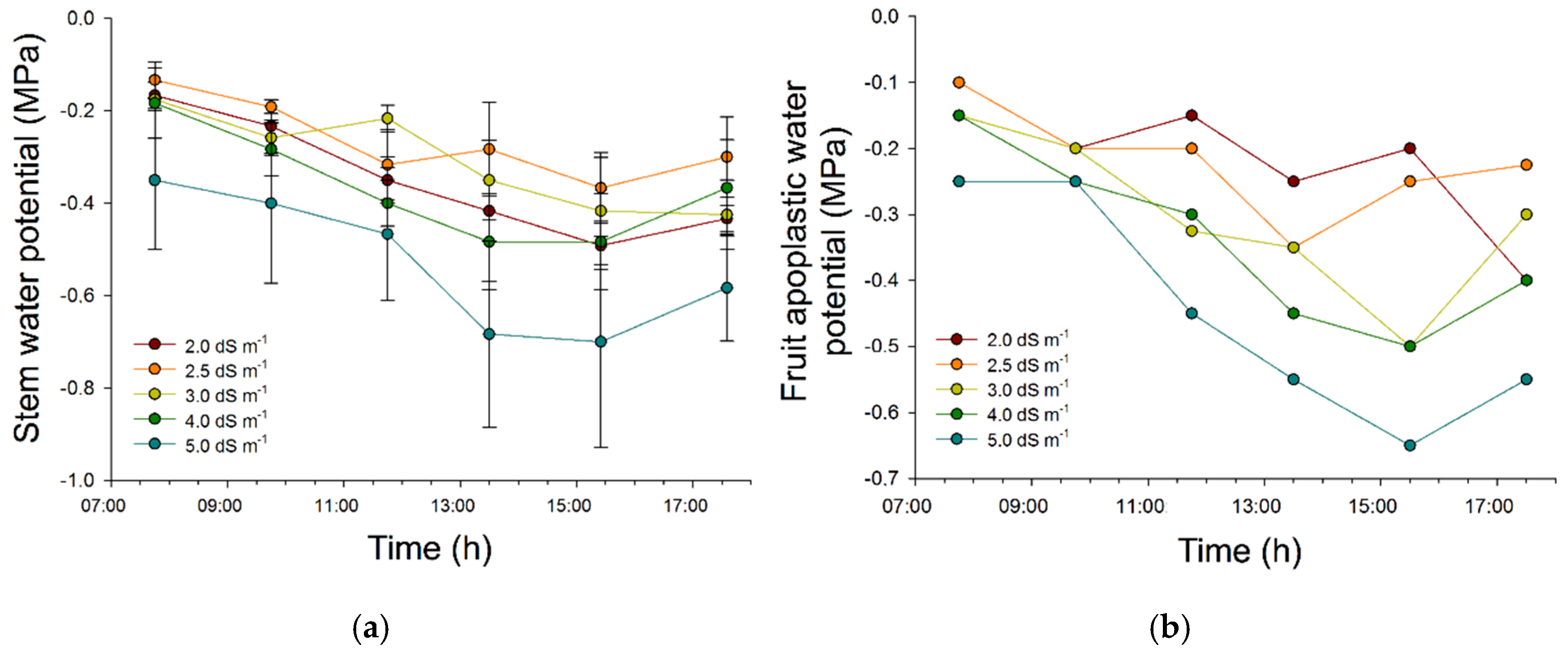
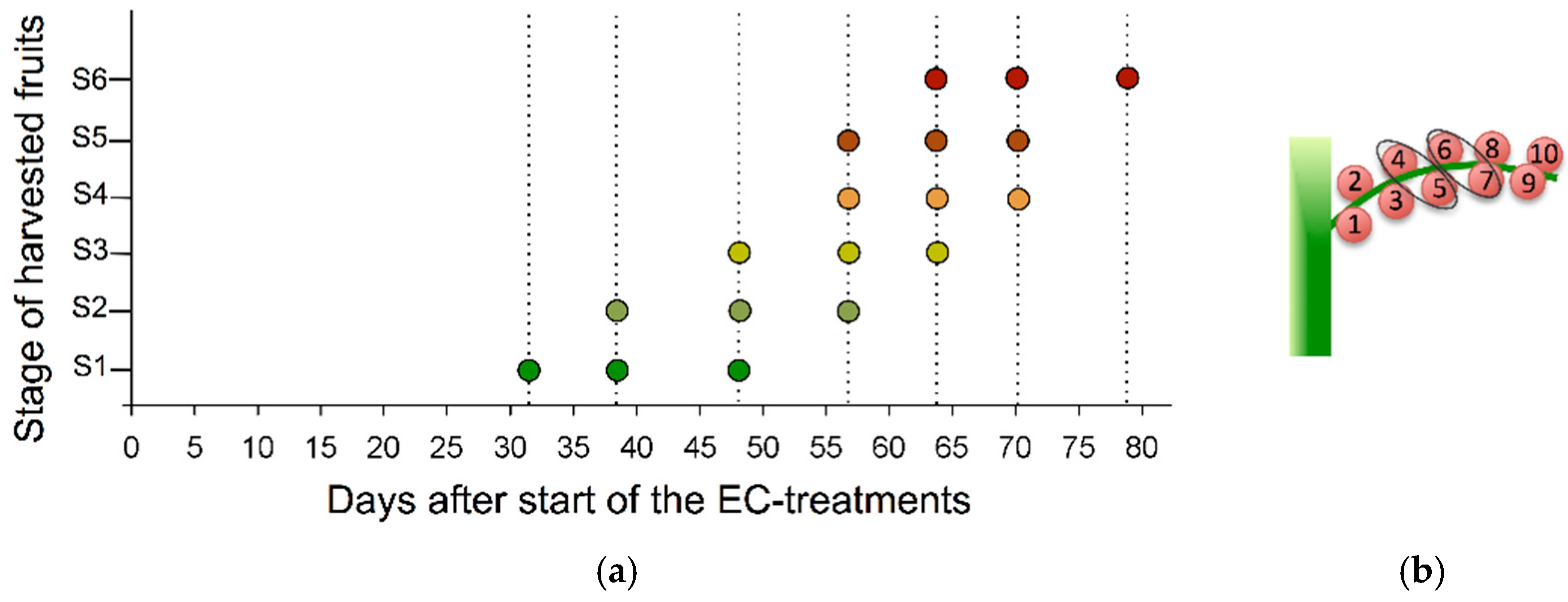
2.1. Effectiveness of Imposed Salinity Treatments
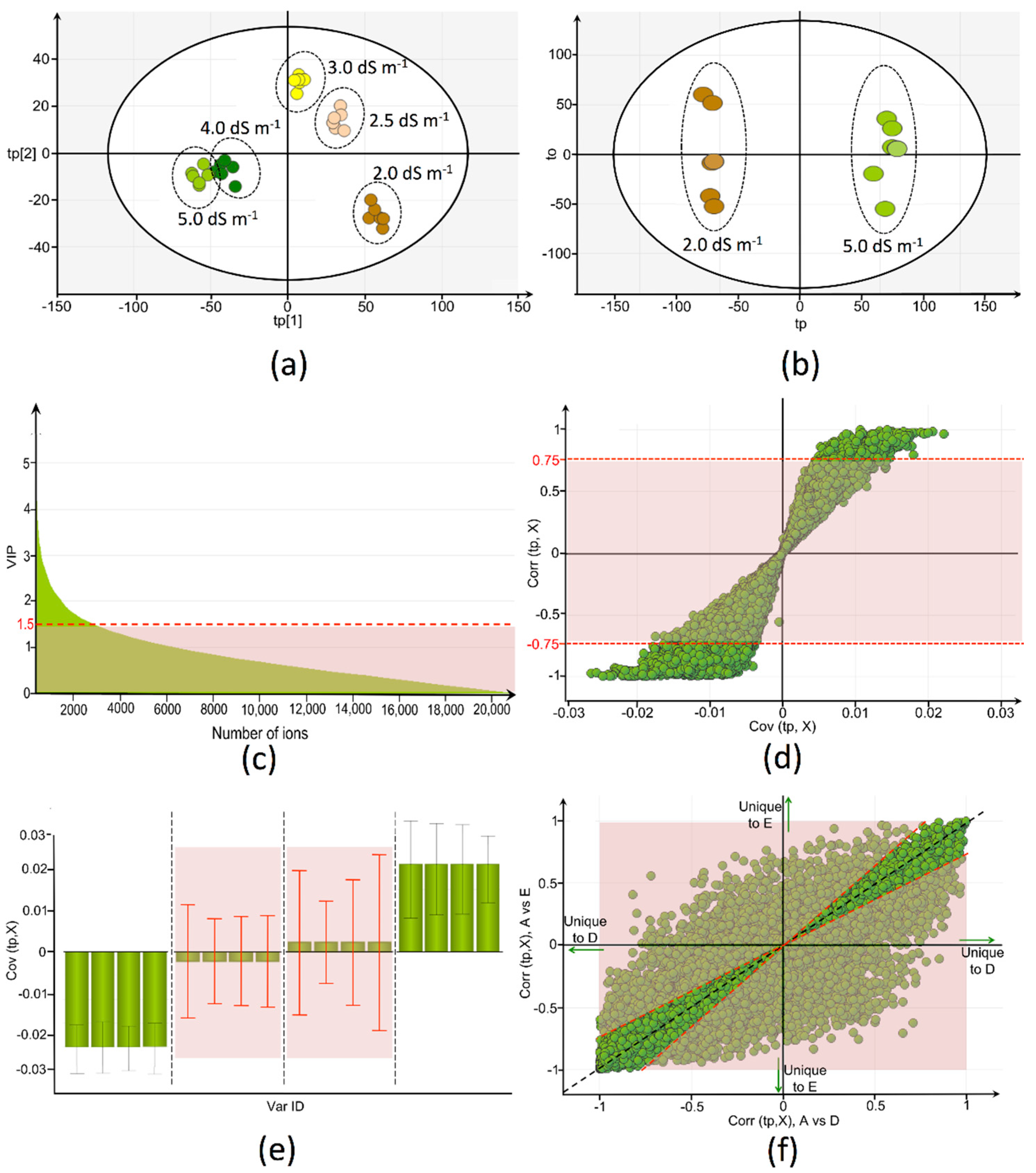
2.2. Metabolic Fingerprinting of Tomato Fruits
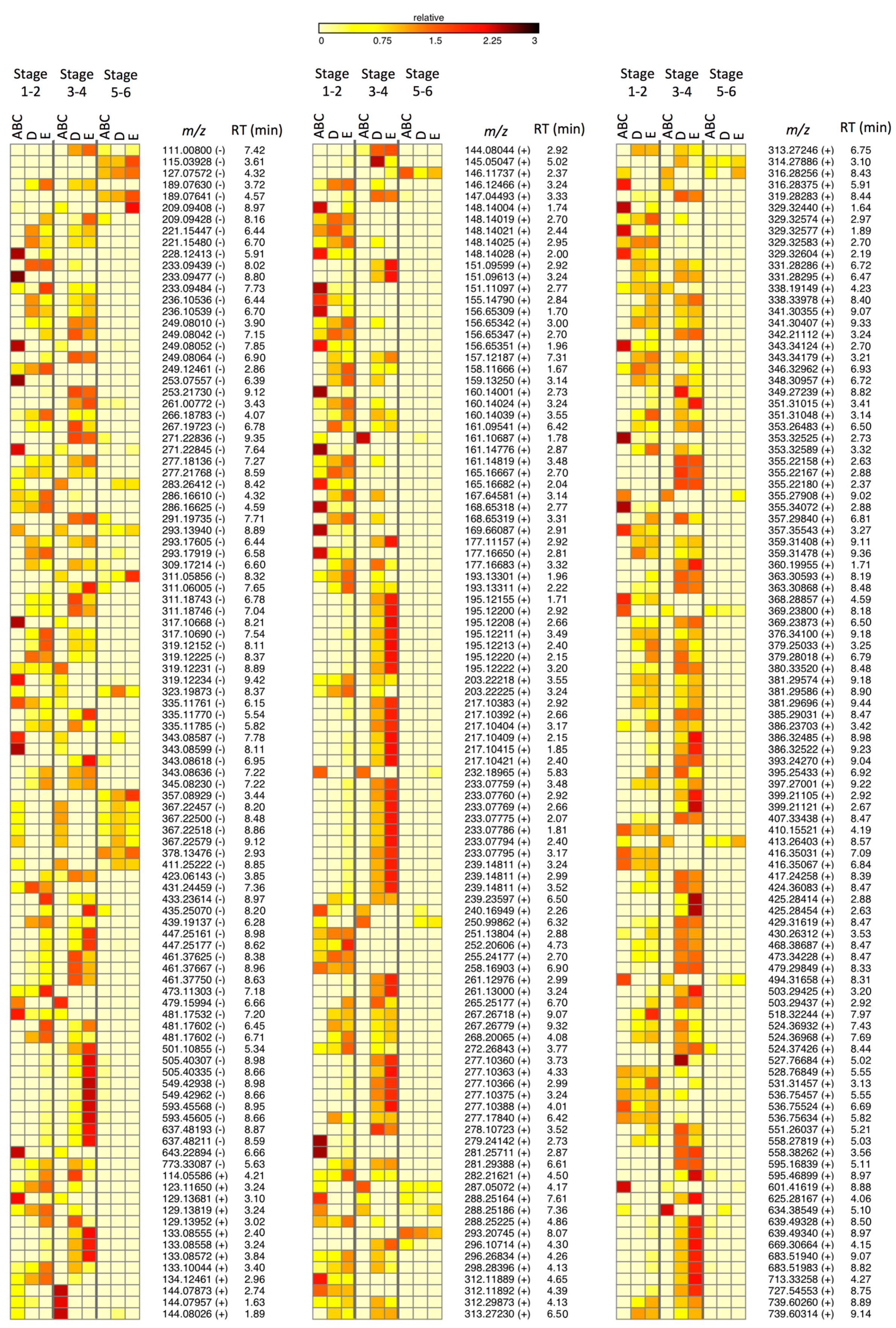
2.3. Metabolites Involved in the Salinity Stress Response
2.4. Metabolites Involved in Carotenoid Metabolism
2.5. Potential of Altered Salinity to Improve Tomato Nutritional Quality
3. Discussion
3.1. Effectiveness of Imposed Salinity Treatments
3.2. Impact of Salinity on the Tomato Fruit Metabolome
3.3. The Potential of Salinity to Improve Carotenoid Content
4. Materials and Methods
4.1. Plant Material and Growing Conditions
4.2. Experimental Set-Up
4.3. Chemicals and Reagents
4.4. Targeted Analysis of Four Tomato Fruit Carotenoids
4.5. Untargeted Analysis of Phytohormones and Related Metabolites
4.6. Establishment of Tomato Fruit Metabolic Fingerprints
4.7. Screening for Metabolites Involved in the Salinity Stress Response
4.8. Screening for Metabolites Involved in Carotenoid Metabolism
4.9. Effectiveness of Imposed Salinity Treatments
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| EC | Electrical conductivity |
| MS | Mass spectrometry |
| OPLS | Orthogonal partial least squares to latent structures |
| DA | Discriminant analysis |
| DAA | Days after anthesis |
| VIP | Variable importance in projection |
| CIJFjk | Jack-knifed confidence interval |
| SUS | Shared and unique structure |
| dd-MS2 | data-dependent fragmentation |
| CID | Compound identifier |
References
- Raiola, A.; Rigano, M.M.; Calafiore, R.; Frusciante, L.; Barone, A. Enhancing the health-promoting effects of tomato fruit for biofortified food. Mediat. Inflamm. 2014. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.L.; Lee, A.H.; Binns, C.W. Lycopene and prostate cancer: Emerging evidence. Expert Rev. Anticancr Ther. 2005, 5, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Bioactivity and protective effects of natural carotenoids. Bba-Mol. Basis Dis. 2005, 1740, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Fanasca, S.; Martino, A.; Heuvelink, E.; Stanghellini, C. Effect of electrical conductivity, fruit pruning, and truss position on quality in greenhouse tomato fruit. J. Hortic. Sci. Biotechnol. 2007, 82, 488–494. [Google Scholar] [CrossRef]
- Wu, M.; Kubota, C. Effects of high electrical conductivity of nutrient solution and its application timing on lycopene, chlorophyll and sugar concentrations of hydroponic tomatoes during ripening. Sci. Hortic. 2008, 116, 122–129. [Google Scholar] [CrossRef]
- Taber, H.; Perkins-Veazie, P.; Li, S.; White, W.; Roderniel, S.; Xu, Y. Enhancement of tomato fruit lycopene by potassium is cultivar dependent. Hortscience 2008, 43, 159–165. [Google Scholar]
- Garcia-Closas, R.; Berenguer, A.; Tormo, M.J.; Sanchez, M.J.; Quiros, J.R.; Navarro, C.; Arnaud, R.; Dorronsoro, M.; Chirlaque, M.D.; Barricarte, A.; et al. Dietary sources of vitamin C, vitamin E and specific carotenoids in Spain. Br. J. Nutr. 2004, 91, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- De Pascale, S.; Maggio, A. Effect of salt stress on water relations and antioxidant activity in tomato. Acta Hortic. 2003, 39–46. [Google Scholar] [CrossRef]
- Veit-Kohler, U.; Krumbein, A.; Kosegarten, H. Effect of different water supply on plant growth and fruit quality of Lycopersicon esculentum. J. Plant Nutr. Soil Sc. 1999, 162, 583–588. [Google Scholar] [CrossRef]
- Gomez, J.M.; Hernandez, J.A.; Jimenez, A.; del Rio, L.A.; Sevilla, F. Differential response of antioxidative enzymes of chloroplasts and mitochondria to long-term NaCl stress of pea plants. Free Radic. Res. 1999, 31, 11–18. [Google Scholar] [CrossRef]
- Smirnoff, N. Antioxidant Systems and Plant Response to the Environment. In Environment and Plant Metabolism; Smirnoff, N., Ed.; Bios Scientific Publishers: Oxford, UK, 1995; pp. 217–243. [Google Scholar]
- Borghesi, E.; Gonzalez-Miret, M.L.; Escudero-Gilete, M.L.; Malorgio, F.; Heredia, F.J.; Melendez-Martinez, A.J. Effects of salinity stress on carotenoids, anthocyanins, and color of diverse tomato genotypes. J. Agric. Food Chem. 2011, 59, 11676–11682. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Papadantonakis, N.; Lydakis-Simantiris, N.; Kefalas, P.; Savvas, D. Effects of silicon and salinity on fruit yield and quality of tomato grown hydroponically. Int. Sym. Manag. Greenh. Crop. Saline Environ. 2003, 609, 141–147. [Google Scholar] [CrossRef]
- Krauss, S.; Schnitzler, W.H.; Grassmann, J.; Woitke, M. The influence of different electrical conductivity values in a simplified recirculating soilless system on inner and outer fruit quality characteristics of tomato. J. Agric. Food Chem. 2006, 54, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Krumbein, A.; Schwarz, D.; Klaring, H.P. Effects of environmental factors on carotenoid content in tomato (Lycopersicon esculentam (L.) Mill.) grown in a greenhouse. J. Appl. Bot. Food Qual. 2006, 80, 160–164. [Google Scholar]
- Van Meulebroek, L.; Vanhaecke, L.; de Swaef, T.; Steppe, K.; de Brabandert, H. U-HPLC-MS/MS to quantify liposoluble antioxidants in red-ripe tomatoes, grown under different salt stress levels. J. Agric. Food Chem. 2012, 60, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Ehret, D.L.; Usher, K.; Helmer, T.; Block, G.; Steinke, D.; Frey, B.; Kuang, T.; Diarra, M. Tomato fruit antioxidants in relation to salinity and greenhouse climate. J. Agric. Food Chem. 2013, 61, 1138–1145. [Google Scholar] [CrossRef] [PubMed]
- Dumas, Y.; Dadomo, M.; di Lucca, G.; Grolier, P. Effects of environmental factors and agricultural techniques on antioxidant content of tomatoes. J. Sci. Food Agric. 2003, 83, 369–382. [Google Scholar] [CrossRef]
- Dorais, M.D.; Ehret, D.L.; Papadopoulos, A. Tomato (Solanum lycopersicum) health components: From the seed to the consumer. Phytochem. Rev. 2008, 7, 231–250. [Google Scholar] [CrossRef]
- Maggio, A.; Barbieri, G.; Raimondi, G.; de Pascale, S. Contrasting effects of GA3 treatments on tomato plants exposed to increasing salinity. J. Plant Growth Regul. 2010, 29, 63–72. [Google Scholar] [CrossRef]
- Srivastava, A.; Handa, A.K. Hormonal regulation of tomato fruit development: A molecular perspective. J. Plant Growth Regul. 2005, 24, 67–82. [Google Scholar] [CrossRef]
- Cho, H.W.; Kim, S.B.; Jeong, M.K.; Park, Y.; Miller, N.G.; Ziegler, T.R.; Jones, D.P. Discovery of metabolite features for the modelling and analysis of high-resolution NMR spectra. Int. J. Data Min. Bioinform. 2008, 2, 176–192. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.K.; Findlay, J. Bioanalytical method validation—A revisit with a decade of progress. Parm. Rev. 2002, 17, 1551–1557. [Google Scholar]
- Wager, S.; Hastie, T.; Efron, B. Confidence intervals for random forests: The jackknife and the infinitesimal jackknife. J. Mach. Learn. Res. 2014, 15, 1625–1651. [Google Scholar] [PubMed]
- Wiklund, S.; Johansson, E.; Sjostrom, L.; Mellerowicz, E.J.; Edlund, U.; Shockcor, J.P.; Gottfries, J.; Moritz, T.; Trygg, J. Visualization of GC/TOF-MS-based metabolomics data for identification of biochemically interesting compounds using OPLS class models. Anal. Chem. 2008, 80, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Uehara, T.; Yanagida, M. Highly accurate chemical formula prediction tool utilizing high-resolution mass spectra, MS/MS fragmentation, heuristic rules, and isotope pattern matching. Anal. Chem. 2012, 84, 4396–4403. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Schmidt, S.; Muller-Hannemann, M.; Neumann, S. In silico fragmentation for computer assisted identification of metabolite mass spectra. BMC Bioinform. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Van Meulebroek, L.; Vanden Bussche, J.; de Clercq, N.; Steppe, K.; Vanhaecke, L. A metabolomics approach to unravel the regulating role of phytohormones towards carotenoid metabolism in tomato fruit. Metabolomics 2015, 11, 667–683. [Google Scholar] [CrossRef]
- Surekha, C.; Aruna, L.V.; Hossain, M.A.; Wani, S.H.; Rao, N.; Neelapu, R. Prospects status of future prospects of transgenic approaches for salt tolerance in plants/crop plants. In Managing Salt Tolerance in Plants; Wani, S.H., Hossain, M.A., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 329–352. [Google Scholar]
- Barciszewski, J.; Siboska, G.; Rattan, S.I.S.; Clark, B.F.C. Occurrence, biosynthesis and properties of kinetin (N6-furfuryladenine). Plant Growth Regul. 2000, 32, 257–265. [Google Scholar] [CrossRef]
- Chakrabarti, N.; Mukherji, S. Alleviation of NaCl stress by pretreatment with phytohormones in Vigna radiata. Biol. Plantarum. 2003, 46, 589–594. [Google Scholar] [CrossRef]
- Liu, L.; Shao, Z.; Zhang, M.; Wang, Q. Regulation of carotenoid metabolism in tomato. Mol. Plant 2015, 8, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.; Osborne, M.; Ebden, M.; Reece, S.; Gibson, N.; Aigrain, S. Gaussian processes for time-series modelling. Philos. Trans. R. Soc. A 2013, 371, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plantarum. 2008, 132, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Carrari, F.; Baxter, C.; Usadel, B.; Urbanczyk-Wochniak, E.; Zanor, M.I.; Nunes-Nesi, A.; Nikiforova, V.; Centero, D.; Ratzka, A.; Pauly, M.; et al. Integrated analysis of metabolite and transcript levels reveals the metabolic shifts that underlie tomato fruit development and highlight regulatory aspects of metabolic network behavior. Plant Physiol. 2006, 142, 1380–1396. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.; Creissen, G.; Kular, B.; Firmin, J.; Robinson, S.; Verhoeyen, M.; Mullineaux, P. Changes in oxidative processes and components of the antioxidant system during tomato fruit ripening. Planta 2002, 214, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Shalata, A.; Tal, M. The effect of salt stress on lipid peroxidation and antioxidants in the leaf of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii. Physiol. Plantarum. 1998, 104, 169–174. [Google Scholar] [CrossRef]
- Ghanem, M.E.; Albacete, A.; Martinez-Andujar, C.; Acosta, M.; Romero-Aranda, R.; Dodd, I.C.; Lutts, S.; Perez-Alfocea, F. Hormonal changes during salinity-induced leaf senescence in tomato (Solanum lycopersicum L.). J. Exp. Bot. 2008, 59, 3039–3050. [Google Scholar] [CrossRef] [PubMed]
- Tanksley, S.D. The genetic, developmental, and molecular bases of fruit size and shape variation in tomato. Plant Cell 2004, 16, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Chretien, S.; Gosselin, A.; Dorais, M. High electrical conductivity and radiation-based water management improve fruit quality of greenhouse tomatoes grown in rockwool. Hortscience 2000, 35, 627–631. [Google Scholar]
- Yurtseven, E.; Kesmez, G.D.; Unlukara, A. The effects of water salinity and potassium levels on yield, fruit quality and water consumption of a native central anatolian tomato species (Lycopersicon esculantum). Agric. Water Manag. 2005, 78, 128–135. [Google Scholar] [CrossRef]
- Rose, J.K.C.; Bennett, A.B. Cooperative disassembly of the cellulose-xyloglucan network of plant cell walls: Parallels between cell expansion and fruit ripening. Trends Plant Sci. 1999, 4, 176–183. [Google Scholar] [CrossRef]
- Sanchez, D.H.; Siahpoosh, M.R.; Roessner, U.; Udvardi, M.; Kopka, J. Plant metabolomics reveals conserved and divergent metabolic responses to salinity. Physiol. Plantarum. 2008, 132, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.K.; Willumsen, J.; Kaack, K. Composition and taste of tomatoes as affected by increased salinity and different salinity sources. J. Hortic. Sci. Biotechnol. 1998, 73, 205–215. [Google Scholar] [CrossRef]
- Shi, J.; le Maguer, M.; Bryan, M. Lycopene from tomatoes. In Functional Foods: Biochemical and Processing Aspects; Shi, J.M., Mazza, G., le Maguer, M., Eds.; CRC Press LLC: Florida, USA, 2003; pp. 135–167. [Google Scholar]
- Dorais, M.; Papadopoulos, A.P.; Gosselin, A. Influence of electric conductivity management on greenhouse tomato yield and fruit quality. Agronomie 2001, 21, 367–383. [Google Scholar] [CrossRef]
- Fanasca, S.; Colla, G.; Maiani, G.; Venneria, E.; Rouphael, Y.; Azzini, E.; Saccardo, F. Changes in antioxidant content of tomato fruits in response to cultivar and nutrient solution composition. J. Agric. Food Chem. 2006, 54, 4319–4325. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Xu, X.N.; Zhu, H.S.; Liu, A.H.; Liu, L.; Li, J.M.; Hua, X.J. Comparative transcriptomic profiling of a salt-tolerant wild tomato species and a salt-sensitive tomato cultivar. Plant Cell Physiol. 2010, 51, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.P.; Antunez, A.; Araya, H.; Pertuze, R.; Fuentes, L.; Lizana, X.C.; Lutts, S. Salt stress differently affects growth, water status and antioxidant enzyme activities in Solanum lycopersicum and its wild relative Solanum chilense. Aust. J. Bot. 2014, 62, 359–368. [Google Scholar] [CrossRef]
- Caro, M.; Cruz, V.; Cuartero, J.; Estan, M.T.; Bolarin, M.C. Salinity tolerance of normal-fruited and cherry tomato cultivars. Plant Soil. 1991, 136, 249–255. [Google Scholar] [CrossRef]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2013. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotox. Environ. Safe 2004, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Cuartero, J.; Fernandez-Munoz, R. Tomato and salinity. Sci. Hortic-Amsterdam. 1999, 78, 83–125. [Google Scholar] [CrossRef]
- Maggio, A.; Raimondi, G.; Martino, A.; de Pascale, S. Salt stress response in tomato beyond the salinity tolerance threshold. Environ. Exp. Bot. 2007, 59, 276–282. [Google Scholar] [CrossRef]
- Van Meulebroek, L.; Vanden Bussche, J.; Steppe, K.; Vanhaecke, L. High-resolution Orbitrap mass spectrometry for the analysis of carotenoids in tomato fruit: Validation and comparative evaluation towards UV-VIS and tandem mass spectrometry. Anal. Bioanal. Chem. 2014, 406, 2613–2626. [Google Scholar] [CrossRef] [PubMed]
- Van Meulebroek, L.; Vanden Bussche, J.; Steppe, K.; Vanhaecke, L. Ultra-high performance liquid chromatography coupled to high resolution Orbitrap mass spectrometry for metabolomic profiling of the endogenous phytohormonal status of the tomato plant. J. Chromatogr. A 2012, 1260, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Trygg, J.; Holmes, E.; Lundstedt, T. Chemometrics in metabonomics. J. Proteome Res. 2007, 6, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, L.; Trygg, J.; Wold, S. CV-ANOVA for significance testing of PLS and OPLS® models. J. Chemometr. 2008, 22, 594–600. [Google Scholar] [CrossRef]
- Suzuki, M.; Kusano, M.; Takahashi, H.; Nakamura, Y.; Hayashi, N.; Kobayashi, M.; Ichikawa, T.; Matsui, M.; Hirochika, H.; Saito, K. Rice-Arabidopsis FOX line screening with FT-NIR-based fingerprinting for GC-TOF/MS-based metabolite profiling. Metabolomics 2010, 6, 137–145. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.S.; Yao, M.; Zhao, X.J.; Fritsche, J.; Schmitt-Kopplin, P.; Cai, Z.W.; Wan, D.F.; Lu, X.; Yang, S.L.; et al. Metabonomics study on the effects of the ginsenoside Rg3 in a β-cyclodextrin-based formulation on tumor-bearing rats by a fully automatic hydrophilic interaction/reversed-phase column-switching HPLC-ESI-MS approach. Anal. Chem. 2008, 80, 4680–4688. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Description | R2(X) | R2(Y) | Q2(Y) | |
|---|---|---|---|---|
| S1 | A versus C | 0.852 | 1.000 | 0.824 |
| A versus D | 0.419 | 0.990 | 0.900 | |
| A versus E | 0.694 | 0.996 | 0.952 | |
| S2 | A versus D | 0.785 | 1.000 | 0.940 |
| A versus E | 0.658 | 0.998 | 0.953 | |
| S3 | A versus E | 0.700 | 0.999 | 0.890 |
| S4 | A versus D | 0.721 | 0.999 | 0.965 |
| A versus E | 0.696 | 1.000 | 0.891 | |
| S5 | A versus D | 0.551 | 0.996 | 0.865 |
| A versus E | 0.562 | 0.999 | 0.861 | |
| S6 | A versus C | 0.626 | 0.992 | 0.884 |
| A versus D | 0.528 | 0.999 | 0.935 | |
| A versus E | 0.585 | 0.938 | 0.938 | |
| Model Description | R2(X) | R2(Y) | Q2(Y) |
|---|---|---|---|
| Lutein | 0.564 | 0.957 | 0.915 |
| Zeaxanthin | 0.660 | 0.906 | 0.698 |
| β-Carotene | 0.592 | 0.647 | 0.308 |
| Lycopene | 0.672 | 0.988 | 0.929 |
| Metabolite ID | Ionization Mode | m/z-Value | Retention Time (min) | CID | Fragmentation Ratio (%) | Elemental Composition |
|---|---|---|---|---|---|---|
| 1 | + | 476.2991 | 7.28 | 11698581 | 100.0 | C27H41N1O6 |
| 2 | + | 269.2096 | 7.13 | 21720337 | 85.0 | C16H28O3 |
| 3 | + | 409.1486 | 5.39 | 44144278 | 81.3 | C20H24O9 |
| 4 | + | 711.2440 | 4.63 | 10349855 | 100.0 | C33H42O17 |
| 5 | + | 576.2689 | 3.89 | 46242589 | 64.2 | C25H42N3O10P |
| 6 a | + | 785.8416 | 4.73 | n.a. | ||
| 7 a | + | 572.4492 | 8.36 | n.a. | ||
| 8 | + | 311.2166 | 6.35 | 20686694 | 57.1 | C13H30N2O6 |
| 9 | + | 337.2015 | 2.99 | 19511475 | 86.7 | C20H24N4O |
| 10 a | + | 602.4972 | 6.26 | n.a. | ||
| 11 | + | 355.1776 | 4.61 | 15690336 | 100.0 | C15H30O7S |
| 12 | + | 275.0906 | 4.70 | 28848874 | 41.7 | C13H12N3O4 |
| 13 | + | 596.4120 | 4.63 | 44409845 | 70.0 | C33H57NO8 |
| 10416091 | ||||||
| 14 | + | 531.2604 | 3.49 | 3188562 | 50.0 | C30H34N4O5 |
| 3188563 | ||||||
| 15 | + | 528.3285 | 5.94 | 44275272 | 50.0 | C24H50NO9P |
| 16 | + | 530.2776 | 6.70 | 24999541 | 50.0 | C30H35N5O4 |
| 17 | + | 485.1479 | 5.26 | 314405 | 100.0 | C22H28O10S |
| 18 | + | 221.1138 | 5.98 | 18519108 | 100.0 | C8H16N2O5 |
| 19 | + | 348.2155 | 6.20 | 22997503 | 76.4 | C20H29NO4 |
| 20 | + | 390.1299 | 4.58 | 46657456 | 78.6 | C19H15N7O3 |
| 21 | + | 462.1048 | 2.85 | 1190411 | 100.0 | C14H19N7O9S |
| 22 | + | 410.1552 | 4.19 | 27682727 | 71.4 | C19H27N3O3S2 |
| 32647280 | ||||||
| 23 | + | 377.1055 | 3.60 | 45420283 | 70.5 | C19H20O6S |
| 24 | + | 775.3393 | 5.67 | 21630904 | 66.7 | C36H54O18 |
| 25 | + | 188.0636 | 4.14 | 89915 | 73.7 | C11H9NO2 |
| 26 | − | 163.0394 | 3.26 | 691 | 100.0 | C9H8O3 |
| 27 a | − | 565.0488 | 5.52 | n.a. | ||
| 28 | − | 773.3309 | 5.64 | 11767898 | 76.9 | C43H46N6O8 |
| 29 | − | 736.3310 | 3.86 | 21681260 | 50.0 | C40H51NO2 |
| 30 a | − | 115.0393 | 3.61 | n.a. | ||
| 31 | − | 431.2446 | 7.37 | 12133739 | 85.8 | C25H36O6 |
| 32 | − | 378.1348 | 2.94 | 25232866 | 83.2 | C22H21NO5 |
| 33 | − | 425.1469 | 5.38 | 36942227 | 75.0 | C21H22N4O6 |
| 34 | − | 569.0984 | 6.55 | 23406912 | 77.8 | C26H26N4O5S3 |
| 35 | − | 603.2794 | 8.08 | 44282378 | 100 | C32H44O11 |
| 36 | − | 189.0764 | 4.58 | 26339616 | 85.6 | C8H14O5 |
| 37 | − | 249.1246 | 2.86 | 45809090 | 100.0 | C13H18N2O3 |
| 18108416 | ||||||
| 43403806 | ||||||
| 43421263 | ||||||
| 43392898 | ||||||
| 38 | − | 286.1661 | 4.33 | 21403706 | 85.7 | C14H25NO5 |
| 39 | − | 378.1351 | 2.65 | 45898135 | 93.3 | C22H21NO5 |
| 40 | − | 461.0163 | 4.36 | 15614249 | 87.5 | C12H21NO4 |
| 41 | − | 242.1400 | 5.52 | 30309738 | 36.4 | C13H17N5 |
| 42 | − | 361.0696 | 3.29 | 1014968 | 100.0 | C17H18N2O3S2 |
| 43 | − | 463.2711 | 7.37 | 10457172 | 66.7 | C26H40O7 |
| 9825709 | ||||||
| 9847284 | ||||||
| 44 | − | 486.0865 | 5.67 | 30663446 | 71.4 | C24H17N5O5S |
| 45 | − | 777.2091 | 5.12 | 20488220 | 100.0 | C35H36N7O12S |
| 46 | − | 189.0764 | 4.83 | 23399204 | 71.4 | C8H14O5 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Meulebroek, L.; Hanssens, J.; Steppe, K.; Vanhaecke, L. Metabolic Fingerprinting to Assess the Impact of Salinity on Carotenoid Content in Developing Tomato Fruits. Int. J. Mol. Sci. 2016, 17, 821. https://doi.org/10.3390/ijms17060821
Van Meulebroek L, Hanssens J, Steppe K, Vanhaecke L. Metabolic Fingerprinting to Assess the Impact of Salinity on Carotenoid Content in Developing Tomato Fruits. International Journal of Molecular Sciences. 2016; 17(6):821. https://doi.org/10.3390/ijms17060821
Chicago/Turabian StyleVan Meulebroek, Lieven, Jochen Hanssens, Kathy Steppe, and Lynn Vanhaecke. 2016. "Metabolic Fingerprinting to Assess the Impact of Salinity on Carotenoid Content in Developing Tomato Fruits" International Journal of Molecular Sciences 17, no. 6: 821. https://doi.org/10.3390/ijms17060821
APA StyleVan Meulebroek, L., Hanssens, J., Steppe, K., & Vanhaecke, L. (2016). Metabolic Fingerprinting to Assess the Impact of Salinity on Carotenoid Content in Developing Tomato Fruits. International Journal of Molecular Sciences, 17(6), 821. https://doi.org/10.3390/ijms17060821