
An Amphiprotic Novel Chitosanase from Bacillus mycoides and Its Application in the Production of Chitooligomers with Their Antioxidant and Anti-Inflammatory Evaluation


 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Screening and Identification of a Chitosanase-Producing Strain
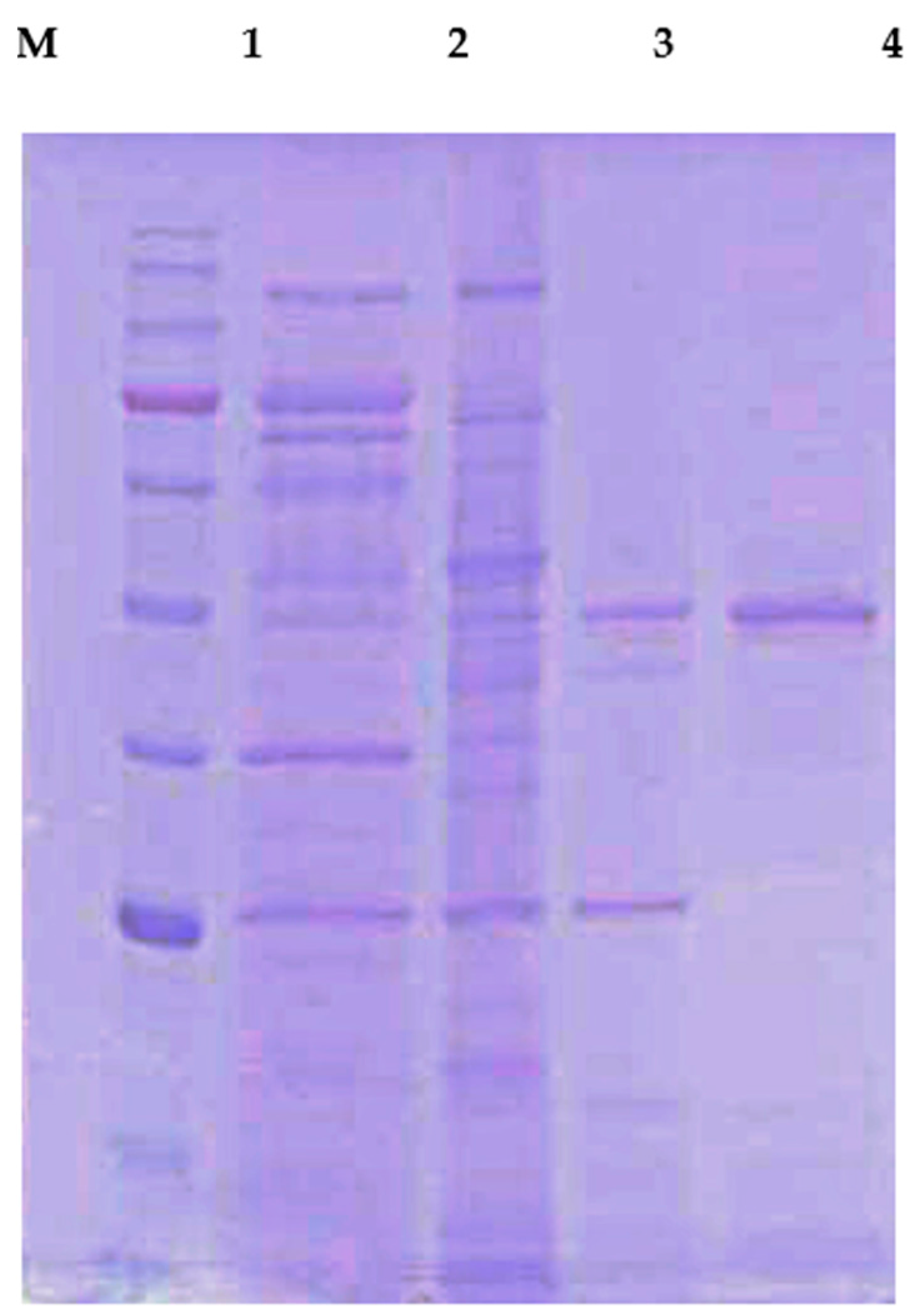
2.2. Production and Purification of Chitosanase
2.3. Identification of CS038 by LC-MS/MS Analysis
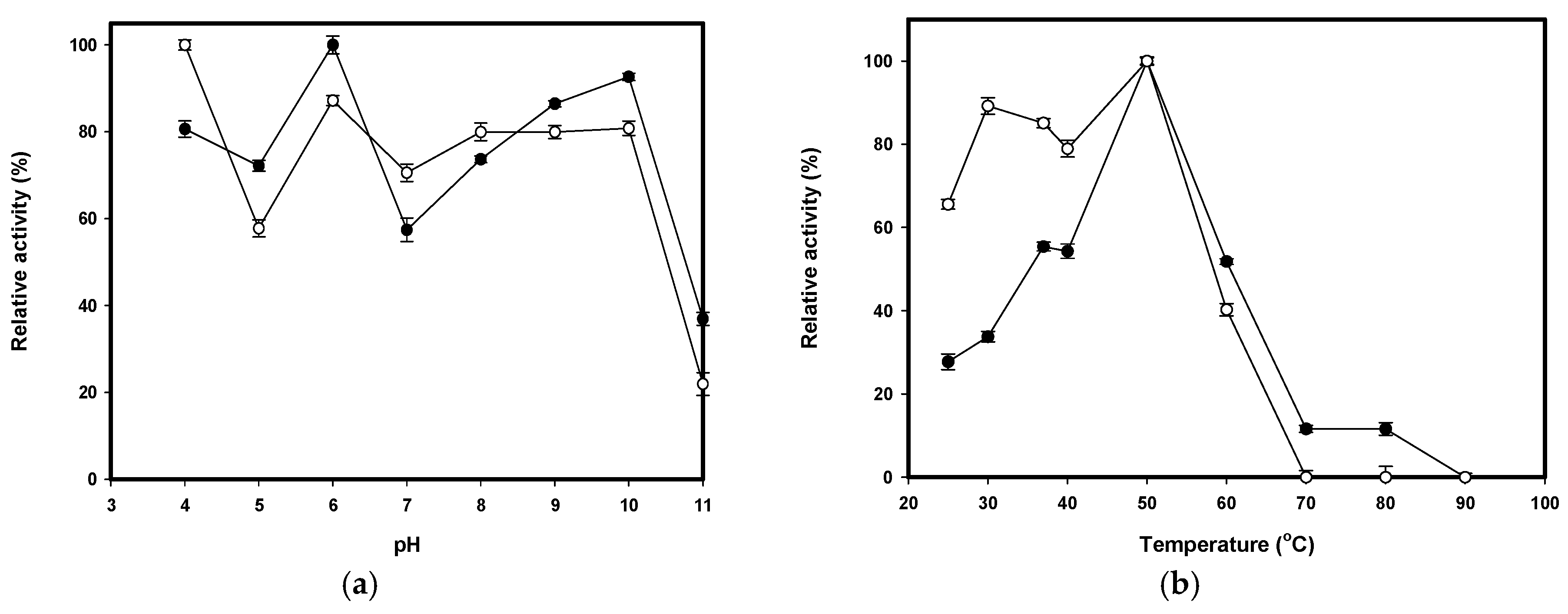
2.4. Effect of pH and Temperature on the Activity and Stability of CS038
2.5. Substrate Specificity and Kinetic Parameters
2.6. Effects of Metal Ions
2.7. Chitosan Hydrolysis
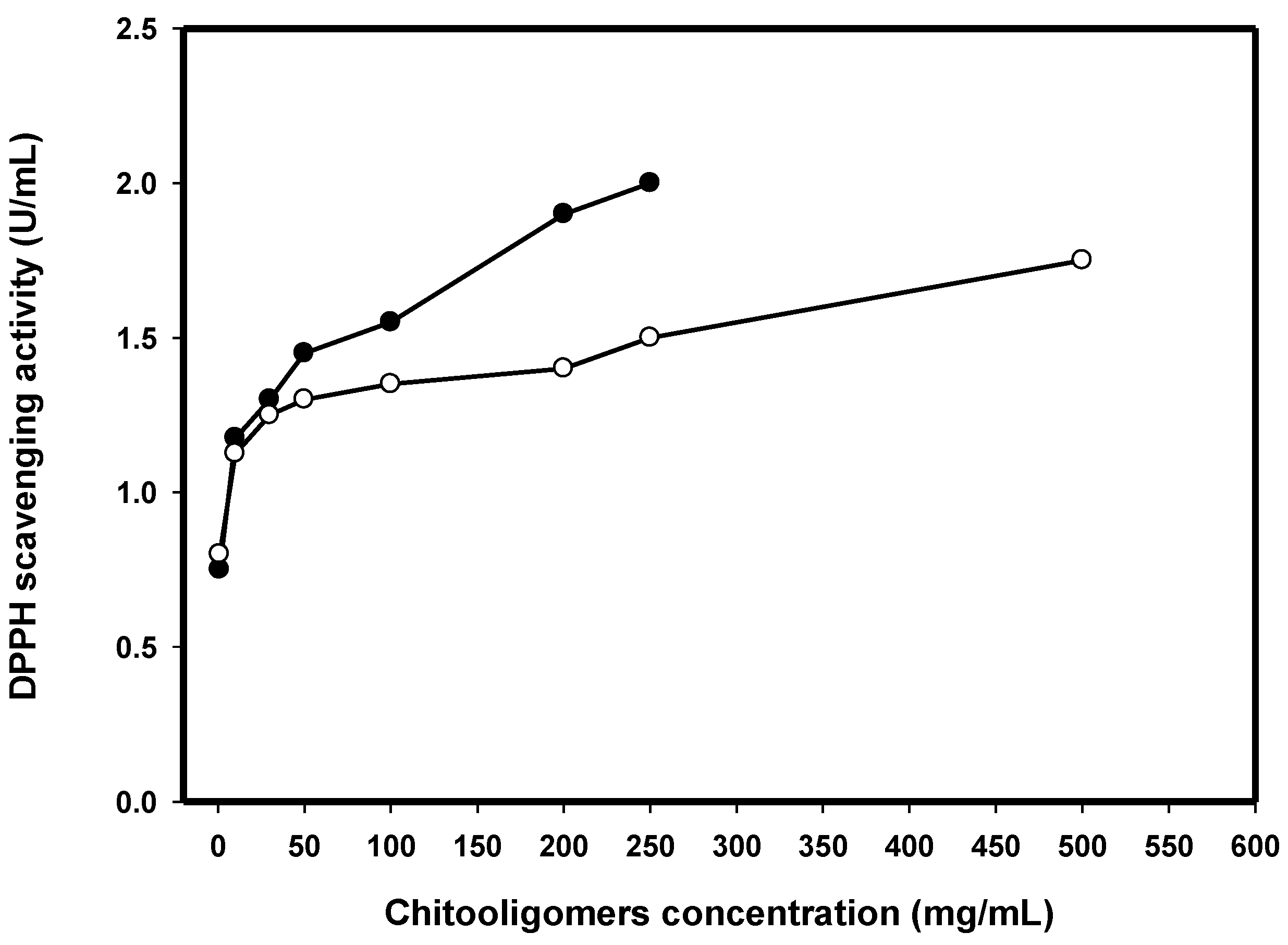
2.8. DPPH Radical Scavenging Activity of COS
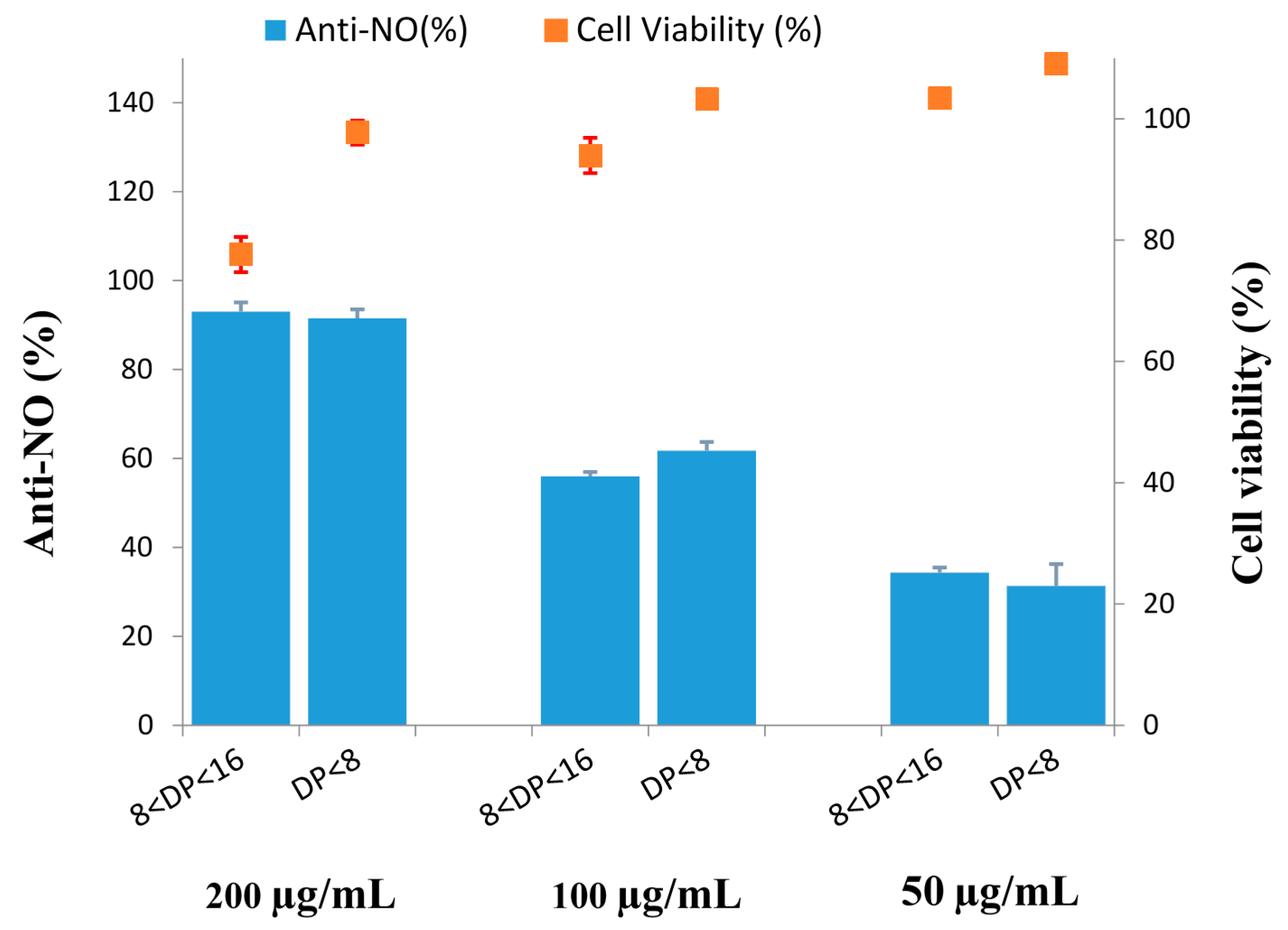
2.9. Effect of COS on Cytotoxicity and Anti-Inflammation
2.10. Antitumoral Activities of COS
3. Experimental
3.1. Materials
3.2. Screening of Chitosanase-Producing Strains
3.3. Chitosanase Activity Assay
3.4. Purification of Chitosanase
3.5. Mass Spectrometry and Protein Identification
3.6. Effects of pH and Temperature on Enzyme Activity and Stability
3.7. Kinetic Parameters
3.8. Effects of Various Metal Ions on Chitosanase Activities
3.9. Enzymatic Production of the Chitosan Oligosaccharides
3.10. Measurement of DPPH Radical Scavenging Activity
3.11. Assay for Anti-Proliferation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, S.L.; Liang, T.W.; Yen, Y.H. Bioconversion of chitin-containing wastes for the production of enzymes and bioactive materials. Carbohydr. Polym. 2011, 84, 732–742. [Google Scholar] [CrossRef]
- Lodhi, G.; Kim, Y.S.; Hwang, J.W.; Kim, S.K.; Jeon, Y.J.; Je, J.Y.; Ahn, C.B.; Moon, S.H.; Jeon, B.T.; Park, P.J. Chitooligosaccharide and its derivatives: Preparation and biological applications. BioMed Res. Int. 2014, 2014, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Liu, K.C.; Liang, T.W.; Kuo, Y.H.; Wang, C.Y. In vitro antioxidant activity of liquor and semi-purified fractions from squid pen biowaste by Serratia ureilytica TKU013. Food Chem. 2010, 119, 1380–1385. [Google Scholar] [CrossRef]
- Busilacchi, A.; Gigante, A.; Mattioli-Belmonte, M.; Manzotti, S.; Muzzarelli, R.A.A. Chitosan stabilizes platelet growth factors and modulates stem cell differentiation toward tissue regeneration. Carbohydr. Polym. 2013, 98, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Chen, Y.J.; Yen, Y.H.; Wang, S.L. The antitumor activity of the hydrolysates of chitinous materials hydrolyzed by crude enzyme from Bacillus amyloliquefaciens V656. Process Biochem. 2007, 42, 527–534. [Google Scholar] [CrossRef]
- Wang, S.L.; Lin, T.Y.; Yen, Y.H.; Liao, H.F.; Chen, Y.J. Bioconversion of shellfish chitin wastes for the production of Bacillus subtilis W-118 chitinase. Carbohydr. Res. 2006, 341, 2507–2515. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Lin, H.T.; Liang, T.W.; Chen, Y.J.; Yen, Y.H.; Guo, S.P. Reclamation of chitinous materials by bromelain for the preparation of antitumor and antifungal materials. Bioresour. Technol. 2008, 99, 4386–4393. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.A.; Ju, W.T.; Jung, W.J.; Park, R.D. Purification and characterization of chitosanase from Bacillus cereus D-11. Carbohydr. Polym. 2008, 72, 513–520. [Google Scholar] [CrossRef]
- Wang, S.L.; Chen, T.R.; Liang, T.W.; Wu, P.C. Conversion and degradation of shellfish wastes by Bacillus cereus TKU018 fermentation for the production of chitosanases and bioactive materials. Biochem. Eng. J. 2009, 48, 111–117. [Google Scholar] [CrossRef]
- Kobayashi, T.; Koide, O.; Deguchi, S.; Horikoshi, K. Characterization of chitosanase of a deep biosphere Bacillus strain. Biosci. Biotechnol. Biochem. 2011, 75, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Pechsrichuang, P.; Yoohat, K.; Yamabhai, M. Production of recombinant Bacillus subtilis chitosanase, suitable for biosynthesis of chitosan-oligosaccharides. Bioresour. Technol. 2013, 127, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Peng, J.H.; Liang, T.W.; Liu, K.C. Purification and characterization of a chitosanase from Serratia marcescens TKU011. Carbohydr. Res. 2008, 343, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, M.G.; Hansen, O.C.; Stougaard, P. Isolation, characterization and heterologous expression of a novel chitosanase from Janthinobacterium sp. strain 4239. Microb. Cell Fact. 2010, 9. [Google Scholar] [CrossRef] [PubMed]
- Zitouni, M.; Fortin, M.; Scheerle, R.K.; Letzel, T.; Matteau, D.; Rodrigue, S.; Brzezinski, R. Biochemical and molecular characterization of a thermostable chitosanase produced by the strain Paenibacillus sp. 1794 newly isolated from compost. Appl. Microbiol. Biotechnol. 2013, 97, 5801–5813. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Chen, S.J.; Wang, C.L. Purification and characterization of chitinases and chitosanases from a new species strain Pseudomonas sp. TKU015 using shrimp shells as a substrate. Carbohydr. Res. 2008, 343, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Tseng, W.N.; Liang, T.W. Biodegradation of shellfish wastes and production of chitosanases by a squid pen-assimilating bacterium, Acinetobacter calcoaceticus TKU024. Biodegradation 2011, 22, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Chen, D.; Chen, L.; Yang, G.; Zou, S. Purification, characterization, and action mode of a chitosanase from Streptomyces roseolus induced by chitin. Carbohydr. Res. 2012, 355, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Hsieh, J.L.; Wang, S.L. Production and purification of a protease, a chitosanase, and chitinoligosaccharides by Bacillus cereus TKU022 fermentation. Carbohydr. Res. 2012, 362, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Su, J.W.; Liang, T.W.; Nguyen, A.D.; Wang, S.L. Production, purification and characterization of a chitosanase from Bacillus cereus. Res. Chem. Intermed. 2014, 40, 2237–2248. [Google Scholar] [CrossRef]
- Liang, T.W.; Huang, C.T.; Nguyen, A.D.; Wang, S.L. Squid pen chitin chitooligomers as food colorants absorbers. Mar. Drugs 2015, 13, 681–696. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Lo, B.C.; Wang, S.L. Chitinolytic bacteria-assisted conversion of squid pen and its effect on dyes and pigments adsorption. Mar. Drugs 2015, 13, 4576–4593. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Kim, E.J.; Piao, Z.; Yun, Y.C.; Shin, Y.C. Purification and characterization of chitosanase from Bacillus sp. strain KCTC 0377BP and its application for the production of chitosan oligosaccharides. Appl. Environ. Microbiol. 2004, 70, 4522–4531. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Kao, T.Y.; Wang, C.L.; Yen, Y.H.; Chern, M.K.; Chen, Y.H. A solvent stable metalloprotease produced by Bacillus sp. TKU004 and its application in the deproteinization of squid pen for β-chitin preparation. Enzym. Microb. Technol. 2006, 39, 724–731. [Google Scholar] [CrossRef]
- Wang, S.L.; Yeh, P.Y. Purification and characterization of a chitosanase from a nattokinase producing strain Bacillus subtilis TKU007. Process Biochem. 2008, 43, 132–138. [Google Scholar] [CrossRef]
- Chiang, C.L.; Chang, C.T.; Sung, H.Y. Purification and properties of chitosanase from a mutant of Bacillus subtilis IMR-NK1. Enzyme. Microb. Technol. 2003, 32, 260–267. [Google Scholar] [CrossRef]
- Lee, Y.S.; Park, I.H.; Yoo, J.S.; Chung, S.Y.; Lee, Y.C.; Cho, Y.S.; Ahn, S.C.; Kim, C.M.; Choi, Y.L. Cloning, purification, and characterization of chitinase from Bacillus sp. DAU101. Bioresour. Technol. 2007, 98, 2734–2741. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.I.; Kang, T.H.; Chung, K.J.; Kim, I.S.; Chung, K.C. Purification of a constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and expression of the gene. FEMS Microbiol. Lett. 2004, 240, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Yabuki, M.; Uchiyama, A.; Suzuki, A.; Ando, A.; Fuji, T. Purification and properties of chitosanase from Bacillus circulans MH-Kl. J. Gen. Appl. Microbiol. 1988, 34, 255–270. [Google Scholar] [CrossRef]
- Ohtakara, A.; Ogata, H.; Taketomi, Y.; Mitsutomi, M. Purification and Characterization of Chitosanase from Streptomyces Griseus; Academic Press: Orlando, FL, USA, 1984; pp. 147–160. [Google Scholar]
- Wang, S.L.; Chang, T.J.; Liang, T.W. Conversion and degradation of shellfish wastes by Serratia sp. TKU016 fermentation for the production of enzymes and bioactive materials. Biodegradation 2010, 21, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Chen, Y.Y.; Pan, P.S.; Wang, S.L. Purification of chitinase/chitosanase from Bacillus cereus and discovery of an enzyme inhibitor. Int. J. Biol. Macromol. 2014, 63, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Hedges, A.; Wolfe, R.S. Extracellular enzyme from Mycobacter A1-1 that exhibits both β-1-4 glucanase and chitosanase activities. J. Bacteriol. 1974, 120, 844–853. [Google Scholar] [PubMed]
- Pradeep, G.C.; Choi, Y.H.; Choi, Y.S.; Suh, S.E.; Seong, J.H.; Cho, S.S.; Bae, M.S.; Yoo, J.C. An extremely alkaline novel chitinase from Streptomyces sp. CS495. Process Biochem. 2014, 49, 223–229. [Google Scholar] [CrossRef]
- Je, J.Y.; Park, P.J.; Kim, S.K. Radical scavenging activity of hetero-chitooligosaccharides. Eur. Food Res. Technol. 2004, 219, 60–65. [Google Scholar] [CrossRef]
- Liu, H.T.; Li, W.M.; Xu, G.; Li, X.Y.; Bai, X.F.; Wei, P.; Yu, C.; Du, Y.G. Chitosan oligosaccharides attenuate hydrogen peroxide-induced stress injury in human umbilicalvein endothelial cells. Pharmacol. Res. 2009, 59, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.N.; Kim, M.M.; Kim, S.K. Chitin oligosaccharides inhibit oxidativestress in live cells. Carbohydr. Polym. 2008, 74, 228–234. [Google Scholar] [CrossRef]
- Yuan, W.P.; Liu, B.; Liu, C.H.; Wang, X.J.; Zhang, M.S.; Meng, X.M.; Xia, X.K. Antioxidant activity of chito-oligosaccharides on pancreatic isletcells in streptozotocin-induced diabetes in rats. World J. Gastroenterol. 2009, 15, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.M.; Yu, S.H.; Zhang, L.S.; Zhao, Z.Y.; Dong, L.L. Effects of several acetylated chitooligosaccharides on antioxidation, antiglycation and NO generation in erythrocyte. Bioorg. Med. Chem. Lett. 2014, 24, 4053–4057. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Lin, C.L.; Liang, T.W.; Liu, K.C.; Kuo, Y.H. Conversion of squid pen by Serratia ureilytica for the production of enzymes and antioxidants. Bioresour. Technol. 2009, 100, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Pinero Estrada, J.E.; Bermejo Bescos, P.; Villar del Fresno, A.M. Antioxidant activity of different fractions of Spriulina platensis protean extract. Farmaco 2001, 56, 497–500. [Google Scholar] [CrossRef]
- Lin, H.Y.; Chou, C.C. Antioxidant activities of water-soluble disaccharide chitosan derivatives. Food Res. Int. 2004, 37, 883–889. [Google Scholar] [CrossRef]
- Xing, R.; Yu, H.; Liu, S.; Zhang, W.; Zhang, Q.; Li, Z. Antioxidative activity of differently regioselective chitosan sulfates in vitro. Bioorg. Med. Chem. 2005, 13, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Xing, R.; Liu, S.; Li, P. Advances in preparation, analysis and biological activities of single chitooligosaccharides. Carbohydr. Polym. 2016, 139, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.J.; Park, J.K.; Park, Y.I. Anti-inflammatory effects of low-molecular weight chitosan oligosaccharides in IgE-antigen complex-stimulated RBL-2H3 cells and asthma model mice. Int. Immunopharmcol. 2012, 12, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Hossaine, M.D.A.; Park, S.C. A potential anti-inflammation activity and depigmentation effect of Lespedeza bicolor extract and its fractions. Saudi J. Biol. Sci. 2016, 23, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Yuan, J.; Wu, S.; Huang, J.; Xu, X.; Wu, Z.; Gao, H. Anti-inflammation furanoditerpenoids from Caesalpiniaminax Hance. Phytochemistry 2015, 117, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Kuda, T.; Kawahara, M.; Nemoto, M.; Takahashi, H.; Kimura, B. In vitro antioxidant and anti-inflammation properties of lactic acid bacteria isolated from fish intestines and fermented fish from the Sanriku Satoumi region in Japan. Food Res. Int. 2014, 64, 248–255. [Google Scholar] [CrossRef]
- Zhai, X.T.; Zhang, Z.Y.; Jiang, C.H.; Chen, J.Q.; Ye, J.Q.; Jia, X.B.; Yang, Y.; Ni, Q.; Wang, S.X.; Song, J.; et al. Nauclea officinalis inhibits inflammation in LPS-mediated RAW 264.7 macrophages by suppressing the NF-κB signaling pathway. J. Ethnopharmacol. 2016, 183, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Wu, W.; Liu, Z.; Xie, C.; Tang, M.; Li, S.; Yang, J.; Tang, H.; Chen, K.; Long, C.; et al. Bioactivity-guided isolation of anti-inflammation flavonoids from the stems of Millettia dielsiana Harms. Fitoterapia 2014, 95, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Chiou, C.T.; Cheng, J.J.; Huang, H.C.; Kuo-Yang, L.M.; Liao, C.C.; Bastow, K.F.; Lee, K.H.; Kuo, Y.H. Cytotoxic polyisoprenyl benzophenonoids from Garcinia subelliptica. J. Nat. Prod. 2010, 73, 557–562. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Total | Specific Activity (U/mg) | Purification (Fold) | Recovery (%) | ||
|---|---|---|---|---|---|---|
| Volume (mL) | Protein (mg) | Activity (U) | ||||
| Culture supernatant | 768 | 2764.8 | 443.9 | 0.16 | 1 | 100 |
| (NH4)2SO4 precipitation | 45 | 751.5 | 386.0 | 0.51 | 3.19 | 87.0 |
| DEAE-sepharose | 40 | 128.2 | 261.4 | 2.04 | 12.75 | 58.9 |
| Macro-Prep DEAE | 10 | 7.5 | 156.1 | 20.81 | 130.06 | 35.2 |
| Strains | MW (kDa) | Optimal | Stability | Inhibitor | References | ||
|---|---|---|---|---|---|---|---|
| Temp. (°C) | pH | Temp. (°C) | pH | ||||
| B. mycoides TKU038 | 48 | 50 | 6, 10 | 25–50 | 4–10 | Cu2+, Ba2+, Zn2+, Fe2+, Mn2+, EDTA, PMSF | This study |
| B. cereus D-11 | 41 | 60 | 6 | <50 | 5–10 | Cu2+, Hg2+, Pb2+ | [8] |
| B. cereus TKU022 | 44 | 60 | 7 | 25–40 | 7–10 | Mn2+ | [18] |
| B. cereus TKU031 | 43 | 50 | 5 | 20–50 | 5–9 | Fe2+, Cu2+, Zn2+, Mn2+, EDTA | [19] |
| B. cereus TKU033 | 43 | 50 | 5 | <40 | 5–7 | Cu2+, Mn2+, EDTA | [20] |
| B. cereus TKU034 | 43 | 50 | 7 | <50 | 4.5–7.5 | Fe2+, Ca2+, Cu2+, Zn2+, Mn2+, EDTA | [21] |
| Bacillus sp. KCTC 0377BP | 45 | 60 | 4–6 | <55 | 4–8 | Mn2+, Hg2+ | [22] |
| Bacillus sp. TKU004 | 29 | 37 | 7 | <40 | 4–7 | Cu2+, Fe2+ | [23] |
| B. subtilis TKU007 | 25 | 37 | 7 | <37 | 4–9 | Cu2+, Fe2+, EDTA | [24] |
| B. subtilis IMR-NK1 | 36 | 45 | 4 | <40 | 5–9 | Hg2+, PHMB | [25] |
| Bacillus sp. DAU101 | 27 | 50 | 7.5 | - | - | Cu2+, Zn2+, Hg2+, Ni2+, Co2+ | [26] |
| Bacillus sp. MET 1299 | 52 | 60 | 5.5 | - | - | Mn2+, Cu2+, Zn2+, Co2+, EDTA | [27] |
| B. criculans MH-K1 | 32 | 50 | 6.5 | - | - | Hg2+, Cd2+, Ni2+, Zn2+, pCMB | [28] |
| Streptomyces griseus | 35 | 37 | 8 | - | - | Ag2+, Hg2+, Fe2+, Cu2+, pCMB | [29] |
| Streptomyces roseolus | 41 | 50 | 5 | 30–60 | 5–7 | Mn2+, Cu2+, Zn2+, Co2+, EDTA | [17] |
| Serratia sp. TKU016 | 65 | 50 | 7 | <50 | 6–7 | Mn2+ | [30] |
| S. marcescens TKU011 | 21 | 50 | 5 | <50 | 4–8 | Mn2+, Cu2+, PMSF | [12] |
| Acinetobacter calcoaceticus TKU024 | 66 | 60 | 7 | <70 | 6–11 | Mn2+, EDTA | [16] |
| 27 | 50 | 6 | <90 | 4–10 | |||
| Peptide Sequence | Identified Protein and Coverage Rate | Accession Number |
|---|---|---|
| 81SYYDNWKK88 | Chitosanase 54% | Bacillus cereus: gi446936339 |
| 93NDLSSLPGGYYVKGEITGDADGFK | ||
| PLGTSEGQGYGMIITVLMAGYDSNAQKIYDGLFK150 | ||
| 157SSQNPNLMGWVVADSKKAQGHFDSATDGD | ||
| LDIAYSLLLAHKQWGSNGTVNYLKEAKDMITK217 | ||
| 221ASNVTNNNRLNLGDWDSKSSLD | ||
| TRPSDWMMSHLRAFYEFTGDK263 | ||
| 283YSPNTGLISDFVVKNPPQPAPKDFLEE | ||
| SEYTNAYYYNASR322 | ||
| 327IVMDYAMYGEK337 | ||
| 346VSSWIQNK353 | ||
| 397WVNSGWDWMK406 |
| Chemicals | Relative Activity (%) |
|---|---|
| None | 100 |
| Na+ | 94 |
| Mg2+ | 93 |
| Fe2+ | 0 |
| Ca2+ | 88 |
| Cu2+ | 21 |
| Ba2+ | 57 |
| Zn2+ | 20 |
| Mn2+ | 0 |
| EDTA | 0 |
| PMSF | 0 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, T.-W.; Chen, W.-T.; Lin, Z.-H.; Kuo, Y.-H.; Nguyen, A.D.; Pan, P.-S.; Wang, S.-L. An Amphiprotic Novel Chitosanase from Bacillus mycoides and Its Application in the Production of Chitooligomers with Their Antioxidant and Anti-Inflammatory Evaluation. Int. J. Mol. Sci. 2016, 17, 1302. https://doi.org/10.3390/ijms17081302
Liang T-W, Chen W-T, Lin Z-H, Kuo Y-H, Nguyen AD, Pan P-S, Wang S-L. An Amphiprotic Novel Chitosanase from Bacillus mycoides and Its Application in the Production of Chitooligomers with Their Antioxidant and Anti-Inflammatory Evaluation. International Journal of Molecular Sciences. 2016; 17(8):1302. https://doi.org/10.3390/ijms17081302
Chicago/Turabian StyleLiang, Tzu-Wen, Wei-Ting Chen, Zhi-Hu Lin, Yao-Haur Kuo, Anh Dzung Nguyen, Po-Shen Pan, and San-Lang Wang. 2016. "An Amphiprotic Novel Chitosanase from Bacillus mycoides and Its Application in the Production of Chitooligomers with Their Antioxidant and Anti-Inflammatory Evaluation" International Journal of Molecular Sciences 17, no. 8: 1302. https://doi.org/10.3390/ijms17081302
APA StyleLiang, T. -W., Chen, W. -T., Lin, Z. -H., Kuo, Y. -H., Nguyen, A. D., Pan, P. -S., & Wang, S. -L. (2016). An Amphiprotic Novel Chitosanase from Bacillus mycoides and Its Application in the Production of Chitooligomers with Their Antioxidant and Anti-Inflammatory Evaluation. International Journal of Molecular Sciences, 17(8), 1302. https://doi.org/10.3390/ijms17081302