
Metabolic Effects of Berries with Structurally Diverse Anthocyanins



Abstract
:
1. Introduction
2. Results
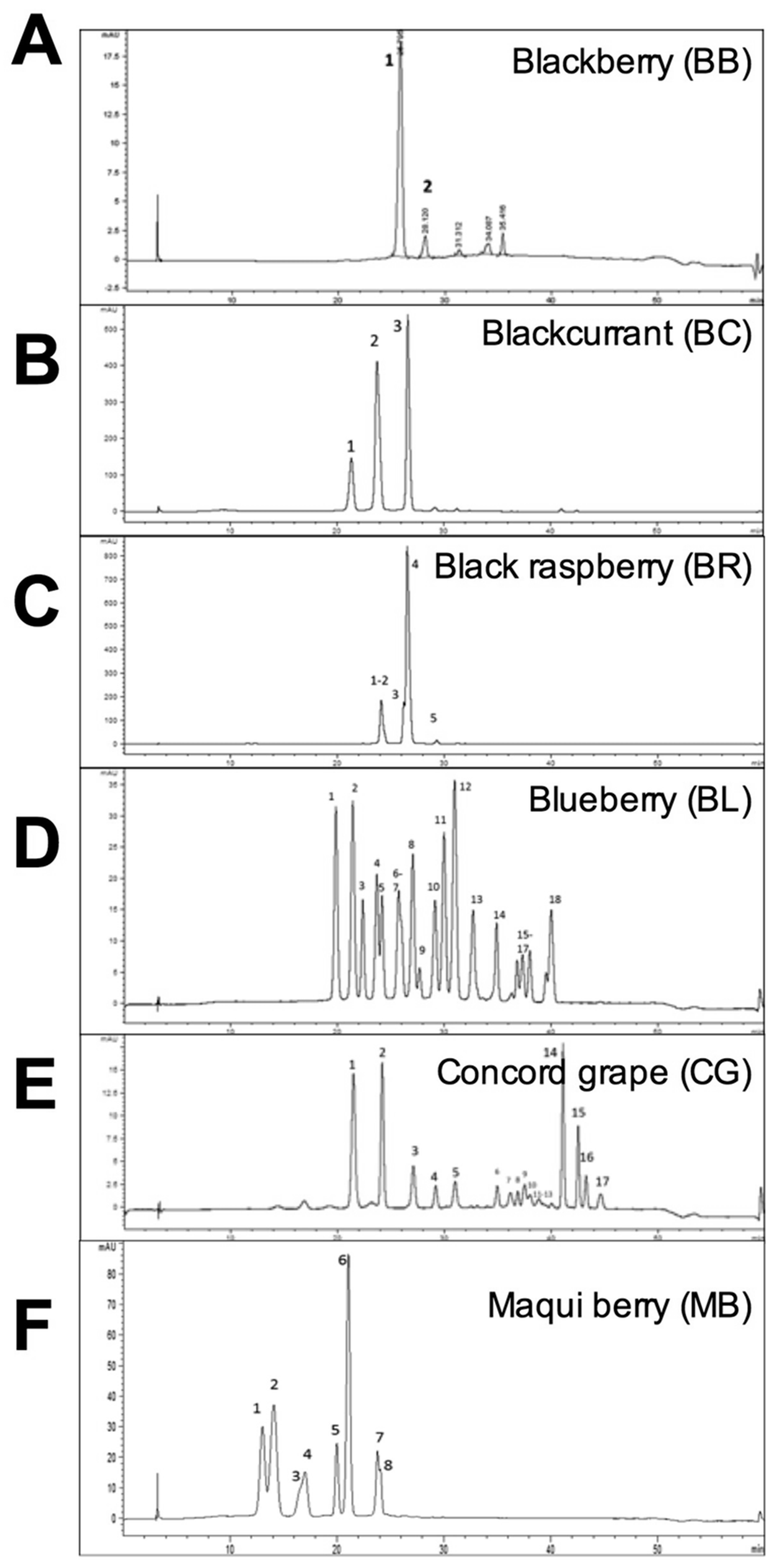
2.1. Characterization of Whole Berry Anthocyanins
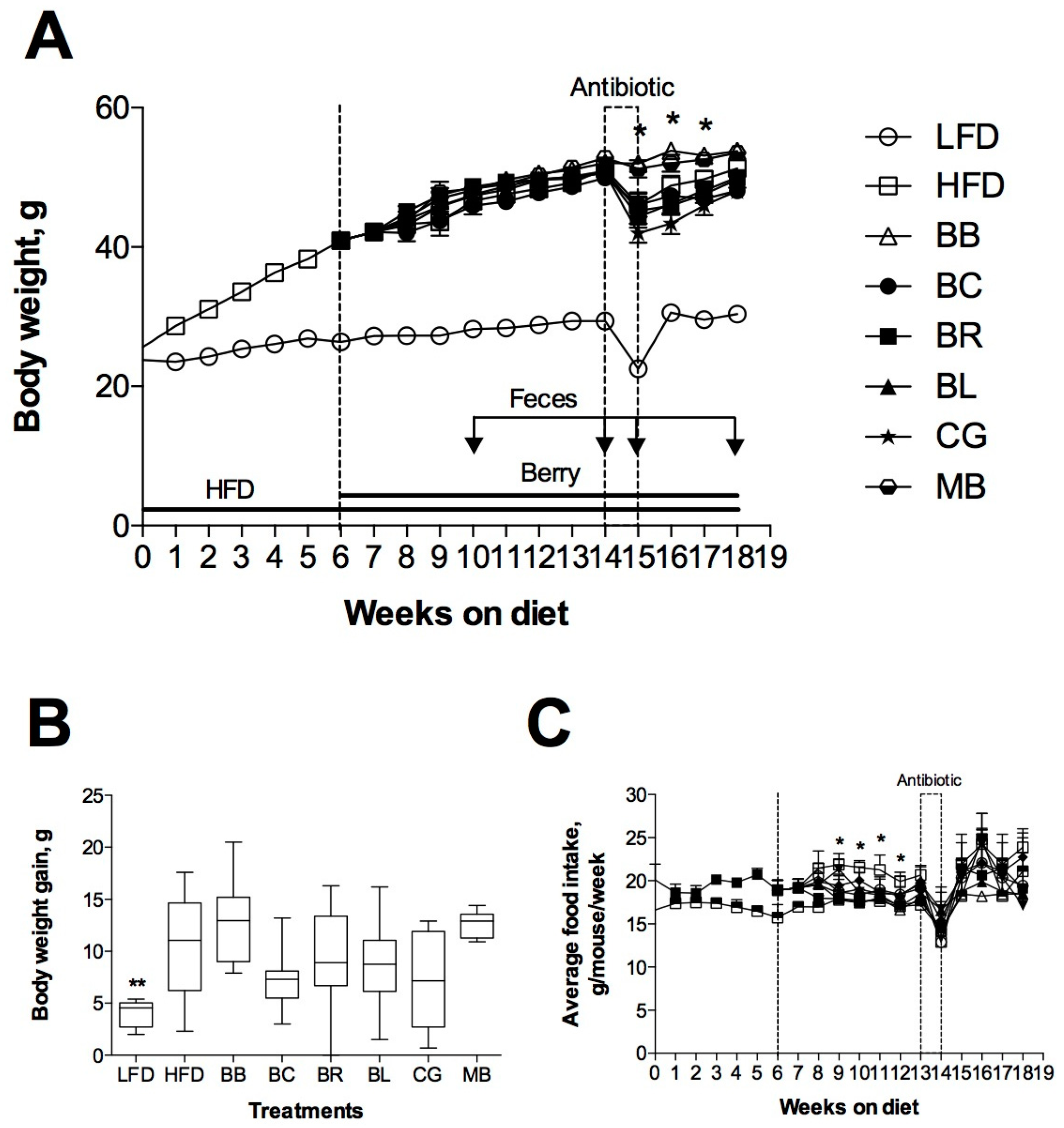
2.2. Changes of Body Weight and Food Intake
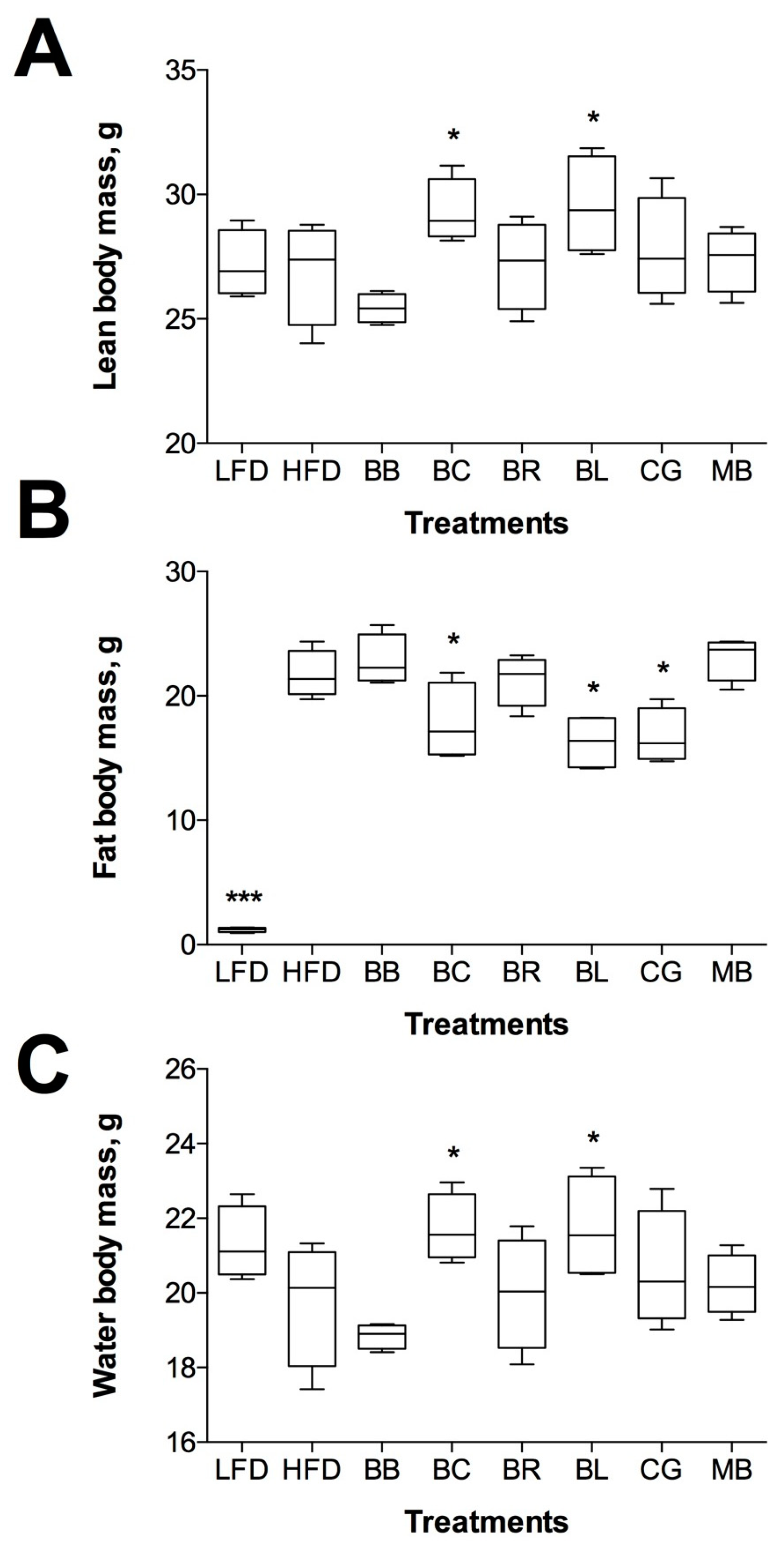
2.3. Changes in Body Composition
2.4. Anthocyanins in Fecal Samples
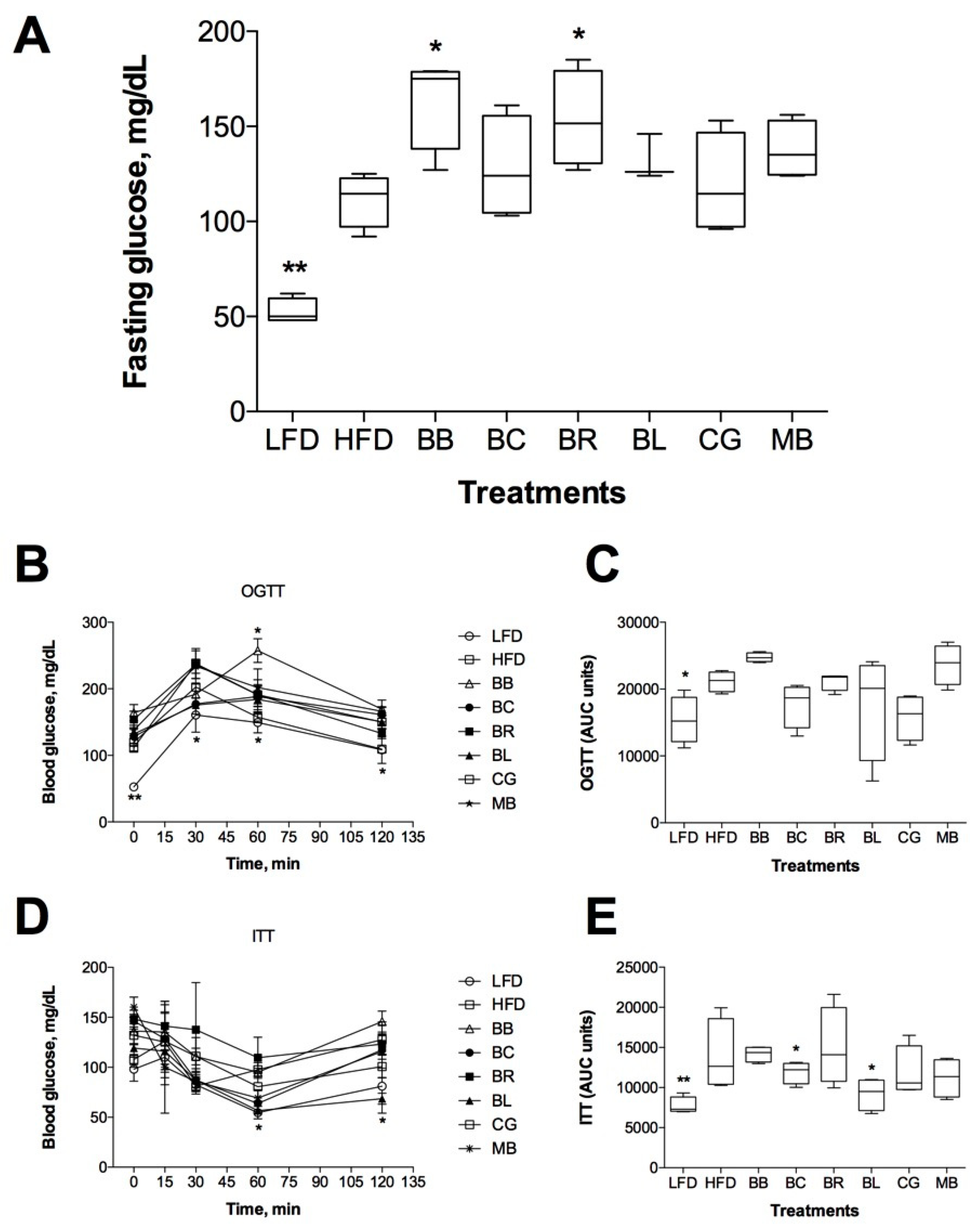
2.5. Effect on Glucose Metabolism and Insulin Sensitivity
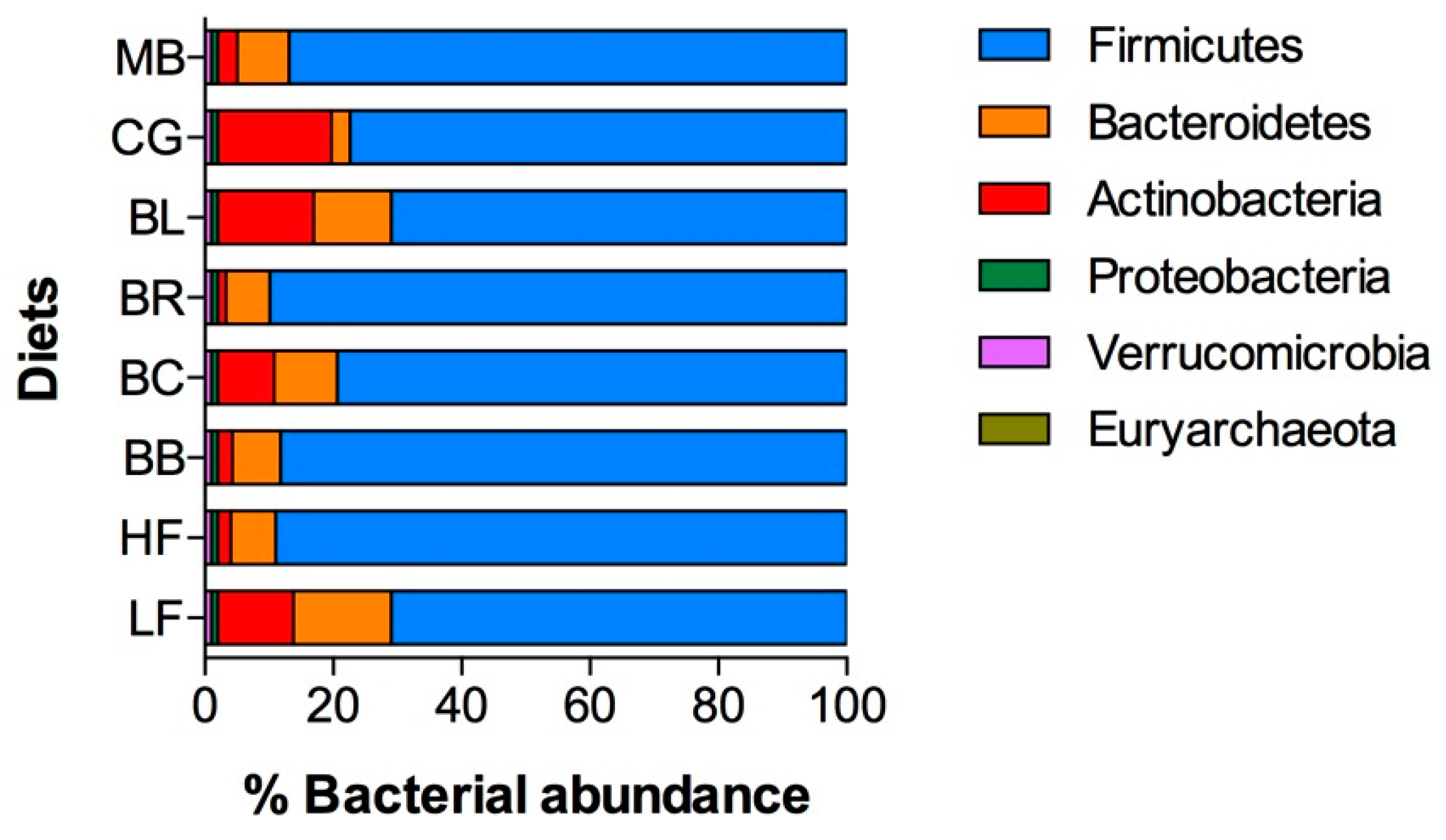
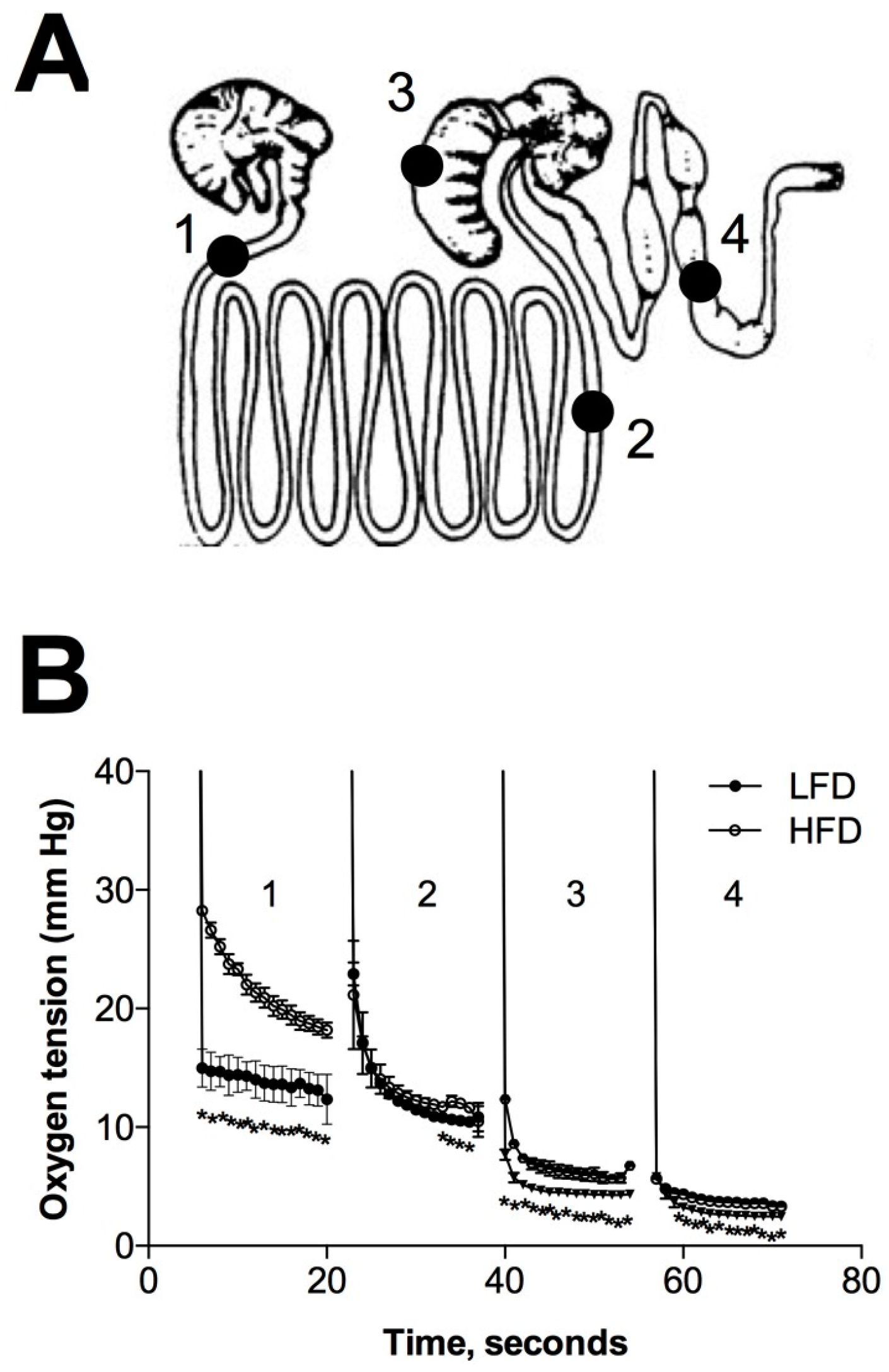
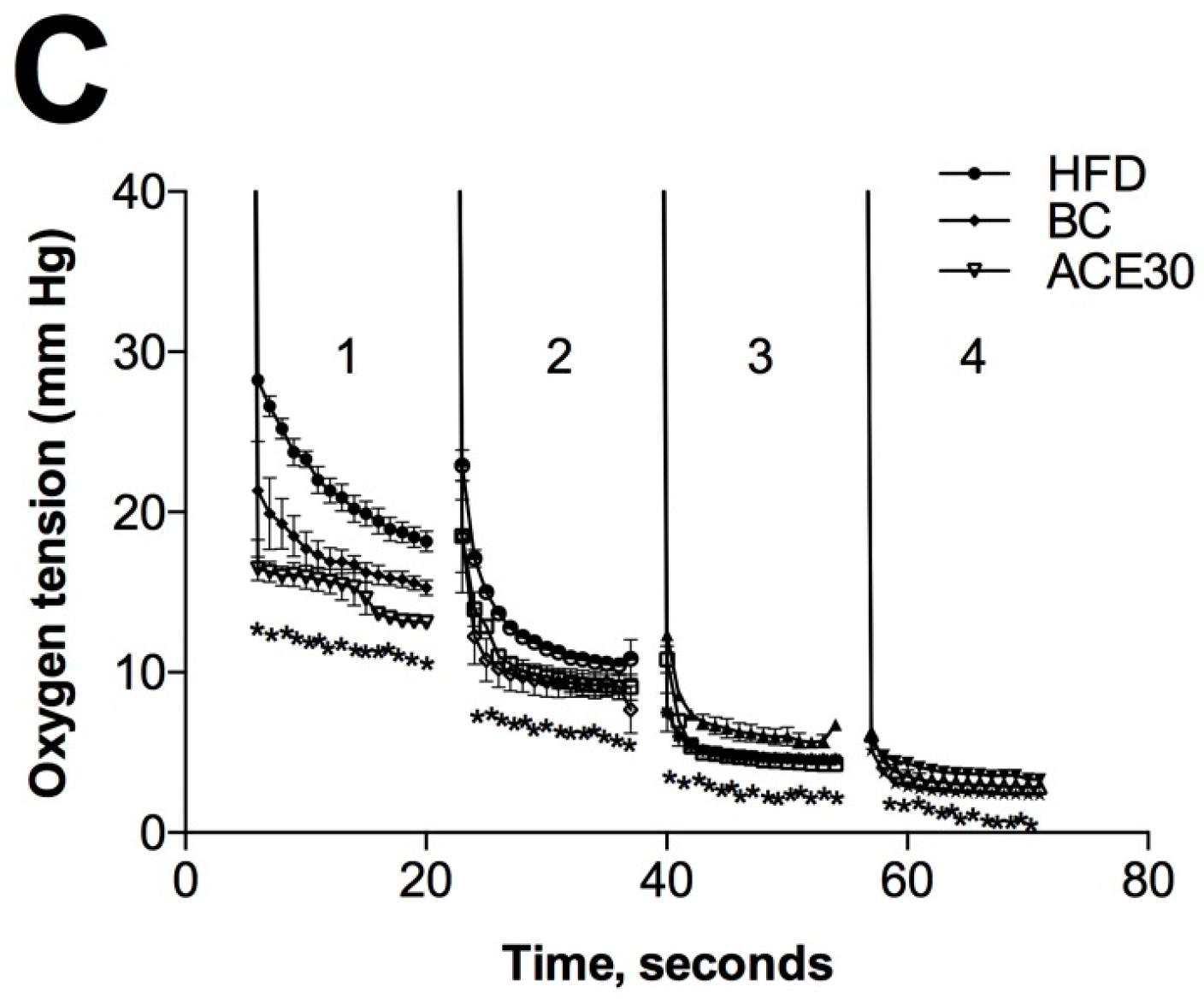
2.6. Changes in Microbiome and Gastrointestinal Lumen Oxygen
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Diets
4.3. Antibiotic Knockdown of Endogenous Gut Microbiome
4.4. Oral Glucose and Insulin Tolerance Tests
4.5. Sample Collection and Oxygen Measurements
4.6. Gastrointestinal Microbial Profiles
4.7. Anthocyanin Extraction and Quantification
4.8. HPLC Analysis of Anthocyanins
4.9. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Manco, M.; Putignani, L.; Bottazzo, G.F. Gut microbiota, lipopolysaccharides, and innate immunity in the pathogenesis of obesity and cardiovascular risk. Endocr. Rev. 2010, 31, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T. Recent Progress in Anti-Obesity and Anti-Diabetes Effect of Berries. Antioxidants 2016, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Jennings, A.; Welch, A.A.; Spector, T.; Macgregor, A.; Cassidy, A. Intakes of anthocyanins and flavones are associated with biomarkers of insulin resistance and inflammation in women. J. Nutr. 2014, 144, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Chen, G.; Liao, D.; Zhu, Y.; Xue, X. Effects of Berries Consumption on Cardiovascular Risk Factors: A Meta-analysis with Trial Sequential Analysis of Randomized Controlled Trials. Sci. Rep. 2016, 6, 23625. [Google Scholar] [CrossRef] [PubMed]
- Stull, A.J.; Cash, K.C.; Johnson, W.D.; Champagne, C.M.; Cefalu, W.T. Bioactives in blueberries improve insulin sensitivity in obese, insulin-resistant men and women. J. Nutr. 2010, 140, 1764–1768. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic patients. J. Nutr. 2015, 145, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, R.M.; Naidoo, N.; Landberg, R. Dietary flavonoids and the development of type 2 diabetes and cardiovascular diseases: Review of recent findings. Curr. Opin. Lipidol. 2013, 24, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, A.; Retterstol, L.; Laake, P.; Paur, I.; Bohn, S.K.; Sandvik, L.; Blomhoff, R. Anthocyanins inhibit nuclear factor-κB activation in monocytes and reduce plasma concentrations of pro-inflammatory mediators in healthy adults. J. Nutr. 2007, 137, 1951–1954. [Google Scholar] [PubMed]
- Zhu, Y.; Xia, M.; Yang, Y.; Liu, F.; Li, Z.; Hao, Y.; Mi, M.; Jin, T.; Ling, W. Purified anthocyanin supplementation improves endothelial function via NO-cGMP activation in hypercholesterolemic individuals. Clin. Chem. 2011, 57, 1524–1533. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.S.; Marckmann, P.; Dragsted, L.O.; Finne Nielsen, I.L.; Nielsen, S.E.; Gronbaek, M. Effect of red wine and red grape extract on blood lipids, haemostatic factors, and other risk factors for cardiovascular disease. Eur. J. Clin. Nutr. 2005, 59, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Curtis, P.J.; Kroon, P.A.; Hollands, W.J.; Walls, R.; Jenkins, G.; Kay, C.D.; Cassidy, A. Cardiovascular disease risk biomarkers and liver and kidney function are not altered in postmenopausal women after ingesting an elderberry extract rich in anthocyanins for 12 weeks. J. Nutr. 2009, 139, 2266–2271. [Google Scholar] [CrossRef] [PubMed]
- Giordano, L.; Coletta, W.; Tamburrelli, C.; D’Imperio, M.; Crescente, M.; Silvestri, C.; Rapisarda, P.; Reforgiato Recupero, G.; de Curtis, A.; Iacoviello, L.; et al. Four-week ingestion of blood orange juice results in measurable anthocyanin urinary levels but does not affect cellular markers related to cardiovascular risk: A randomized cross-over study in healthy volunteers. Eur. J. Nutr. 2012, 51, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Wright, O.R.; Netzel, G.A.; Sakzewski, A.R. A randomized, double-blind, placebo-controlled trial of the effect of dried purple carrot on body mass, lipids, blood pressure, body composition, and inflammatory markers in overweight and obese adults: The QUENCH trial. Can. J. Physiol. Pharmacol. 2013, 91, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black Currant Anthocyanins Attenuate Weight Gain and Improve Glucose Metabolism in Diet-Induced Obese Mice with Intact, but Not Disrupted, Gut Microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T. Dietary anthocyanin-rich plants: Biochemical basis and recent progress in health benefits studies. Mol. Nutr. Food Res. 2012, 56, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human metabolism and elimination of the anthocyanin, cyanidin-3-glucoside: A (13)C-tracer study. Am. J. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, A.; Fernandes, I.; Norberto, S.; Mateus, N.; Calhau, C. Interplay between Anthocyanins and Gut Microbiota. J. Agric. Food Chem. 2014, 62, 6898–6902. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X. Anthocyanins: Structural characteristics that result in unique metabolic patterns and biological activities. Free Radic. Res. 2006, 40, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, S.; Rudloff, S.; Asseburg, H.; Borsch, C.; Frohling, B.; Unger, F.; Dold, S.; Spengler, B.; Rompp, A.; Kunz, C. Uptake and bioavailability of anthocyanins and phenolic acids from grape/blueberry juice and smoothie in vitro and in vivo. Br. J. Nutr. 2015, 113, 1044–1055. [Google Scholar] [CrossRef] [PubMed]
- Charron, C.S.; Kurilich, A.C.; Clevidence, B.A.; Simon, P.W.; Harrison, D.J.; Britz, S.J.; Baer, D.J.; Novotny, J.A. Bioavailability of anthocyanins from purple carrot juice: Effects of acylation and plant matrix. J. Agric. Food Chem. 2009, 57, 1226–1230. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, A.; Licht, T.R.; Wilcks, A.; Andersen, J.B.; Schmidt, L.R.; Gronlund, H.A.; Vigsnaes, L.K.; Michaelsen, K.F.; Bahl, M.I. Introducing GUt low-density array (GULDA): A validated approach for qPCR-based intestinal microbial community analysis. FEMS Microbiol. Lett. 2012, 337, 38–47. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Magnuson, B.A.; Giusti, M.M. Analysis of anthocyanins in rat intestinal contents--impact of anthocyanin chemical structure on fecal excretion. J. Agric. Food Chem. 2005, 53, 2859–2866. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Durst, R.W.; Wrolstad, R.E. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [PubMed]
- Zhang, L.; Zhou, J.; Liu, H.; Khan, M.A.; Huang, K.; Gu, Z. Compositions of anthocyanins in blackberry juice and their thermal degradation in relation to antioxidant activity. Eur. Food Res. Technol. 2012, 235, 637–645. [Google Scholar] [CrossRef]
- Dossett, M.; Lee, J.; Finn, C.E. Variation in anthocyanins and total phenolics of black raspberry populations. J. Funct. Foods 2010, 2, 292–297. [Google Scholar] [CrossRef]
- Fredes, C.; Yousef, G.G.; Robert, P.; Grace, M.H.; Lila, M.A.; Gomez, M.; Gebauer, M.; Montenegro, G. Anthocyanin profiling of wild maqui berries (Aristotelia chilensis [Mol.] Stuntz) from different geographical regions in Chile. J. Sci. Food Agric. 2014, 94, 2639–2648. [Google Scholar] [CrossRef] [PubMed]
- Slimestad, R.; Solheim, H. Anthocyanins from black currants (Ribes nigrum L.). J. Agric. Food Chem. 2002, 50, 3228–3231. [Google Scholar] [CrossRef] [PubMed]
- Yousef, G.G.; Brown, A.F.; Funakoshi, Y.; Mbeunkui, F.; Grace, M.H.; Ballington, J.R.; Loraine, A.; Lila, M.A. Efficient quantification of the health-relevant anthocyanin and phenolic acid profiles in commercial cultivars and breeding selections of blueberries (Vaccinium spp.). J. Agric. Food Chem. 2013, 61, 4806–4815. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Race, E.J.; Shrikhande, A.J. Characterization of anthocyanins in grape juices by ion trap liquid chromatography-mass spectrometry. J. Agric. Food Chem. 2003, 51, 1839–1844. [Google Scholar] [CrossRef] [PubMed]
- Mbeunkui, F.; Grace, M.H.; Yousef, G.G.; Lila, M.A. Isolation and characterization of flavonols from blackcurrant by high-performance counter-current chromatography and electrospray ionization tandem mass spectrometry. J. Sep. Sci. 2012, 35, 1682–1689. [Google Scholar] [CrossRef] [PubMed]
- McGhie, T.K.; Ainge, G.D.; Barnett, L.E.; Cooney, J.M.; Jensen, D.J. Anthocyanin glycosides from berry fruit are absorbed and excreted unmetabolized by both humans and rats. J. Agric. Food Chem. 2003, 51, 4539–4548. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.; Hollman, P.C.; Katan, M.B.; Kromhout, D. Intake of potentially anticarcinogenic flavonoids and their determinants in adults in The Netherlands. Nutr. Cancer 1993, 20, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wilkes, S.; Rogers, T.; Khanal, R.C.; Wu, X.; Hager, T.J.; Hager, A.; Howard, L. Dietary black raspberry anthocyanins do not alter development of obesity in mice fed an obesogenic high-fat diet. J. Agric. Food Chem. 2010, 58, 3977–3983. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.Y.; Dragano, N.R.; Lenquiste, S.A.; Batista, A.G.; Palazzo, C.C.; Marostica, M.R., Jr. Freeze-dried jaboticaba peel powder rich in anthocyanins did not reduce weight gain and lipid content in mice and rats. Archiv. Latinoam. Nutr. 2012, 62, 37–43. [Google Scholar]
- Tsuda, T.; Horio, F.; Uchida, K.; Aoki, H.; Osawa, T. Dietary cyanidin 3-O-β-d-glucoside-rich purple corn color prevents obesity and ameliorates hyperglycemia in mice. J. Nutr. 2003, 133, 2125–2130. [Google Scholar] [PubMed]
- Kwon, S.H.; Ahn, I.S.; Kim, S.O.; Kong, C.S.; Chung, H.Y.; Do, M.S.; Park, K.Y. Anti-obesity and hypolipidemic effects of black soybean anthocyanins. J. Med. Food 2007, 10, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.P.; Choi, J.H.; Han, E.H.; Kim, H.G.; Wee, J.H.; Jung, K.O.; Jung, K.H.; Kwon, K.I.; Jeong, T.C.; Chung, Y.C.; et al. Purple sweet potato anthocyanins attenuate hepatic lipid accumulation through activating adenosine monophosphate-activated protein kinase in human HepG2 cells and obese mice. Nutr. Res. 2011, 31, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Qi, X.; Liu, Y.; Guo, J.; Zhu, R.; Chen, W.; Zheng, X.; Yu, T. Dietary supplementation with purified mulberry (Morus australis Poir) anthocyanins suppresses body weight gain in high-fat diet fed C57BL/6 mice. Food Chem. 2013, 141, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Tang, Q.; Yu, Z.; Gao, Z.; Hu, H.; Chen, W.; Zheng, X.; Yu, T. Inhibitory effects of sweet cherry anthocyanins on the obesity development in C57BL/6 mice. Int. J. Food Sci. Nutr. 2014, 65, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Anderson, R.A. An extract of chokeberry attenuates weight gain and modulates insulin, adipogenic and inflammatory signalling pathways in epididymal adipose tissue of rats fed a fructose-rich diet. Br. J. Nutr. 2012, 108, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Xia, M.; Zou, T.; Ling, W.; Zhong, R.; Zhang, W. Cyanidin 3-glucoside attenuates obesity-associated insulin resistance and hepatic steatosis in high-fat diet-fed and db/db mice via the transcription factor FoxO1. J. Nutr. Biochem. 2012, 23, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Rojo, L.E.; Ribnicky, D.; Logendra, S.; Poulev, A.; Rojas-Silva, P.; Kuhn, P.; Dorn, R.; Grace, M.H.; Lila, M.A.; Raskin, I. In Vitro and in Vivo Anti-Diabetic Effects of Anthocyanins from Maqui Berry (Aristotelia chilensis). Food Chem. 2012, 131, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Benn, T.; Kim, B.; Park, Y.K.; Wegner, C.J.; Harness, E.; Nam, T.G.; Kim, D.O.; Lee, J.S.; Lee, J.Y. Polyphenol-rich blackcurrant extract prevents inflammation in diet-induced obese mice. J. Nutr. Biochem. 2014, 25, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Ichiyanagi, T.; Shida, Y.; Rahman, M.M.; Hatano, Y.; Konishi, T. Bioavailability and tissue distribution of anthocyanins in bilberry (Vaccinium myrtillus L.) extract in rats. J. Agric. Food Chem. 2006, 54, 6578–6587. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Feng, Q.; Liang, S.; Sonne, S.B.; Xia, Z.; Qiu, X.; Li, X.; Long, H.; Zhang, J.; Zhang, D.; et al. A catalog of the mouse gut metagenome. Nat. Biotechnol. 2015, 33, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.S.; Barbehennb, R.V. Oxygen levels in the gut lumens of herbivorous insects. J. Insect. Physiol. 2000, 46, 897–903. [Google Scholar] [CrossRef]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H.; et al. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology 2014, 147, 1055–1063. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Berry | Anthocyanins (Total, mg/g Dry Weight) | Individual Anthocyanins | Percent % (Total Anthocyanins) |
|---|---|---|---|
| Blackberry (BB) | 9.42 ± 0.03 | 1. Cyanidin-3-O-glucoside | 77.58 |
| 2. Cyanidin-3-O-rutinoside | 6.90 | ||
| 3. Non-identified | trace | ||
| 4. Cyanidin-3-O-(malonyl)glucoside | 15.52 | ||
| 5. Cyanidin-3-O-(dioxalyl)glucoside | trace | ||
| Black currant (BC) | 16.42 ± 0.24 | 1. Delphinidin-3-O-glucoside | 14.61 |
| 2. Delphinidin-3-O-rutinoside | 43.92 | ||
| 3. Cyanidin-3-O-rutinoside | 39.54 | ||
| 4. Cyanidin-3-O-glucoside | 2.01 | ||
| Black raspberry (BR) | 24.75 ± 1.19 | 1. Cyanidin-3-O-sambubioside | 8.61 |
| 2. Cyanidin-3-O-glucoside | 8.62 | ||
| 3. Cyanidin-3-O-(xylosyl)rutinoside | 73.62 | ||
| 4. Cyanidin-3-O-rutinoside | 8.18 | ||
| 5. Pelargonidin-3-O-rutinoside | 1.13 | ||
| Blueberry (BL) | 9.33 ± 0.26 | 1. Delphinidin-3-O-galactoside | 9.21 |
| 2. Delphinidin-3-O-glucoside | 10.22 | ||
| 3. Cyanidin-3-O-galactoside | 4.12 | ||
| 4. Delphinidin-3-O-arabinoside | 6.03 | ||
| 5. Cyanidin-3-O-glucoside | 4.34 | ||
| 6. Petunidin-3-O-galactoside | 7.72 | ||
| 7. Cyanidin-3-O-arabinoside | 2.63 | ||
| 8. Petunidin-3-O-glucoside | 7.51 | ||
| 9. Peonidin-3-O-galactoside | 1.44 | ||
| 10. Petunidin-3-O-arabinoside | 5.33 | ||
| 11. Malvidin-3-O-galactoside | 8.81 | ||
| 12. Malvidin-3-O-glucoside | 12.70 | ||
| 13. Malvidin-3-O-arabinoside | 5.12 | ||
| 14. Delphinidin-3-O-(acetyl)glucoside | 3.54 | ||
| 15. Cyanidin-3-O-(acetyl)glucoside | 1.43 | ||
| 16. Malvidin-3-O-(acetyl)galactoside | 2.11 | ||
| 17. Petunidin-3-O-(acetyl)glucoside | 2.24 | ||
| 18. Malvidin-3-O-(acetyl)glucoside | 4.83 | ||
| Concord grape (CG) | 2.37 ± 0.03 | 1. Delphinidin-3-O-glucoside | 13.61 |
| 2. Cyanidin-3-O-glucoside | 12.01 | ||
| 3. Petunidin-3-O-glucoside | 7.41 | ||
| 4. Peonidin-3-O-glucoside | 5.92 | ||
| 5. Malvidin-3-O-glucoside | 6.42 | ||
| 6. Delphinidin-3-O-(acetyl)glucoside | 5.75 | ||
| 7. Delphinidin-3,5-O-(coumaroyl)diglucoside | 5.60 | ||
| 8. Cyanidin-3-O-(acetyl)glucoside | 5.31 | ||
| 9. Cyanidin-3,5-O-(coumaroyl)diglucoside | 6.55 | ||
| 10. Petunidin-3-O-(acetyl)glucoside | trace | ||
| 11. Malvidin-3,5-O-(coumaroyl)diglucoside | trace | ||
| 12. Peonidin-3,5-O-(coumaroyl)diglucoside | trace | ||
| 13. Peonidin-3-O-(acetyl)glucoside | trace | ||
| 14. Delphinidin-3,5-O-(coumaroyl)diglucoside | 11.36 | ||
| 15. Cyanidin-3-O-(coumaroyl)glucoside | 8.05 | ||
| 16. Petunidin-3-O-(coumaroyl)glucoside | 6.24 | ||
| 17. Peonidin-3-O-(coumaroyl)glucoside | 5.77 | ||
| 18. Malvidin-3-O-(coumaroyl)glucoside | trace | ||
| Maqui berry (MB) | 10.95 ± 0.12 | 1. Delphinidin-3-O-sambubioside-5-O-glucoside | 15.36 |
| 2. Delphinidin-3,5-O-diglucoside | 23.91 | ||
| 3. Cyanidin-3,5-O-diglucoside | trace | ||
| 4. Cyanidin-3-O-sambubioside-5-O-glucoside | 11.32 | ||
| 5. Delphinidin-3-O-sambubioside | 7.76 | ||
| 6. Delphinidin-3-O-glucoside | 31.26 | ||
| 7. Cyanidin-3-O-glucoside | 6.86 | ||
| 8. Cyanidin-3-O-sambubioside | 3.53 |
| Berry | Anthocyanidins (Aglycones) | Glycosylation Ratio (Di/Mono) | Acetylation Ratio (Yes/No) | ||||
|---|---|---|---|---|---|---|---|
| Cy | De | Mv | Pt | Peo | |||
| Blackberry (BB) | 100 | 0 | 0 | 0 | 0 | 7/93 | 16/84 |
| Black currant (BC) | 42 | 58 | 0 | 0 | 0 | 84/16 | 0/100 |
| Black raspberry (BR) | 99 | 0 | 0 | 0 | 0 | 91/9 | 0/100 |
| Blueberry (BL) | 12 | 29 | 34 | 23 | 2 | 0/100 | 13/87 |
| Concord grape (CG) | 32 | 37 | 7 | 19 | 7 | 23/77 | 54/46 |
| Maqui berry (MB) | 21 | 78 | 0 | 0 | 0 | 62/38 | 0/100 |
| Diet * | Time on Diet, Weeks | |||
|---|---|---|---|---|
| 4 | 8 | 9 (1 Week Antibiotic) | 12 | |
| HFD | nd | nd | nd | nd |
| Blackberry (BB) | 2.37 ± 0.08 | 3.48 ± 0.10 | 8.51 ± 0.18 | 4.83 ± 1.13 |
| Black currant (BC) | 26.35 ± 1.25 | 34.26 ± 0.14 | 326.82 ± 31.89 | 48.17 ± 2.74 |
| Black raspberry (BR) | 44.93 ± 6.64 | 62.10 ± 9.42 | 120.97 ± 2.13 | 83.93 ± 1.30 |
| Blueberry (BL) | 5.30 ± 0.66 | 17.05 ± 3.10 | 46.57 ± 6.00 | 18.57 ± 0.91 |
| Concord grape (CG) | 9.24 ± 5.13 | 22.41 ± 3.74 | 260.58 ± 10.46 | 34.26 ± 0.60 |
| Maqui berry (MB) | 30.38 ± 1.18 | 31.22 ± 1.37 | 339.32 ± 52.69 | 172.16 ± 5.44 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Overall, J.; Bonney, S.A.; Wilson, M.; Beermann, A., III; Grace, M.H.; Esposito, D.; Lila, M.A.; Komarnytsky, S. Metabolic Effects of Berries with Structurally Diverse Anthocyanins. Int. J. Mol. Sci. 2017, 18, 422. https://doi.org/10.3390/ijms18020422
Overall J, Bonney SA, Wilson M, Beermann A III, Grace MH, Esposito D, Lila MA, Komarnytsky S. Metabolic Effects of Berries with Structurally Diverse Anthocyanins. International Journal of Molecular Sciences. 2017; 18(2):422. https://doi.org/10.3390/ijms18020422
Chicago/Turabian StyleOverall, John, Sierra A. Bonney, Mickey Wilson, Arnold Beermann, III, Mary H. Grace, Debora Esposito, Mary Ann Lila, and Slavko Komarnytsky. 2017. "Metabolic Effects of Berries with Structurally Diverse Anthocyanins" International Journal of Molecular Sciences 18, no. 2: 422. https://doi.org/10.3390/ijms18020422
APA StyleOverall, J., Bonney, S. A., Wilson, M., Beermann, A., III, Grace, M. H., Esposito, D., Lila, M. A., & Komarnytsky, S. (2017). Metabolic Effects of Berries with Structurally Diverse Anthocyanins. International Journal of Molecular Sciences, 18(2), 422. https://doi.org/10.3390/ijms18020422