
Identification of Differentially Expressed Micrornas Associate with Glucose Metabolism in Different Organs of Blunt Snout Bream (Megalobrama amblycephala)

 ,
, 

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
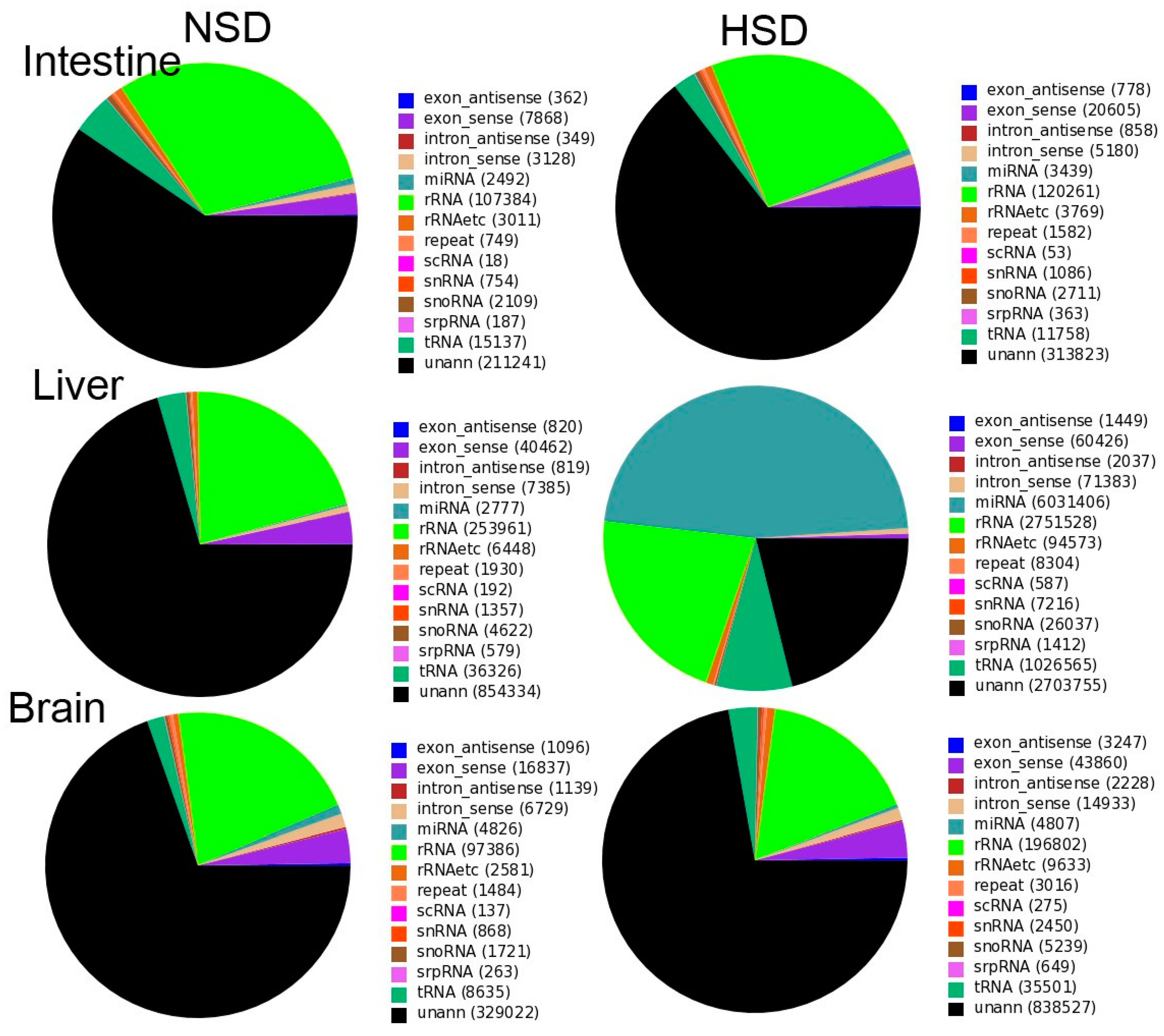
2.1. Small RNA Profiles in M. amblycephala
2.2. Identification of Known miRNAs
2.3. Identification of Novel miRNAs
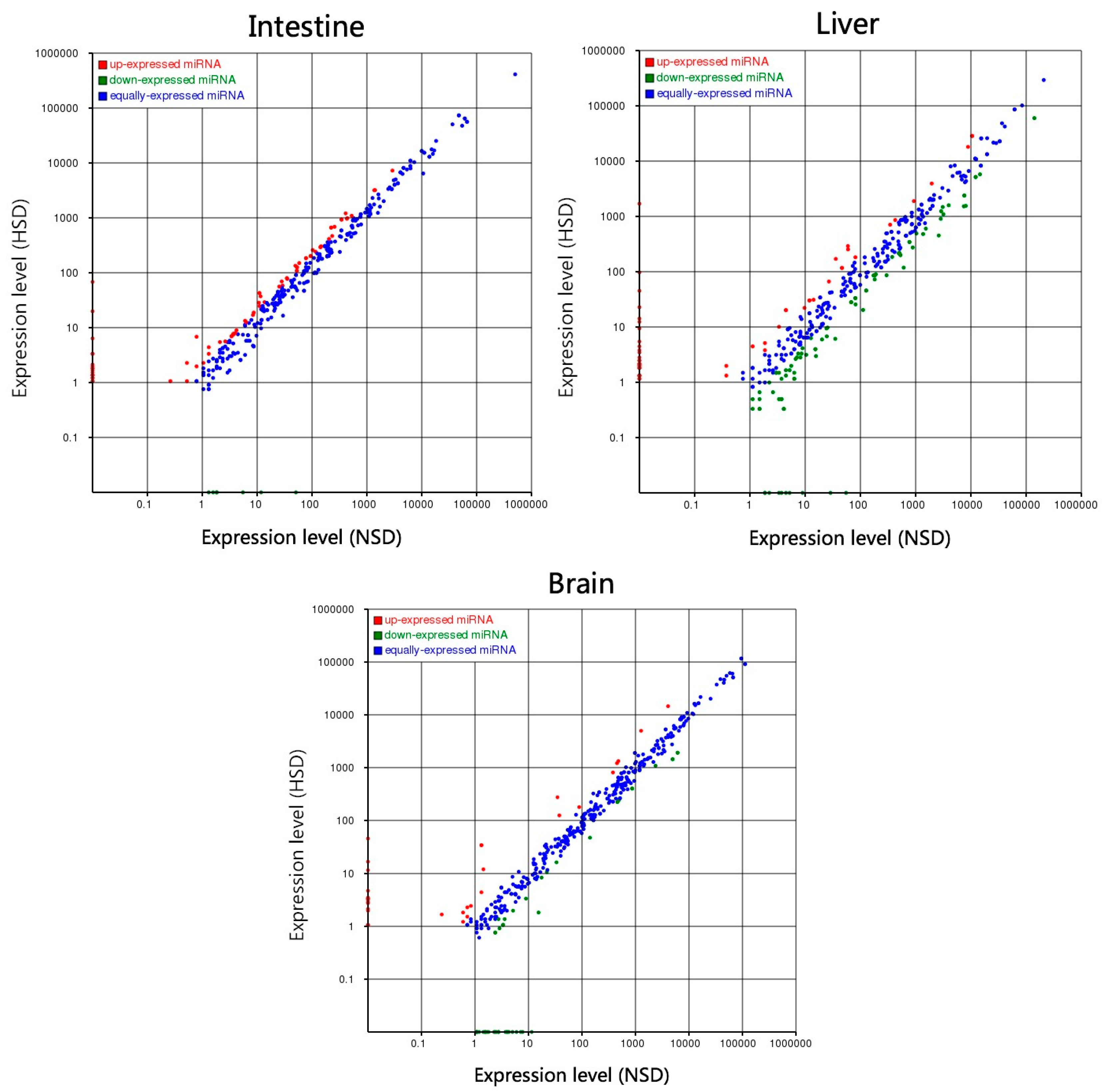
2.4. Identification of DEMs
2.5. Prediction of Potential Targets of DEMs
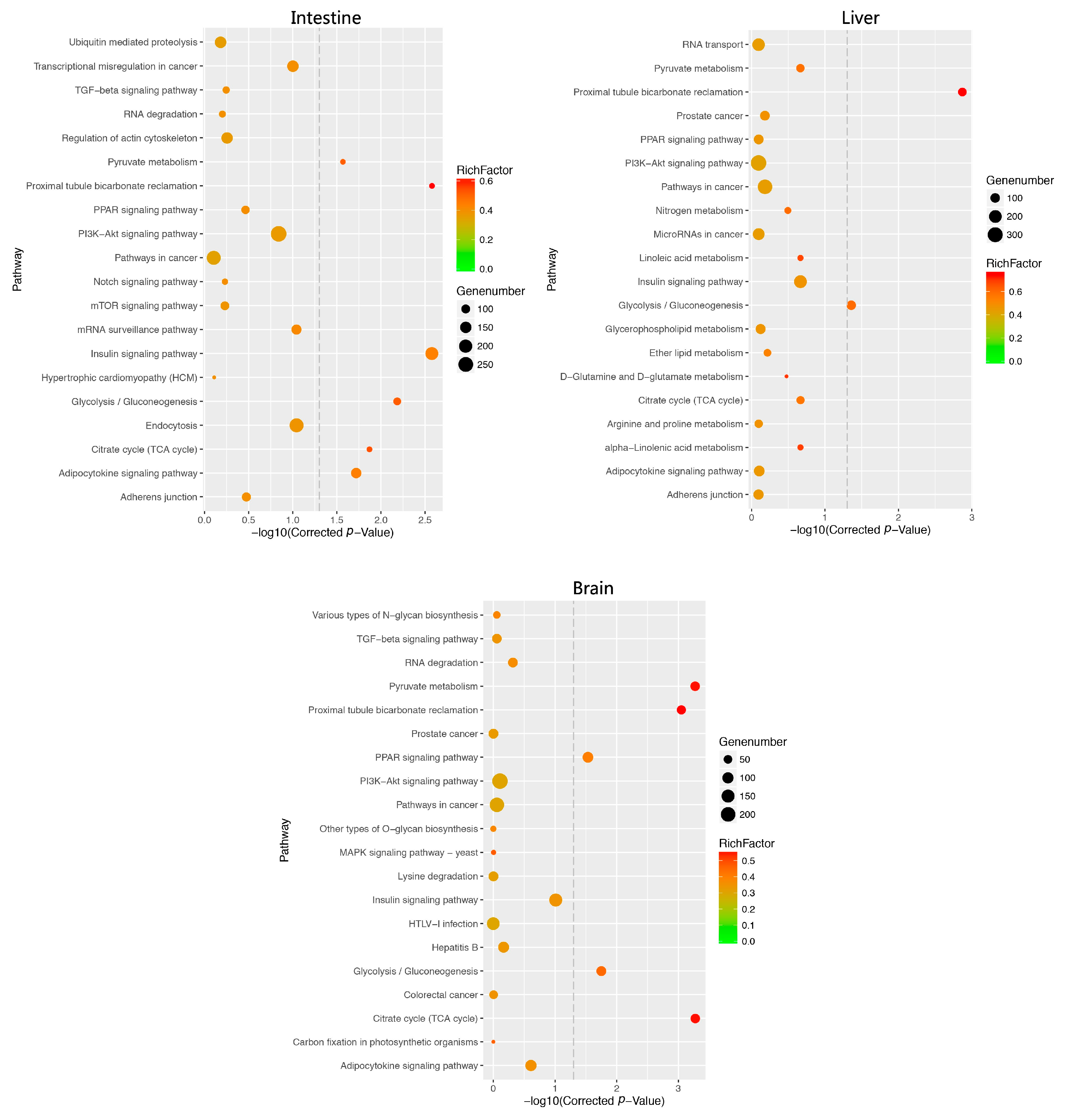
2.6. Function Analysis of DEM Target Genes
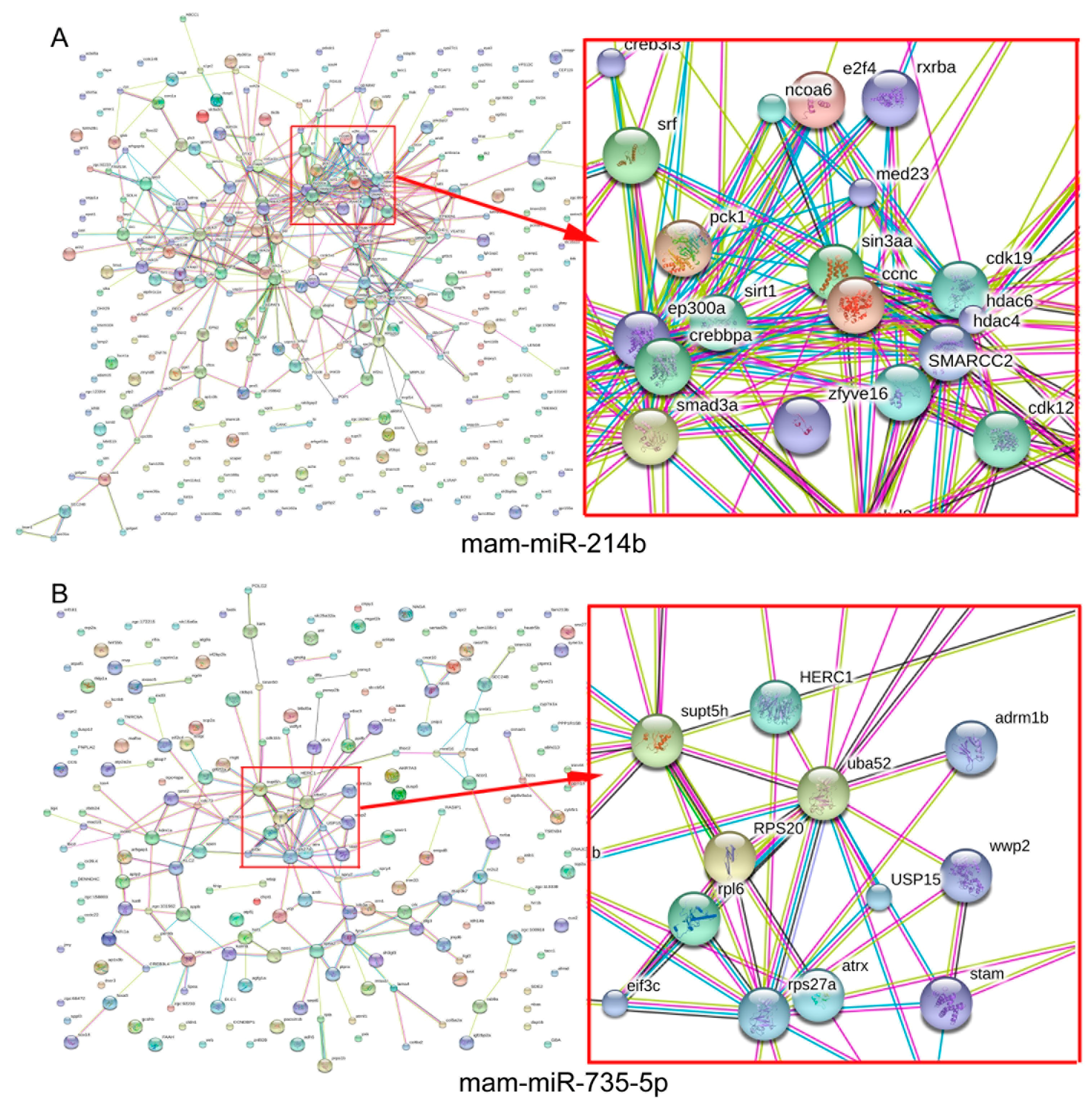
2.7. PPI (Protein–Protein Internetwork) and Interaction Analysis
3. Discussion
4. Materials and Methods
4.1. Fish and Experimental Procedures
4.2. Small RNA Library Preparation and Sequencing
4.3. Sequencing Analysis for miRNA Profiling
4.4. Data Normalization, Processing and Identification of Differential Expressed miRNAs (DEMs)
4.5. Target Prediction for DEMs
4.6. Enrichment Analysis for DEM Targets
4.7. MiRNA-mRNA Regulatory Network and PPI (Protein–Protein Internetwork) and the Module Analysis
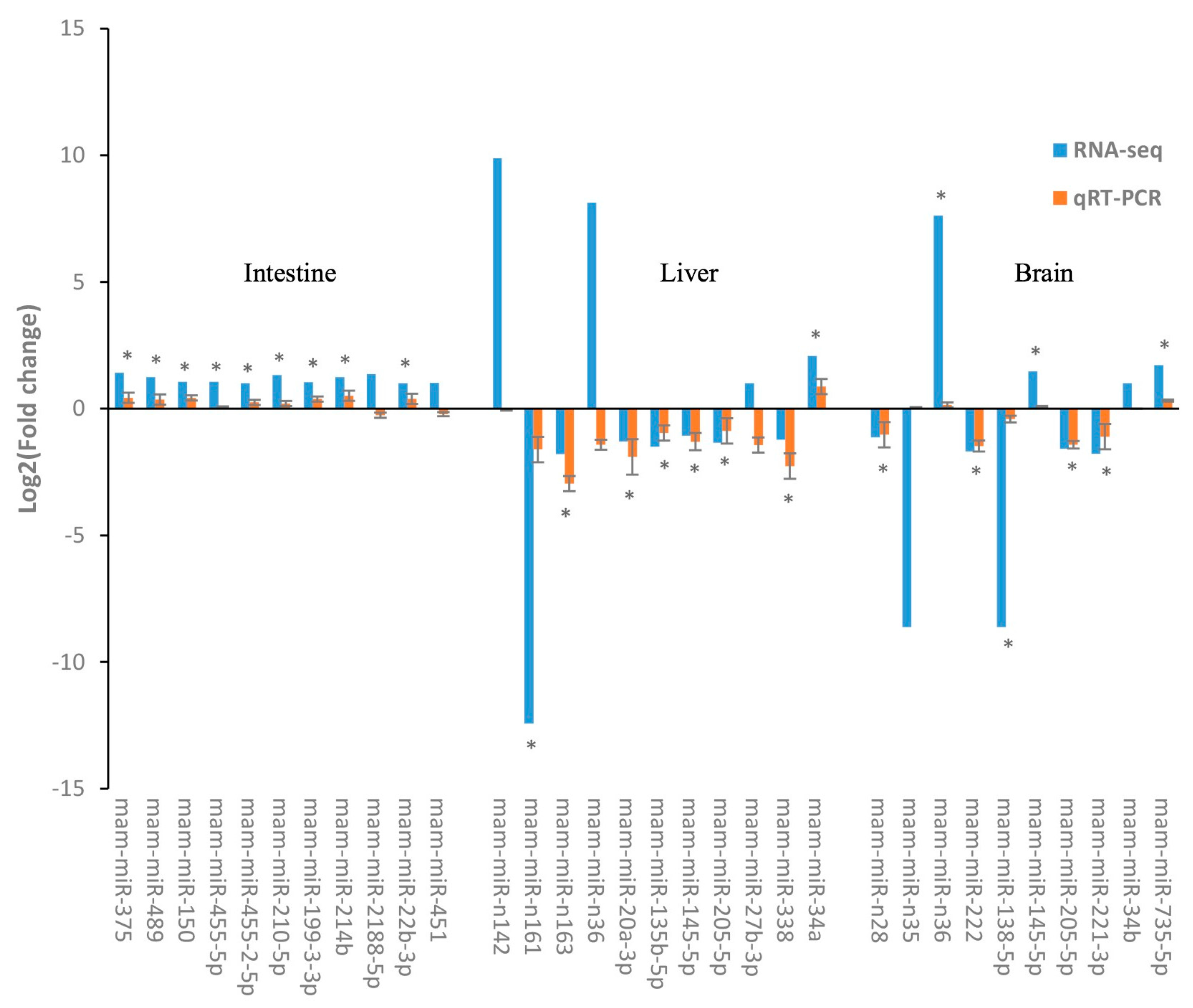
4.8. Quantitative Real-Time PCR (qPCR) Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- National Research Council, Division on Earth and Life Studies, Board on Agriculture and Natural Resources, Committee on the Nutrient Requirements of Fish and Shrimp. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Li, X.; Liu, W.; Jiang, Y.; Zhu, H.; Ge, X. Effects of dietary protein and lipid levels in practical diets on growth performance and body composition of blunt snout bream (Megalobrama amblycephala) fingerlings. Aquaculture 2010, 303, 65–70. [Google Scholar] [CrossRef]
- Li, X.F.; Wang, Y.; Liu, W.B.; Jiang, G.Z.; Zhu, J. Effects of dietary carbohydrate/lipid ratios on growth performance, body composition and glucose metabolism of fingerling blunt snout bream Megalobrama amblycephala. Aquacult. Nutr. 2013, 19, 701–708. [Google Scholar] [CrossRef]
- Li, X.F.; Tian, H.Y.; Zhang, D.D.; Jiang, G.Z.; Liu, W.B. Feeding frequency affects stress, innate immunity and disease resistance of juvenile blunt snout bream Megalobrama amblycephala. Fish Shellfish Immun. 2014, 38, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Wei, X.; Cheng, G. Leptin activates STAT3 and ERK 1/2 pathways and induces human breast cancer cell line MCF-7 proliferation. Chin. J. Exp. Surg. 2007, 24, 1551–1553. [Google Scholar]
- Zhang, D.; Lu, K.; Dong, Z.; Jiang, G.; Xu, W.; Liu, W. The effect of exposure to a high-fat diet on microRNA expression in the liver of blunt snout bream (Megalobrama amblycephala). PLoS ONE 2014, 9, e96132. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Agriculture. China Fishery Statistical Yearbook; Chinese Agriculture Press: Beijing, China, 2016. [Google Scholar]
- Zhou, C.; Liu, B.; Xie, J.; Ge, X. Effect of High Dietary Carbohydrate on Growth, Serum Physiological Response, and Hepatic Heat Shock Protein 70 Expression of Wuchang Bream (Megalobrama amblycephala) at Two Temperatures. Turk. J. Fish Aquat. Sci. 2013, 13, 211–220. [Google Scholar] [CrossRef]
- Zhou, C.; Liu, B.; Ge, X.; Xie, J.; Xu, P. Effect of dietary carbohydrate on the growth performance, immune response, hepatic antioxidant abilities and heat shock protein 70 expression of Wuchang bream, Megalobrama amblycephala. J. Appl. Ichthyol. 2013, 29, 1348–1356. [Google Scholar] [CrossRef]
- Zhou, C.; Ge, X.; Liu, B.; Xie, J.; Xu, P.; Zhou, Q.; Pan, L.; Chen, R. Effect of Dietary Carbohydrate Level on Growth Performance, Blood Chemistry, Hepatic Enzyme Activity, and Growth Hormone Gene Expression in Wuchang Bream (Megalobrama amblycephala). Isr. J. Aquac. Bamidgeh 2013, 65, 1–8. [Google Scholar] [CrossRef]
- Bensinger, S.J.; Tontonoz, P. Integration of metabolism and inflammation by lipid-activated nuclear receptors. Nature 2008, 454, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Guerre-Millo, M.; Rouault, C.; Poulain, P.; André, J.; Poitout, V.; Peters, J.M.; Gonzalez, F.J.; Fruchart, J.C.; Reach, G.; Staels, B. PPAR-α–Null Mice Are Protected From High-Fat Diet–Induced Insulin Resistance. Diabetes 2001, 50, 2809–2814. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, Y.; Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Nakayama, R.; Orikawa, Y.; Yoshioka, Y.; Tanaka, K.; Ueki, K.; Kadowaki, T. A Novel Peroxisome Proliferator-activated Receptor (PPAR)α Agonist and PPARγ Antagonist, Z-551, Ameliorates High-fat Diet-induced Obesity and Metabolic Disorders in Mice. J. Biol. Chem. 2015, 290, 14567–14581. [Google Scholar] [CrossRef] [PubMed]
- Zingg, J.M.; Hasan, S.T.; Nakagawa, K.; Canepa, E.; Ricciarelli, R.; Villacorta, L.; Azzi, A.; Meydani, M. Modulation of cAMP levels by high-fat diet and curcumin and regulatory effects on CD36/FAT scavenger receptor/fatty acids transporter gene expression. Biofactors 2016, 43, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Cortez, M.; Carmo, L.S.; Rogero, M.M.; Borelli, P.; Fock, R.A. A High-Fat Diet Increases IL-1, IL-6, and TNF-α Production by Increasing NF-κB and Attenuating PPAR-γ Expression in Bone Marrow Mesenchymal Stem Cells. Inflammation 2013, 36, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Gao, Z.X.; Zhao, H.; Zeng, C.; Luo, W.; Chen, B.; Wang, W.M. Identification and characterization of microRNAs involved in growth of blunt snout bream (Megalobrama amblycephala) by Solexa sequencing. BMC Genom. 2013, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bushati, N.; Cohen, S.M. MicroRNA functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 175. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Chan, W.C.; Hu, L.Y.; Lai, C.H.; Hsu, C.N.; Lin, W.C. Identification of homologous microRNAs in 56 animal genomes. Genomics 2010, 96, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhou, Q.; Li, S.; Luo, Q.; Cai, Y.; Lin, W.; Chen, H.; Yang, Y.; Hu, S.; Yu, J. The Silkworm (Bombyx mori) microRNAs and their expressions in multiple developmental stages. PLoS ONE 2008, 3, e2997. [Google Scholar] [CrossRef] [PubMed]
- Hafner, H.; Landgraf, P.; Ludwig, J.; Rice, A.; Ojo, T.; Lin, C.; Tuschl, T. Identification of microRNAs and other small regulat ory RNAs using cDNA library sequencing. Methods 2008, 44, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.J.; Rayner, K.J.; Suárez, Y.; Fernández-Hernando, C. The role of microRNAs in cholesterol efflux and hepatic lipid metabolism. Annu. Rev. Nutr. 2010, 31, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alonso, P.; Giardina, S.; Salas-Salvadó, J.; Arcelin, P.; Bulló, M. Chronic pistachio intake modulates circulating microRNAs related to glucose metabolism and insulin resistance in prediabetic subjects. Eur. J. Nutr. 2016, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.D.; Krüger, M.; Willmes, D.M.; Redemann, N.; Wunderlich, F.T.; Brönneke, H.S.; Merkwirth, C.; Kashkar, H.; Olkkonen, V.M.; Böttger, T. Obesity-induced overexpression of miRNA-143 inhibits insulin-stimulated Akt activation and impairs glucose metabolism. Nat. Cell Biol. 2011, 13, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Takanabe, R.; Ono, K.; Abe, Y.; Takaya, T.; Horie, T.; Wada, H.; Kita, T.; Satoh, N.; Shimatsu, A.; Hasegawa, K. Up-regulated expression of microRNA-143 in association with obesity in adipose tissue of mice fed high-fat diet. Biochem. Biophys. Res. Commun. 2008, 376, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Yuhong, J.; Leilei, T.; Fuyun, Z.; Hongyang, J.; Xiaowen, L.; Liying, Y.; Lei, Z.; Jingrong, M.; Jinpeng, Y. Identification and characterization of immune-related microRNAs in blunt snout bream, Megalobrama amblycephala. Fish Shellfish Immun. 2016, 49, 470–492. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhao, Y.; Wen, L.; Liu, Z.; Yan, F.; Gao, C. Identification and characterization of microRNAs in the spleen of common carp immune organ. J. Cell Biochem. 2014, 115, 1768–1778. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Guo, J.T.; Zhao, L.H.; Zhao, J.L. microRNA expression signature in skeletal muscle of Nile tilapia. Aquaculture 2012. [Google Scholar] [CrossRef]
- Fu, Y.; Shi, Z.; Wu, M.; Zhang, J.; Jia, L.; Chen, X. Identification and Differential Expression of microRNAs during Metamorphosis of the Japanese Flounder (Paralichthys olivaceus). PLoS ONE 2011, 6, e22957. [Google Scholar] [CrossRef] [PubMed]
- Goedeke, L.; Fernándezhernando, C. microRNAs: A connection between cholesterol metabolism and neurodegeneration. Neurobiol. Dis. 2014, 72, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Vosgha, H.; Salajegheh, A.; Smith, R.A.; Lam, A.K. The Important Roles of miR-205 in Normal Physiology, Cancers and as a Potential Therapeutic Target. Curr. Cancer Drug Target. 2014, 14, 621–637. [Google Scholar] [CrossRef]
- Lai, K.P.; Li, J.W.; Tse, A.C.; Chan, T.F.; Wu, R.S. Hypoxia alters steroidogenesis in female marine medaka through miRNAs regulation. Aquat. Toxicol. 2015, 172, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Betel, D.; Koppal, A.; Agius, P.; Sander, C.; Leslie, C. Comprehensive modeling of microRNA targets predicts functional non-conserved and non-canonical sites. Genome Biol. 2010, 11, r90. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Rehmsmeier, M.; Steffen, P.; Hochsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef] [PubMed]
- Mir-Coll, J.; Duran, J.; Slebe, F.; García-Rocha, M.; Gomis, R.; Gasa, R.; Guinovart, J.J. Genetic models rule out a major role of β cell glycogen in the control of glucose homeostasis. Diabetologia 2016, 59, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xirouchaki, C.E.; Mangiafico, S.P.; Bate, K.; Zheng, R.; Huang, A.M.; Bing, W.T.; Lamont, B.; Pong, W.; Favaloro, J.; Blair, A.R. Impaired glucose metabolism and exercise capacity with muscle-specific glycogen synthase 1 (gys1) deletion in adult mice. Mol. Metab. 2016, 5, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Beale, E.G.; Hammer, R.E.; Antoine, B.; Forest, C. Disregulated glyceroneogenesis: PCK1 as a candidate diabetes and obesity gene. Trends Endocrin. Metab. 2004, 15, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Herrera, I.; Martín, M.Á.; Goya, L.; Ramos, S. Cocoa flavonoids attenuate high glucose-induced insulin signalling blockade and modulate glucose uptake and production in human HepG2 cells. Food Chem. Toxicol. 2013, 64, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Stamateris, R.E.; Sharma, R.B.; Kong, Y.; Ebrahimpour, P.; Panday, D.; Ranganath, P.; Zou, B.; Levitt, H.; Parambil, N.A.; O'Donnell, C.P. Glucose induces mouse β cell proliferation via IRS2, mTOR and cyclin D2 but not the insulin receptor. Diabetes 2016, 65, 981–995. [Google Scholar] [CrossRef] [PubMed]
- Sylow, L.; Kleinert, M.; Pehmøller, C.; Prats, C.; Chiu, T.T.; Klip, A.; Richter, E.A.; Jensen, T.E. Akt and Rac1 signaling are jointly required for insulin-stimulated glucose uptake in skeletal muscle and downregulated in insulin resistance. Cell Signal. 2014, 26, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Bae, E.; Kim, H.E.; Koh, E.; Kim, K.S. Phosphoglucomutase1 is necessary for sustained cell growth under repetitive glucose depletion. Febs. Lett. 2014, 588, 3074–3080. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Fang, R.; Li, Y.; Li, L.; Zhang, W.; Wang, H.; Chen, F.; Zhang, S.; Zhang, X.; Ye, L. The oncoprotein HBXIP suppresses gluconeogenesis through modulating PCK1 to enhance the growth of hepatoma cells. Cancer Lett. 2016, 382, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Chen, S.; Yu, H.; Lu, X.; Chen, J.; Wang, L.; Huang, J.; Fan, Z.; Gu, D. Associations of PLA2G7 gene polymorphisms with plasma lipoprotein-associated phospholipase A2 activity and coronary heart disease in a Chinese Han population: The Beijing atherosclerosis study. Hum. Genet. 2009, 125, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Beth, S.S.; David, R.C.; Svati, H.S.; Sarah, C.N.; Anthony, B.; Hale, A.B.; Carol, H.; Pascal, J.G.; Jeffery, M.V.; David, S.; William, E.K.; Simon, G.G.; Elizabeth, R.H. Comprehensive genetic analysis of the platelet activating factor acetylhydrolase (PLA2G7) gene and cardiovascular disease in case–control and family datasets. Hum. Mol. Genet. 2008, 17, 1318–1328. [Google Scholar] [CrossRef]
- Leuker, C.E.; Sonneborn, A.; Delbrück, S.; Ernst, J.F. Sequence and promoter regulation of the PCK1 gene encoding phosphoenolpyruvate carboxykinase of the fungal pathogen Candida albicans. Gene 1997, 192, 235–240. [Google Scholar] [CrossRef]
- Ramsey, K.M.; Mills, K.F.; Satoh, A.; Imai, S. Age-associated loss of Sirt1-mediated enhancement of glucose-stimulated insulin secretion in β cell-specific Sirt1-overexpressing (BESTO) mice. Aging Cell 2008, 7, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Milne, J.C.; Lambert, P.D.; Schenk, S.; Carney, D.P.; Smith, J.J.; Gagne, D.J.; Jin, L.; Boss, O.; Perni, R.B.; Vu, C.B. Small molecule activators of SIRT1 as therapeutics for the treatment of type 2 diabetes. Nature 2007, 450, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Pfluger, P.T.; Herranz, D.; Velasco-Miguel, S.; Serrano, M.; Tschöp, M.H. Sirt1 protects against high-fat diet-induced metabolic damage. Proc. Natl. Acad. Sci. USA 2008, 105, 9793–9798. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, J.; Zhang, L.; Xiong, Y.; Chen, S.; Xing, H.; Tian, Z.; Tang, K.; Wei, H.; Rao, Q. RPS27a promotes proliferation, regulates cell cycle progression and inhibits apoptosis of leukemia cells. Biochem. Biophys. Res. Commun. 2014, 446, 1204–1210. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.X.; DeVine, T.; Challagundla, K.B.; Dai, M.S. Interplay between Ribosomal Protein S27a and MDM2 Protein in p53 Activation in Response to Ribosomal Stress. J. Biol. Chem. 2011, 286, 22730–22741. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Geng, Y.; Luo, J.; Shen, W.; Zhu, W.; Meng, C.; Li, M.; Zhou, X.; Zhang, S.; Cao, J. Downregulation of ubiquitin inhibits the proliferation and radioresistance of non-small cell lung cancer cells in vitro and in vivo. Sci. Rep. 2015, 5, 9476. [Google Scholar] [CrossRef] [PubMed]
- Ouk, T.; Gautier, S.; Pétrault, M.; Montaigne, D.; Maréchal, X.; Masse, I.; Devedjian, J.C.; Deplanque, D.; Bastide, M.; Nevière, R. Effects of the PPAR-α agonist fenofibrate on acute and short-term consequences of brain ischemia. J. Cereb. Blood Flow Metab. 2014, 34, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.S.; Tsai, H.D.; Cheung, W.M.; Hsu, C.Y.; Lin, T.N. PPAR-γ Ameliorates Neuronal Apoptosis and Ischemic Brain Injury via Suppressing NF-κB-Driven p22phox Transcription. Mol. Neurobiol. 2016, 53, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Sarruf, D.A.; Talukdar, S.; Sharma, S.; Li, P.; Bandyopadhyay, G.; Nalbandian, S.; Fan, W.; Gayen, J.R.; Mahata, S.K. Brain PPARγ Promotes Obesity and is Required for the Insulin–Sensitizing Effect of Thiazolidinediones. Nat. Med. 2011, 17, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Rome, S.; Meugnier, E.; Vidal, H. The ubiquitin-proteasome pathway is a new partner for the control of insulin signaling. Curr. Opin. Clin. Nutr. 2004, 7, 249–254. [Google Scholar] [CrossRef]
- Genini, D.; Carbone, G.M.; Catapano, C.V. Multiple Interactions between Peroxisome Proliferators-Activated Receptors and the Ubiquitin-Proteasome System and Implications for Cancer Pathogenesis. PPAR Res. 2008. [Google Scholar] [CrossRef] [PubMed]
- Marfella, R.; D’Amico, M.; Esposito, K.; Baldi, A.; di Filippo, C.; Siniscalchi, M.; Sasso, F.C.; Portoghese, M.; Cirillo, F.; Cacciapuoti, F. The Ubiquitin-Proteasome System and Inflammatory Activity in Diabetic Atherosclerotic Plaques. Diabetes 2006, 55, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Nguyen, M.A.; Yoshizaki, T.; Favelyukis, S.; Patsouris, D.; Imamura, T.; Verma, I.M.; Olefsky, J.M. Suppression of PPAR-γ attenuates insulin-stimulated glucose uptake by affecting both GLUT1 and GLUT4 in 3T3-L1 adipocytes. Am. J. Physiol. 2007, 293, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, Y.; Zhang, Y.; Ning, Z.; Li, Y.; Zhao, Q.; Lu, H.; Huang, R.; Xia, X.; Feng, Q. The draft genome of the grass carp (Ctenopharyngodon idellus) provides insights into its evolution and vegetarian adaptation. Nat. Genet. 2015, 47, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Cui, X.; Zhang, Y.; Yang, C.; Jiang, Y. Identification of miRNAs associated with sexual maturity in chicken ovary by Illumina small RNA deep sequencing. BMC Genom. 2013, 14, 352. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, A.J. Java Treeview-extensible visualization of microarray data. Bioinformatics 2004, 20, 3246–3248. [Google Scholar] [CrossRef] [PubMed]
- Hulsegge, I.; Kommadath, A.; Smits, M.A. Globaltest and GOEAST: Two different approaches for Gene Ontology analysis. BMC Proc. 2009, 3, S10. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Mao, X.; Cai, T.; Luo, J.; Wei, L. KOBAS server: A web-based platform for automated annotation and pathway identification. Nucleic Acids Res. 2006, 34, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Kuhn, M.; Simonovic, M.; Roth, A.; Minguez, P.; Doerks, T.; Stark, M.; Muller, J.; Bork, P. The STRING database in 2011: functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, L.-H.; Lin, Y.; Pan, W.-J.; Huang, X.; Ge, X.-P.; Ren, M.-C.; Zhou, Q.-L.; Liu, B. Identification of Differentially Expressed Micrornas Associate with Glucose Metabolism in Different Organs of Blunt Snout Bream (Megalobrama amblycephala). Int. J. Mol. Sci. 2017, 18, 1161. https://doi.org/10.3390/ijms18061161
Miao L-H, Lin Y, Pan W-J, Huang X, Ge X-P, Ren M-C, Zhou Q-L, Liu B. Identification of Differentially Expressed Micrornas Associate with Glucose Metabolism in Different Organs of Blunt Snout Bream (Megalobrama amblycephala). International Journal of Molecular Sciences. 2017; 18(6):1161. https://doi.org/10.3390/ijms18061161
Chicago/Turabian StyleMiao, Ling-Hong, Yan Lin, Wen-Jing Pan, Xin Huang, Xian-Ping Ge, Ming-Chun Ren, Qun-Lan Zhou, and Bo Liu. 2017. "Identification of Differentially Expressed Micrornas Associate with Glucose Metabolism in Different Organs of Blunt Snout Bream (Megalobrama amblycephala)" International Journal of Molecular Sciences 18, no. 6: 1161. https://doi.org/10.3390/ijms18061161
APA StyleMiao, L. -H., Lin, Y., Pan, W. -J., Huang, X., Ge, X. -P., Ren, M. -C., Zhou, Q. -L., & Liu, B. (2017). Identification of Differentially Expressed Micrornas Associate with Glucose Metabolism in Different Organs of Blunt Snout Bream (Megalobrama amblycephala). International Journal of Molecular Sciences, 18(6), 1161. https://doi.org/10.3390/ijms18061161