The AGPase Family Proteins in Banana: Genome-Wide Identification, Phylogeny, and Expression Analyses Reveal Their Involvement in the Development, Ripening, and Abiotic/Biotic Stress Responses

 ,
,
Abstract
:
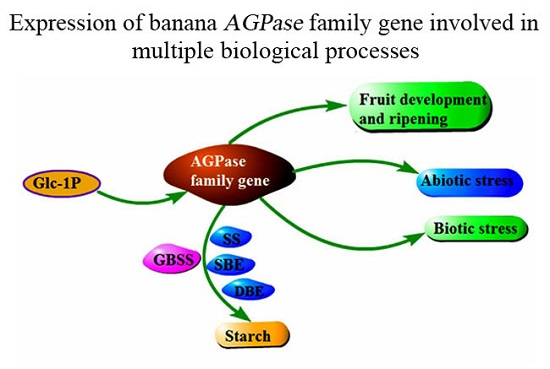
1. Introduction
2. Results
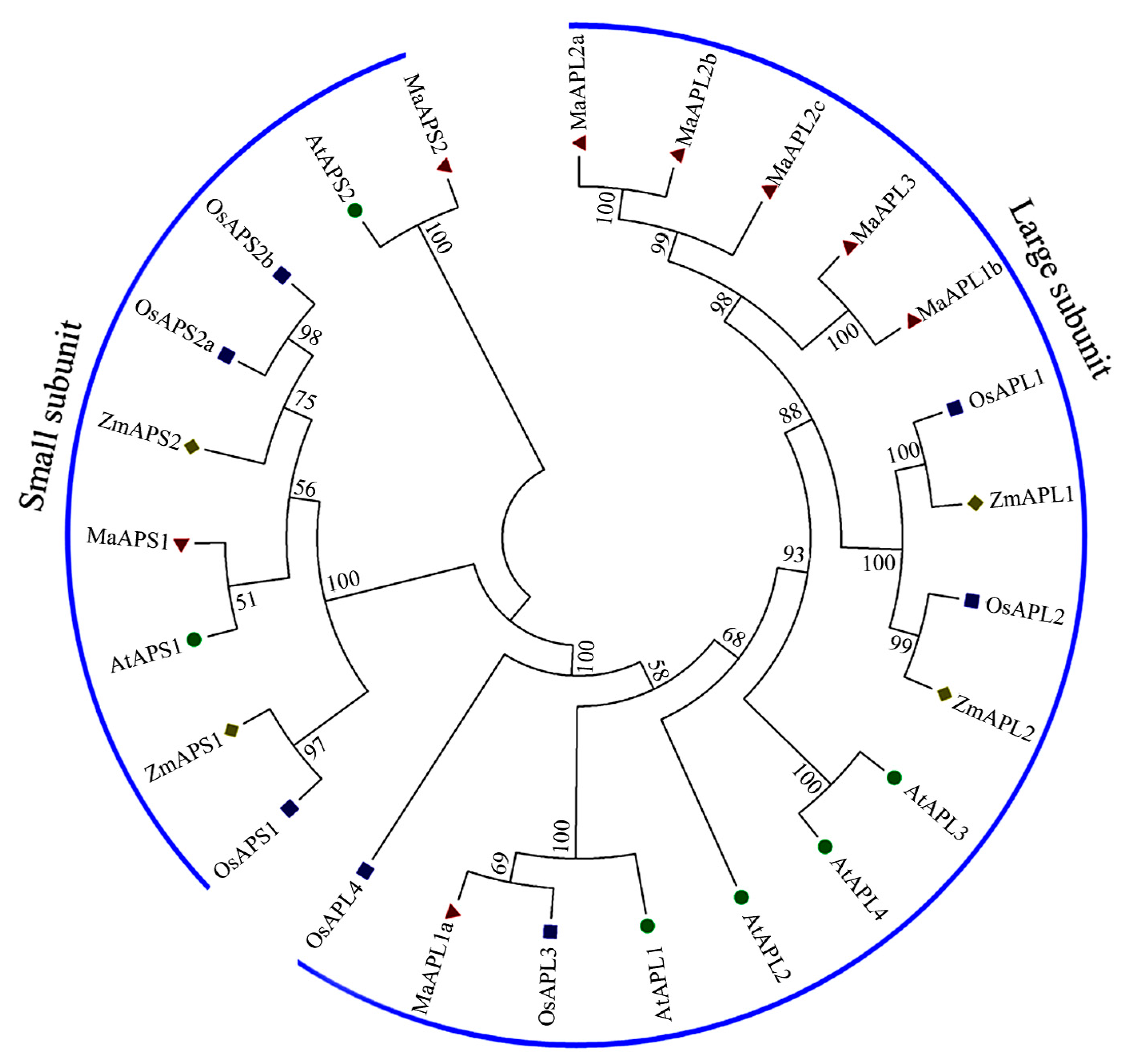
2.1. Identification and Phylogenetic Analysis of Banana MaAGPase Genes
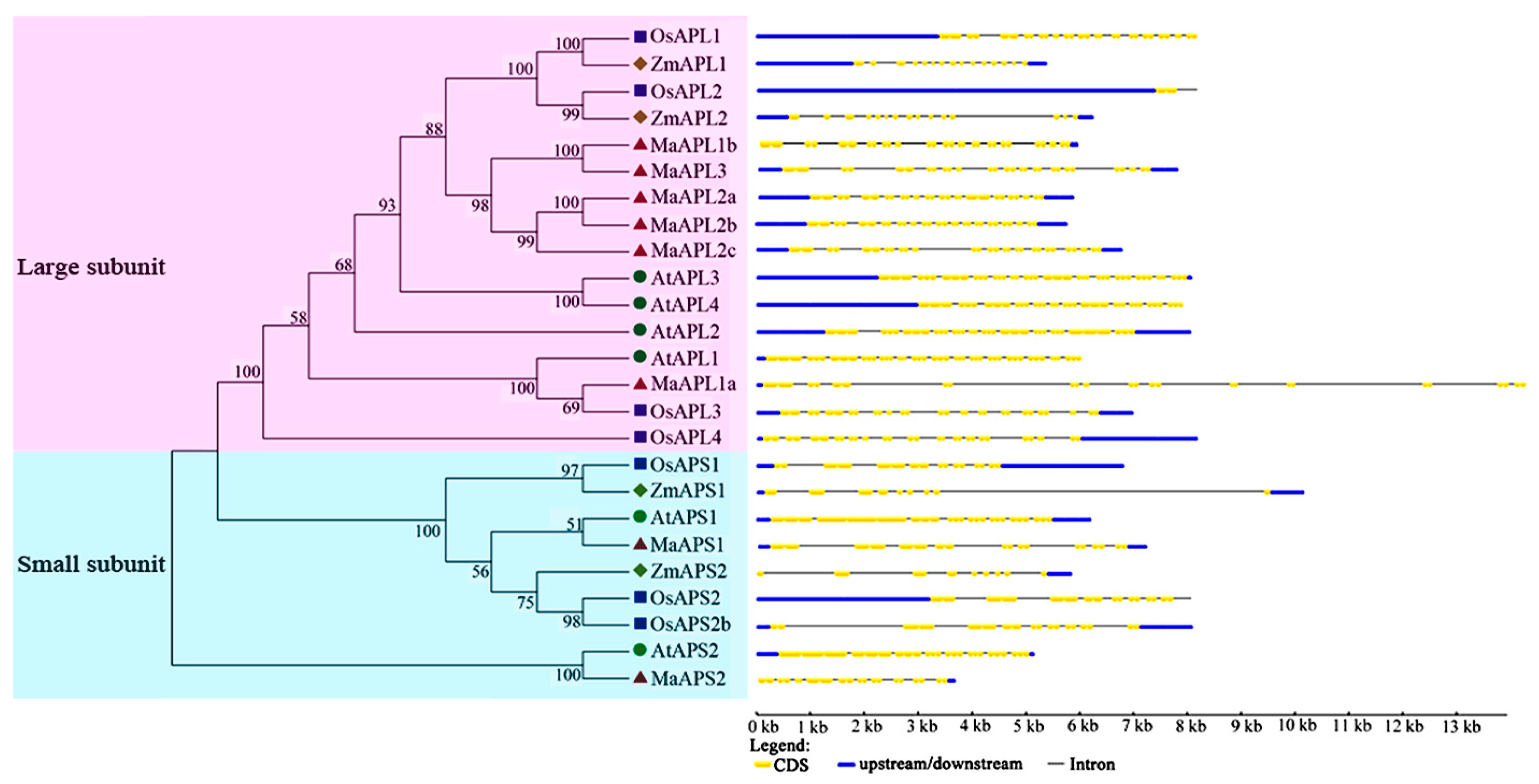
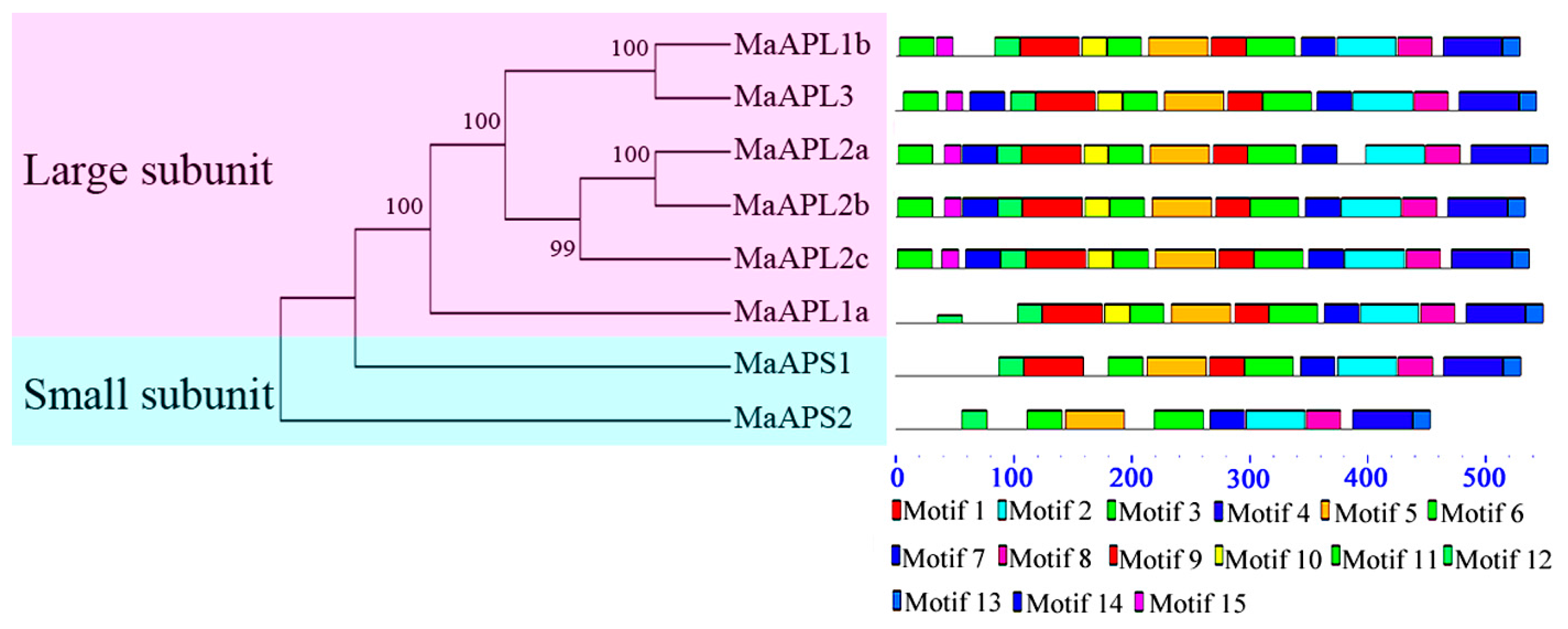
2.2. Gene Structure and Conserved Motif Analysis of Banana MaAGPase Genes
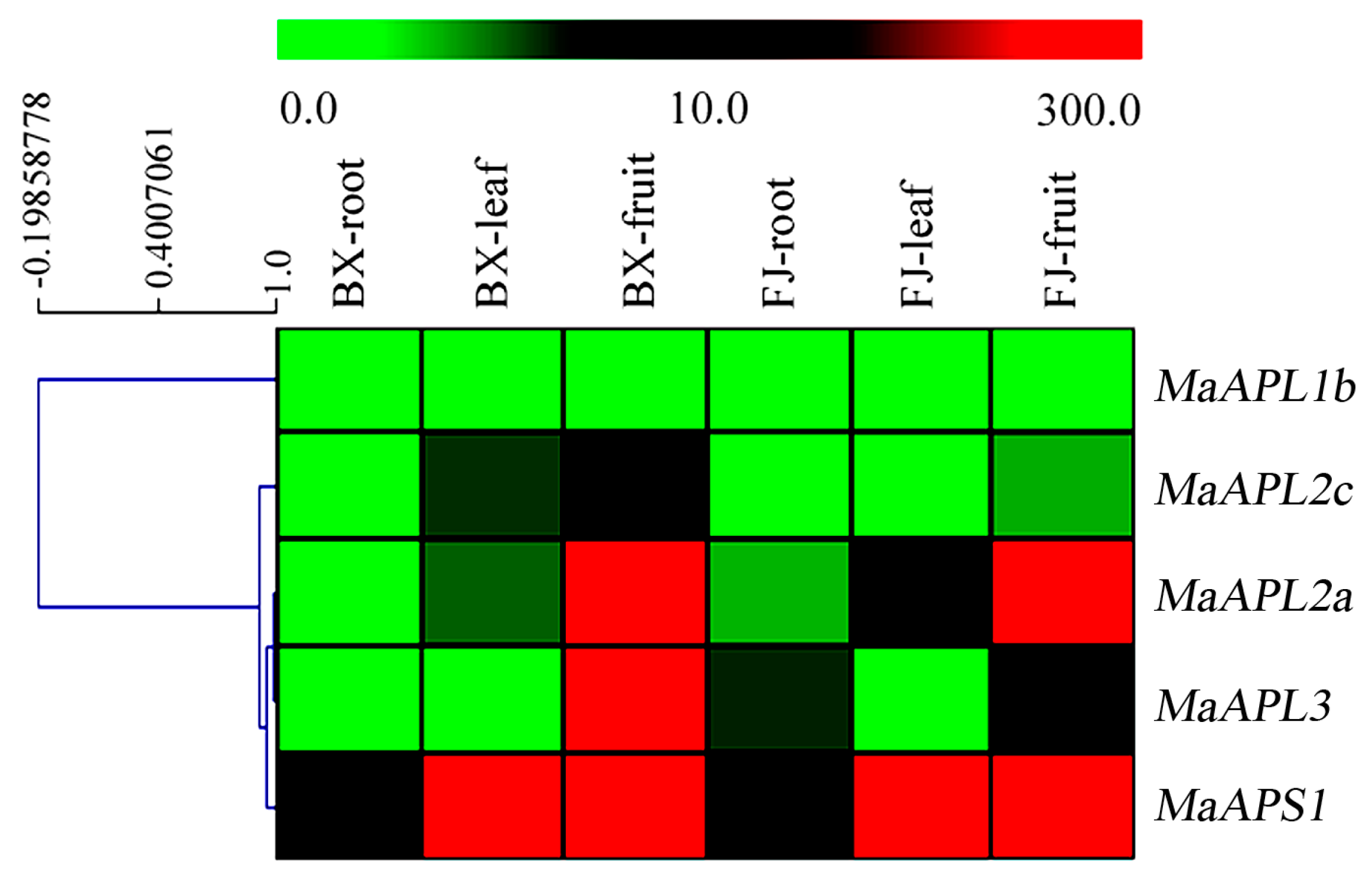
2.3. Spatial Expression Analysis of MaAGPase Genes in Banana
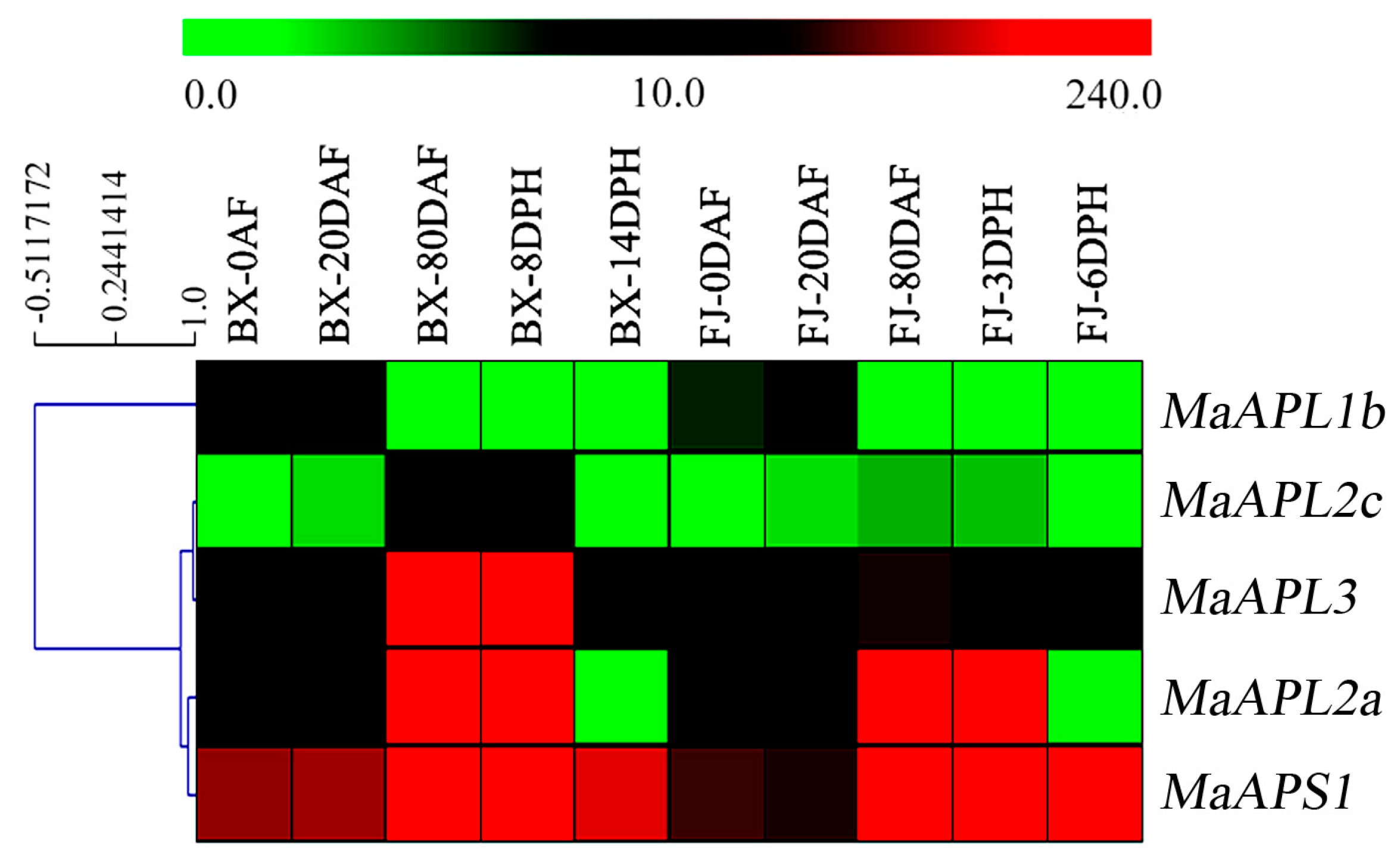
2.4. Temporal Expression Analysis of MaAGPase Genes during Banana Fruit Development and Ripening
2.5. Expression of MaAGPases under Cold, Salt, and Osmotic Stresses
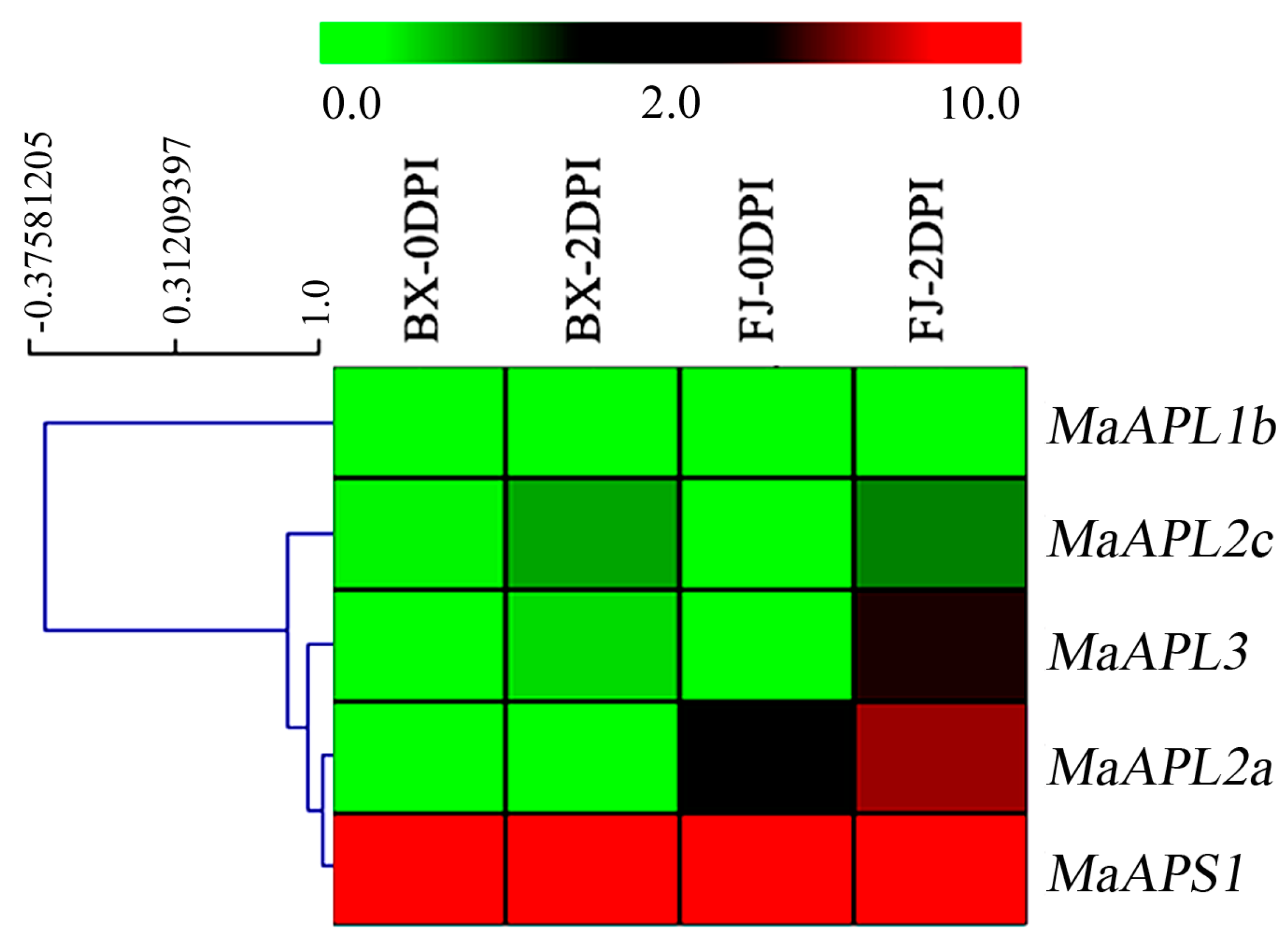
2.6. Expression Profiles of Banana MaAGPases in Response to Fusarium Oxysporum f.sp. Cubense (Foc) Tropical Race 4 (TR4) Infection
2.7. Validation of the Differentially Expressed MaAGPase Genes by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
2.8. In Silico Identification of Hormone-Related and Stress-Related Cis-Acting Elements in the Promoters of MaAGPase Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Identification and Phylogenetic Analyses of MaAGPase Proteins in Banana
4.3. Characterization Analysis of Protein Properties and Gene Structure
4.4. Transcriptomic Analysis
4.5. QRT-PCR Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, Y.; Li, C.; Gu, Z.; Hong, Y.; Cheng, L.; Li, Z. Effect of modification with 1,4-α-glucan branching enzyme on the rheological properties of cassava starch. Int. J. Biol. Macromol. 2017, 103, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Tiessen, A.; Hendriks, J.H.; Stitt, M.; Branscheid, A.; Gibon, Y.; Farré, E.M.; Geigenberger, P. Starch synthesis in potato tubers is regulated by post-translational redox modification of ADP-glucose pyrophosphorylase: A novel regulatory mechanism linking starch synthesis to the sucrose supply. Plant Cell 2002, 14, 2191–2213. [Google Scholar] [CrossRef] [PubMed]
- Toyosawa, Y.; Kawagoe, Y.; Matsushima, R.; Crofts, N.; Ogawa, M.; Fukuda, M.; Kumamaru, T.; Okazaki, Y.; Kusano, M.; Saito, K.; et al. Deficiency of starch synthase IIIa and IVb alters starch granule morphology from polyhedral to spherical in rice endosperm. Plant Physiol. 2016, 170, 1255–1270. [Google Scholar] [CrossRef] [PubMed]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [PubMed]
- Jourda, C.; Cardi, C.; Gibert, O.; Giraldo Toro, A.; Ricci, J.; Mbéguié-A-Mbéguié, D.; Yahiaoui, N. Lineage-specific evolutionary histories and regulation of major starch metabolism genes during banana ripening. Front. Plant Sci. 2016, 7, 1778. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.X.; Sun, P.G.; Liu, W.X.; Xu, B.Y.; Jin, Z.Q. Identification of genes encoding granule-bound starch synthase involved in amylose metabolism in banana fruit. PLoS ONE 2014, 9, e88077. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.X.; Sun, P.G.; Liu, Q.; Jia, C.H.; Liu, J.H.; Hu, W.; Jin, Z.Q.; Xu, B.Y. Soluble starch synthase III-1 in amylopectin metabolism of banana fruit: Characterization, expression, enzyme activity, and functional analyses. Front. Plant Sci. 2017, 8, 454. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.X.; Sun, P.G.; Liu, Q.; Miao, Y.L.; Liu, J.H.; Zhang, K.X.; Hu, W.; Zhang, J.B.; Wang, J.Y.; Wang, Z.; et al. Genome-wide analyses of SWEET family proteins reveal involvement in fruit development and abiotic/biotic stress responses in banana. Sci. Rep. 2017, 7, 3536. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Denyer, K.; Martin, C. The synthesis of the starch granule. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Batra, R.; Saripalli, G.; Mohan, A.; Gupta, S.; Gill, K.S.; Varadwaj, P.K.; Balyan, H.S.; Gupta, P.K. Comparative analysis of AGPase genes and encoded proteins in eight monocots and three dicots with emphasis on wheat. Front. Plant Sci. 2017, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Smith, A.M. Starch biosynthesis. Plant Cell 1995, 7, 971–985. [Google Scholar] [CrossRef] [PubMed]
- Saripalli, G.; Gupta, P.K. AGPase: Its role in crop productivity with emphasis on heat tolerance in cereals. Theor. Appl. Genet. 2015, 128, 1893–1916. [Google Scholar] [CrossRef] [PubMed]
- Hannah, L.C.; Shaw, J.R.; Giroux, M.J.; Reyss, A.; Prioul, J.L.; Bae, J.M.; Lee, J.Y. Maize genes encoding the small subunit of ADP-glucose pyrophosphorylase. Plant Physiol. 2001, 127, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Tiessen, A.; Nerlich, A.; Faix, B.; Hümmer, C.; Fox, S.; Trafford, K.; Weber, H.; Weschke, W.; Geigenberger, P. Subcelluar analysis of starch metabolism in developing barley seeds using a non-aqueous fractionation method. J. Exp. Bot. 2012, 63, 2071–2087. [Google Scholar] [CrossRef] [PubMed]
- Crevillén, P.; Ventriglia, T.; Pinto, F.; Orea, A.; Mérida, Á.; Romero, J.M. Differential pattern of expression and sugar regulation of Arabidopsis thaliana ADP-glucose pyrophosphorylase-encoding genes. J. Biol. Chem. 2005, 280, 8143–8149. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.H.; Park, Y.J. Sequence variations in OsAGPase significantly associated with amylose content and viscosity properties in rice (Oryza sativa L.). Genet. Res. 2012, 94, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.J.; Peng, C.; Zhang, J.; Cai, Y.; You, X.M.; Kong, F.; Yan, H.G.; Wang, G.X.; Wang, L.; Jin, J.; et al. ADP-glucose pyrophosphorylase large subunit 2 is essential for storage substance accumulation and subunit interactions in rice endosperm. Plant Sci. 2016, 249, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Hennen-Bierwagen, T.A.; Myers, A.M. Functions of multiple genes encoding ADP-glucose pyrophosphorylase subunits in maize endosperm, embryo, and leaf. Plant Physiol. 2014, 164, 596–611. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.X.; Chen, Y.X.; Tao, X.; Cheng, X.J.; Wang, H.Y. Isolation and characterization of cDNAs and genomic DNAs encoding ADP-glucose pyrophosphorylase large and small subunits from sweet potato. Mol. Genet. Genom. 2016, 291, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Au, S.L.; Tan, S.H.; Harikrishna, K.; Napis, S. Isolation and characterization of cDNA clones encoding ADP-glucose pyrophosphorylase from sago palm (Metroxylon sagu). J. Biochem. Mol. Biol. Biophys. 2002, 6, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Nardozza, S.; Boldingh, H.L.; Osorio, S.; Höhne, M.; Wohlers, M.; Gleave, A.P.; MacRae, E.A.; Richardson, A.C.; Atkinson, R.G.; Sulpice, R.; et al. Metabolic analysis of kiwifruit (Actinidia deliciosa) berries from extreme genotypes reveals hallmarks for fruit starch metabolism. J. Exp. Bot. 2013, 64, 5049–5063. [Google Scholar] [CrossRef] [PubMed]
- Petreikov, M.; Shen, S.; Yeselson, Y.; Levin, I.; Bar, M.; Schaffer, A.A. Temporally extended gene expression of the ADP-Glc pyrophosphorylase large subunit (AgpL1) leads to increased enzyme activity in developing tomato fruit. Planta 2006, 224, 1465–1479. [Google Scholar] [CrossRef] [PubMed]
- Mugford, S.T.; Fernandez, O.; Brinton, J.; Flis, A.; Krohn, N.; Encke, B.; Feil, R.; Sulpice, R.; Lunn, J.E.; Stitt, M.; et al. Regulatory properties of ADP glucose pyrophosphorylase are required for adjustment of leaf starch synthesis in different photoperiods. Plant Physiol. 2014, 166, 1733–1747. [Google Scholar] [CrossRef] [PubMed]
- Sulmon, C.; Gouesbet, G.; Ramel, F.; Cabello-Hurtado, F.; Penno, C.; Bechtold, N.; Couée, I.; El Amrani, A. Carbon dynamics, development and stress responses in Arabidopsis: Involvement of the APL4 subunit of ADP-glucose pyrophosphorylase (starch synthesis). PLoS ONE 2011, 6, e26855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, M.S.; Min, S.R.; Lee, S.M.; Kim, K.N.; Liu, J.R.; Paek, K.H.; Shin, J.S.; Bae, J.M. A sepal-expressed ADP-glucose pyrophosphorylase gene (NtAGP) is required for petal expansion growth in 'Xanthi' tobacco. Plant Physiol. 2007, 145, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Taliercio, E. Characterization of an ADP-glucose pyrophosphorylase small subunit gene expressed in developing cotton (Gossypium hirsutum) fibers. Mol. Biol. Rep. 2011, 38, 2967–2973. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Eom, J.S.; Hwang, S.K.; Shin, D.; An, G.; Okita, T.W.; Jeon, J.S. Plastidic phosphoglucomutase and ADP-glucose pyrophosphorylase mutants impair starch synthesis in rice pollen grains and cause male sterility. J. Exp. Bot. 2016, 67, 5557–5569. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.G.; Kobayashi, Y.; Sanuki, A.; Kondo, S.; Fukuda, N.; Ezura, H.; Sugaya, S.; Matsukura, C. Salinity induces carbohydrate accumulation and sugar-regulated starch biosynthetic genes in tomato (Solanum lycopersicum L. cv. ‘Micro-Tom’) fruits in an ABA- and osmotic stress-independent manner. J. Exp. Bot. 2010, 61, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Gamm, M.; Héloir, M.C.; Bligny, R.; Vaillant-Gaveau, N.; Trouvelot, S.; Alcaraz, G.; Frettinger, P.; Clément, C.; Pugin, A.; Wendehenne, D.; et al. Changes in carbohydrate metabolism in Plasmopara viticola-infected grapevine leaves. Mol. Plant Microbe Interact. 2011, 24, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Wiberley-Bradford, A.E.; Busse, J.S.; Jiang, J.; Bethke, P.C. Sugar metabolism, chip color, invertase activity, and gene expression during long-term cold storage of potato (Solanum tuberosum) tubers from wild-type and vacuolar invertase silencing lines of Katahdin. BMC Res. Notes 2014, 7, 801. [Google Scholar] [CrossRef] [PubMed]
- Majoul-Haddad, T.; Bancel, E.; Martre, P.; Triboi, E.; Branlard, G. Effect of short heat shocks applied during grain development on wheat (Triticum aestivum L.) grain proteome. J. Cereal Sci. 2013, 57, 486–495. [Google Scholar] [CrossRef]
- Thitisaksakul, M.; Jimenez, R.C.; Arias, M.C.; Beckles, D.M. Effects of environmental factors on cereal starch biosynthesis and composition. J. Cereal Sci. 2012, 56, 67–80. [Google Scholar] [CrossRef]
- Gibert, O.; Dufour, D.; Giraldo, A.; Sánchez, T.; Reynes, M.; Pain, J.P.; González, A.; Fernández, A.; Díaz, A. Differentiation between cooking bananas and dessert bananas. 1. Morphological and compositional characterization of cultivated Colombian Musaceae (Musa sp.) in relation to consumer preferences. J. Agric. Food Chem. 2009, 57, 7857–7869. [Google Scholar] [CrossRef] [PubMed]
- D’Hont, A.; Denoeud, F.; Aury, J.M.; Baurens, F.C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Wang, L.; Tie, W.; Yan, Y.; Ding, Z.; Liu, J.; Li, M.; Peng, M.; Xu, B.; Jin, Z. Genome-wide analyses of the bZIP family reveal their involvement in the development, ripening and abiotic stress response in banana. Sci. Rep. 2016, 6, 30203. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.H.; Lakhwani, D.; Pathak, S.; Bhambhani, S.; Baq, S.K.; Trivedi, P.K. Genome-wide identification and expression analysis of the mitogen-activated protein kinase gene family from banana suggest involvement of specific members in different stages of fruit ripening. Funct. Integr. Genom. 2014, 14, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Backiyarani, S.; Uma, S.; Nithya, S.; Chandrasekar, A.; Saraswathi, M.S.; Thanqavelu, R.; Mayilvaqanan, M.; Sundararaju, P.; Sinqh, N.K. Genome-wide analysis and differential expression of chitinases in banana against root lesion nematode (Pratylenchus coffeae) and eumusa leaf spot (Mycosphaerella eumusae) pathogens. Appl. Biochem. Biotechnol. 2015, 175, 3585–3598. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.K.; Huang, X.Q.; Brûlé-Babel, A. Molecular characterization and sequence diversity of genes encoding the large subunit of the ADP-glucose pyrophosphorylase in wheat (Triticum aestivum L.). J. Appl. Genet. 2016, 57, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Noh, S.A.; Kwak, M.S.; Lee, H.S.; Huh, G.H.; Liu, J.R.; Shin, J.S.; Bae, J.M. Genomic organizations of two small subunit ADP-glucose pyrophosphorylase genes from sweet potato. Gene 2004, 339, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Thorbjørnsen, T.; Villand, P.; Kleczkowski, L.A.; Olsen, O.A. A single gene encodes two different transcripts for the ADP-glucose pyrophosphorylase small subunit from barley (Hordeum vulgare). Biochem. J. 1996, 313, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, B.; Wang, L.; Chen, X.; Wang, W.; Wang, Z.; Xiong, F. Systematic analysis of pericarp starch accumulation and degradation during wheat caryopsis development. PLoS ONE 2015, 10, e0138228. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.; Odipio, J.; Tripathi, J.N.; Tusiime, G. A rapid technique for screening banana cultivars for resistance to Xanthomonas wilt. Eur. J. Plant Pathol. 2008, 121, 9–19. [Google Scholar] [CrossRef]
- Vanhove, A.C.; Vermaelen, W.; Panis, B.; Swennen, R.; Carpentier, S.C. Screening the banana biodiversity for drought tolerance: Can an in vitro growth model and proteomics be used as a tool to discover tolerant varieties and understand homeostasis. Front. Plant Sci. 2012, 3, 176. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, J.; Jia, C.; Liu, J.; Li, Y.; Yin, X.; Xu, B.; Jin, Z. De novo characterization of the banana root transcriptome and analysis of gene expression under Fusarium oxysporum f. sp. Cubense tropical race 4 infection. BMC Genom. 2012, 13, 650. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Sagaram, U.S.; Burns, J.K.; Li, J.L.; Wang, N. Response of sweet orange (Citrus sinensis) to ‘Candidatus Liberibacter asiaticus’ infection: Microscopy and microarray analyses. Phytopathology 2009, 99, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Horst, R.J.; Engelsdorf, T.; Sonnewald, U.; Voll, L.M. Infection of maize leaves with Ustilago maydis prevents establishment of C4 photosynthesis. J. Plant Physiol. 2008, 165, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Thorneycroft, D.; Hosein, F.; Thangavelu, M.; Clark, J.; Vizir, L.; Burrell, M.M.; Ainsworth, C. Characterization of a gene from chromosome 1B encoding the large subunit of ADP glucose pyrophorylase from wheat: Evolutionary divergence and differential expression of Agp2 genes between leaves and developing endosperm. Plant Biotechnol. J. 2003, 1, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wu, Y.; Xu, M.; Gao, T.; Wang, P.; Wang, L.; Guo, T.; Kang, G. Virus-induced gene silencing identifies an important role of the TaRSR1 transcription factor in starch synthesis in bread wheat. Int. J. Mol. Sci. 2016, 17, 1557. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Kuang, S.; Zhang, A.D.; Zhang, W.S.; Chen, M.J.; Yin, X.R.; Chen, K.S. Characterization of starch degradation related genes in postharvest kiwifruit. Int. J. Mol. Sci. 2016, 17, 2112. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Miao, H.X.; Ma, Y.W.; Wang, L.; Hu, G.B.; Ye, Z.X.; Zhao, J.T.; Qin, Y.H. CrSKP1, an SKP1-like gene, is involved in the self-incompatibility reaction of “Wuzishatangju” (Citrus reticulata Blanco). Int. J. Mol. Sci. 2015, 16, 21695–21710. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | ABRE (ABA) | ARE (Anaerobic) | AuxRR(Auxin) | Box-W1 (Fungal) | CGTCA-Motif (MeJA) | Circadian | ERE (Ethylene) | GARE (Gibberellin) | HSE (Heat) | LTR (Cold) | MBS (Drought) | TCA-Element (Salicylic Acid) | TC-Rich Repeats (Defense) | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MaAPL1a | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 7 |
| MaAPL1b | 0 | 2 | 0 | 1 | 3 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 10 |
| MaAPL2a | 3 | 0 | 1 | 1 | 5 | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 3 | 16 |
| MaAPL2b | 3 | 0 | 1 | 1 | 4 | 2 | 0 | 0 | 0 | 0 | 1 | 2 | 3 | 17 |
| MaAPL2c | 4 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 2 | 0 | 1 | 1 | 12 |
| MaAPL3 | 4 | 0 | 1 | 0 | 7 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 15 |
| MaAPS1 | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 6 |
| MaAPS2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 2 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, H.; Sun, P.; Liu, Q.; Liu, J.; Xu, B.; Jin, Z. The AGPase Family Proteins in Banana: Genome-Wide Identification, Phylogeny, and Expression Analyses Reveal Their Involvement in the Development, Ripening, and Abiotic/Biotic Stress Responses. Int. J. Mol. Sci. 2017, 18, 1581. https://doi.org/10.3390/ijms18081581
Miao H, Sun P, Liu Q, Liu J, Xu B, Jin Z. The AGPase Family Proteins in Banana: Genome-Wide Identification, Phylogeny, and Expression Analyses Reveal Their Involvement in the Development, Ripening, and Abiotic/Biotic Stress Responses. International Journal of Molecular Sciences. 2017; 18(8):1581. https://doi.org/10.3390/ijms18081581
Chicago/Turabian StyleMiao, Hongxia, Peiguang Sun, Qing Liu, Juhua Liu, Biyu Xu, and Zhiqiang Jin. 2017. "The AGPase Family Proteins in Banana: Genome-Wide Identification, Phylogeny, and Expression Analyses Reveal Their Involvement in the Development, Ripening, and Abiotic/Biotic Stress Responses" International Journal of Molecular Sciences 18, no. 8: 1581. https://doi.org/10.3390/ijms18081581
APA StyleMiao, H., Sun, P., Liu, Q., Liu, J., Xu, B., & Jin, Z. (2017). The AGPase Family Proteins in Banana: Genome-Wide Identification, Phylogeny, and Expression Analyses Reveal Their Involvement in the Development, Ripening, and Abiotic/Biotic Stress Responses. International Journal of Molecular Sciences, 18(8), 1581. https://doi.org/10.3390/ijms18081581