BRS1 Function in Facilitating Lateral Root Emergence in Arabidopsis


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
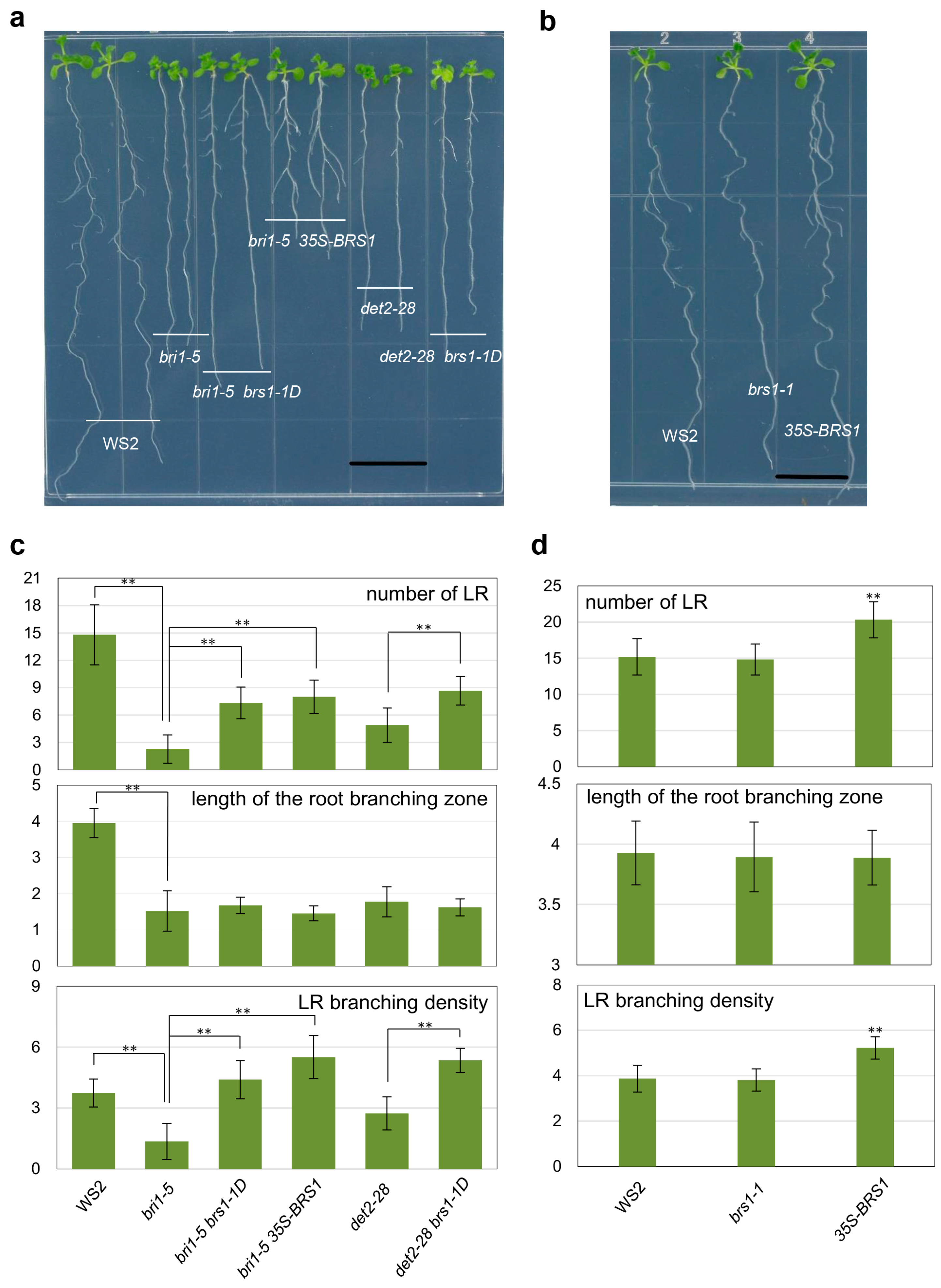
2.1. Overexpression of BRS1 Increases the Number of LRs in Arabidopsis
2.2. Overexpression of BRS1 Promotes LR Emergence
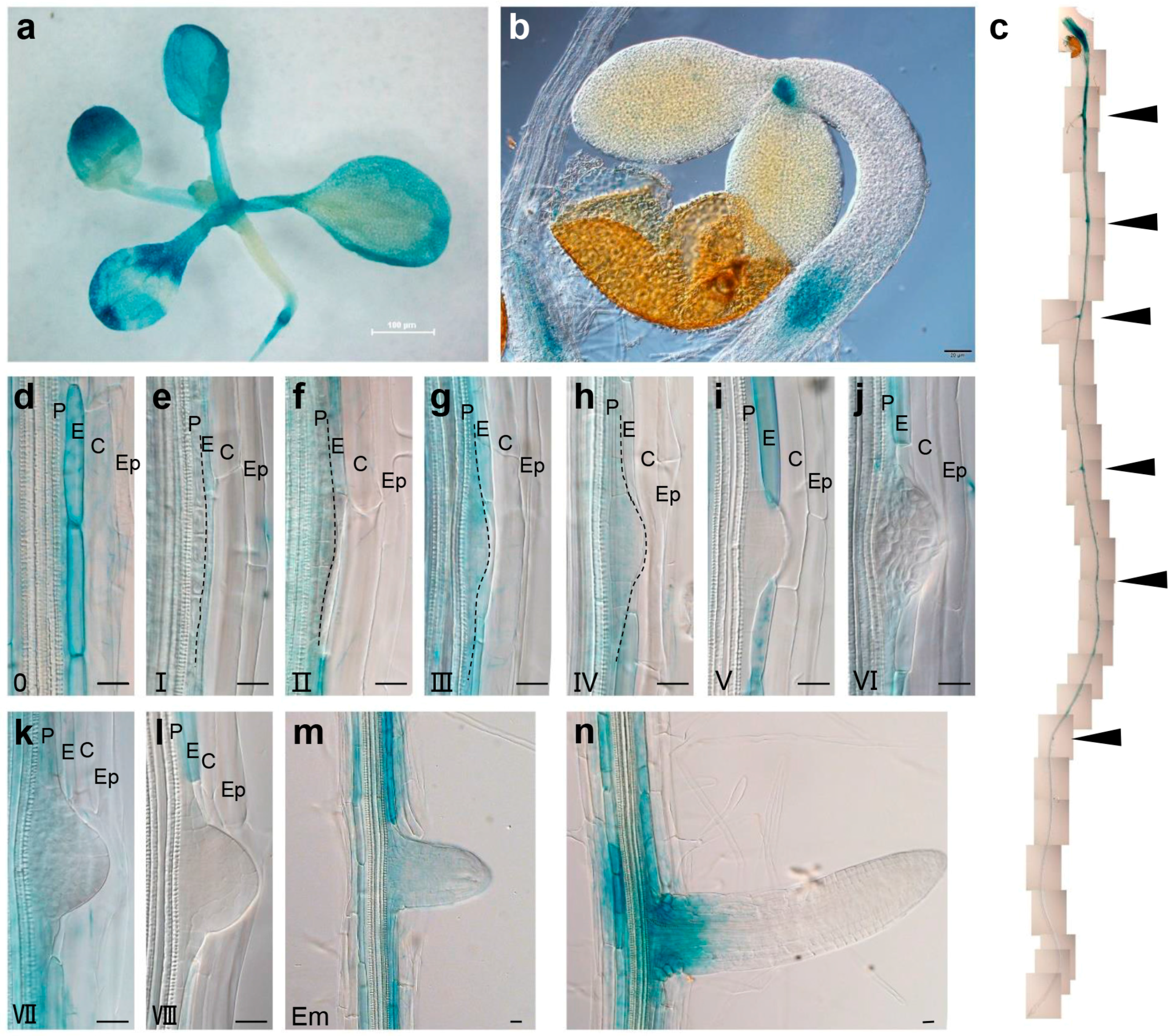
2.3. BRS1 Is Highly Expressed in the Root Endodermis
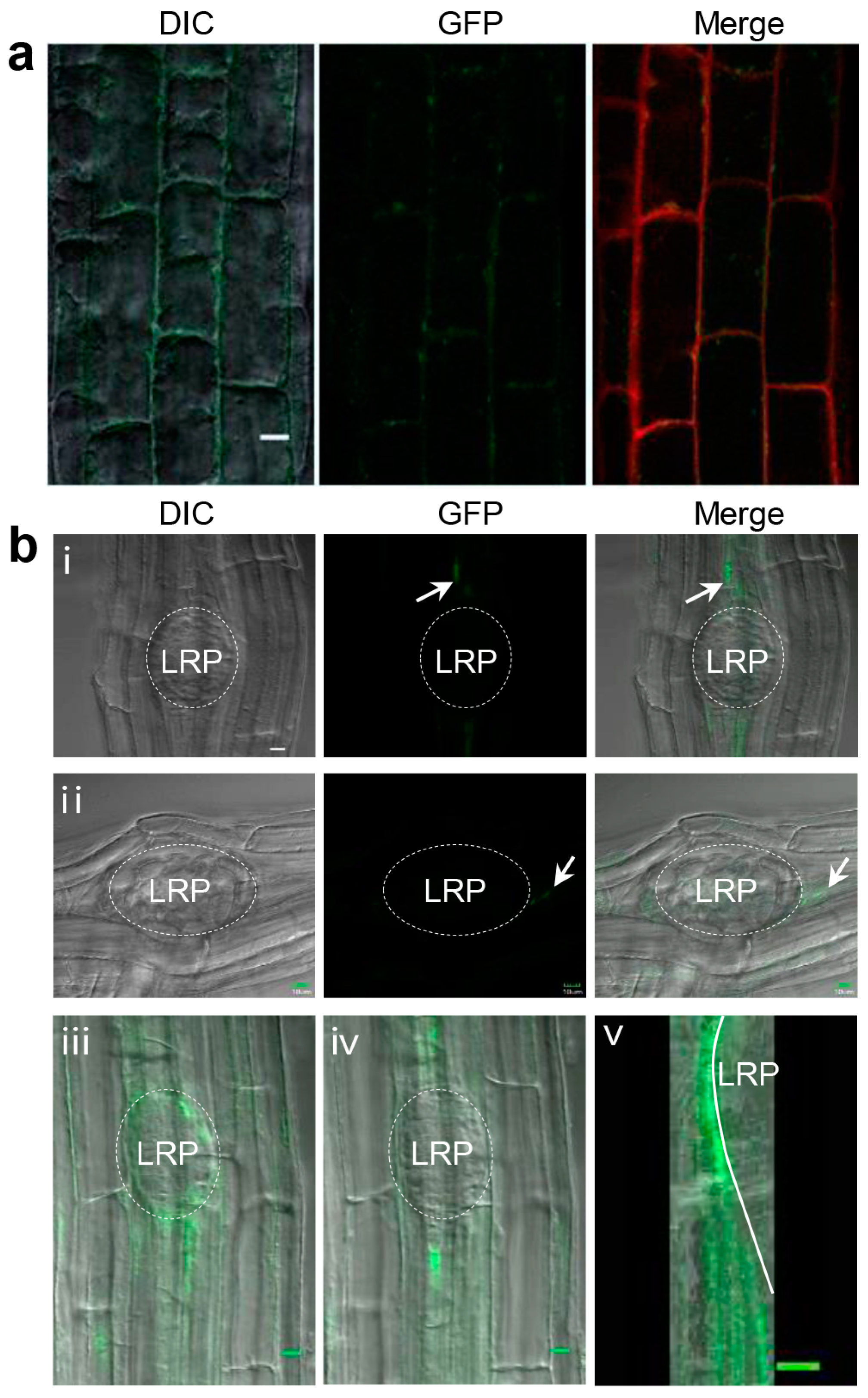
2.4. BRS1 Localizes in the Extracellular Space around the LRP
2.5. Transcription of BRS1 Is Regulated by the Brassinosteroid (BR) Signaling
3. Discussion
4. Materials and Methods
4.1. Growth Conditions and Plant Materials
4.2. RNA Isolation and Quantitative Real-Time RT-PCR Analyses
4.3. Histochemical β-Glucuronidase Assays
4.4. Microscopic Analysis
4.5. LRP Developmental Observations
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, J.; Lease, K.A.; Tax, F.E.; Walker, J.C. BRS1, a serine carboxypeptidase, regulates BRI1 signaling in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2001, 98, 5916–5921. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, R.; Moore, S.; Stein, W.H. Serine at the active center of yeast carboxypeptidase. J. Biol. Chem. 1973, 248, 8366–8369. [Google Scholar] [PubMed]
- Moura, D.S.; Bergey, D.R.; Ryan, C.A. Characterization and localization of a wound-inducible type I serine-carboxypeptidase from leaves of tomato plants (Lycopersicon esculentum Mill.). Planta 2001, 212, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Granat, S.J.; Wilson, K.A.; Tan-Wilson, A.L. New serine carboxypeptidase in mung bean seedling cotyledons. J. Plant Physiol. 2003, 160, 1263–1266. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, F.; Cejudo, F.J. Germination-related genes encoding proteolytic enzymes are expressed in the nucellus of developing wheat grains. Plant J. 1998, 15, 569–574. [Google Scholar] [CrossRef]
- Domínguez, F.; González, M.; Cejudo, F.J. A germination-related gene encoding a serine carboxypeptidase is expressed during the differentiation of the vascular tissue in wheat grains and seedlings. Planta 2002, 215, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Cercós, M.; Urbez, C.; Carbonell, J. A serine carboxypeptidase gene (PsCP), expressed in early steps of reproductive and vegetative development in Pisum sativum, is induced by gibberellins. Plant Mol. Biol. 2003, 51, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Milkowski, C.; Strack, D. Serine carboxypeptidase-like acyltransferases. Phytochemistry 2004, 65, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Schaller, A. A cut above the rest: The regulatory function of plant proteases. Planta 2004, 220, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tang, L.; Qiu, J.; Zhang, W.; Wang, Y.; Tong, X.; Wei, X.; Hou, Y.; Zhang, J. Serine carboxypeptidase 46 Regulates Grain Filling and Seed Germination in Rice (Oryza sativa L.). PLoS ONE 2016, 11, e0159737. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.P.; Sowdhamini, R. Cross genome comparisons of serine proteases in Arabidopsis and rice. BMC Genom. 2006, 7, 200. [Google Scholar] [CrossRef] [PubMed]
- Breddam, K.; Sørensen, S.B.; Ottesen, M. Isolation of carboxypeptidase II from malted barley by affinity chromatography. Carlsberg Res. Commun. 1985, 50, 199–209. [Google Scholar] [CrossRef]
- Breddam, K.; Sørensen, S.B.; Svendsen, I. Primary structure and enzymatic properties of carboxypeptidase II from wheat bran. Carlsberg Res. Commun. 1987, 52, 297. [Google Scholar] [CrossRef]
- Doan, N.P.; Fincher, G. The A-and B-chains of carboxypeptidase I from germinated barley originate from a single precursor polypeptide. J. Biol. Chem. 1988, 263, 11106–11110. [Google Scholar] [PubMed]
- Wen, J.; Li, J.; Walker, J.C. Overexpression of a serine carboxypeptidase increases carpel number and seed production in Arabidopsis thaliana. Food Energy Secur. 2012, 1, 61–69. [Google Scholar] [CrossRef]
- Zhou, A.; Li, J. Arabidopsis BRS1 is a secreted and active serine carboxypeptidase. J. Biol. Chem. 2005, 280, 35554–35561. [Google Scholar] [CrossRef] [PubMed]
- Bienert, M.D.; Delannoy, M.; Navarre, C.; Boutry, M. NtSCP1 from tobacco is an extracellular serine carboxypeptidase III that has an impact on cell elongation. Plant Physiol. 2012, 158, 1220–1229. [Google Scholar] [CrossRef] [PubMed]
- Dolan, L.; Janmaat, K.; Willemsen, V.; Linstead, P.; Poethig, S.; Roberts, K.; Scheres, B. Cellular organisation of the Arabidopsis thaliana root. Development 1993, 119, 71–84. [Google Scholar] [PubMed]
- Casimiro, I.; Beeckman, T.; Graham, N.; Bhalerao, R.; Zhang, H.; Casero, P.; Sandberg, G.; Bennett, M.J. Dissecting Arabidopsis lateral root development. Trends Plant Sci. 2003, 8, 165–171. [Google Scholar] [CrossRef]
- Péret, B.; de Rybel, B.; Casimiro, I.; Benková, E.; Swarup, R.; Laplaze, L.; Beeckman, T.; Bennett, M.J. Arabidopsis lateral root development: An emerging story. Trends Plant Sci. 2009, 14, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Malamy, J.E.; Benfey, P.N. Organization and cell differentiation in lateral roots of Arabidopsis thaliana. Development 1997, 124, 33–44. [Google Scholar] [PubMed]
- Dubrovsky, J.G.; Rost, T.L.; Colón-Carmona, A.; Doerner, P. Early primordium morphogenesis during lateral root initiation in Arabidopsis thaliana. Planta 2001, 214, 30–36. [Google Scholar] [CrossRef] [PubMed]
- De Smet, I. Lateral root initiation: One step at a time. New Phytol. 2012, 193, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, M.; Biller, S.; Stanley, K.; Kajstura, T.; Prusty, R. Expression profiling of auxin-treated Arabidopsis roots: Toward a molecular analysis of lateral root emergence. Plant Cell Physiol. 2006, 47, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Kumpf, R.P.; Shi, C.L.; Larrieu, A.; Stø, I.M.; Butenko, M.A.; Péret, B.; Riiser, E.S.; Bennett, M.J.; Aalen, R.B. Floral organ abscission peptide IDA and its HAE/HSL2 receptors control cell separation during lateral root emergence. Proc. Natl. Acad. Sci. USA 2013, 110, 5235–5240. [Google Scholar] [CrossRef] [PubMed]
- Vermeer, J.E.; von Wangenheim, D.; Barberon, M.; Lee, Y.; Stelzer, E.H.; Maizel, A.; Geldner, N. A spatial accommodation by neighboring cells is required for organ initiation in Arabidopsis. Science 2014, 343, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Naseer, S.; Lee, Y.; Lapierre, C.; Franke, R.; Nawrath, C.; Geldner, N. Casparian strip diffusion barrier in Arabidopsis is made of a lignin polymer without suberin. Proc. Natl. Acad. Sci. USA 2012, 109, 10101–10106. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Rubio, M.C.; Alassimone, J.; Geldner, N. A mechanism for localized lignin deposition in the endodermis. Cell 2013, 153, 402–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilches-Barro, A.; Maizel, A. Talking through walls: Mechanisms of lateral root emergence in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2015, 23, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Goh, T.; Kasahara, H.; Mimura, T.; Kamiya, Y.; Fukaki, H. Multiple AUX/IAA-ARF modules regulate lateral root formation: The role of Arabidopsis SHY2/IAA3-mediated auxin signalling. Philos. Trans. R. Soc. B 2012, 367, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Porco, S.; Larrieu, A.; Du, Y.; Gaudinier, A.; Goh, T.; Swarup, K.; Swarup, R.; Kuempers, B.; Bishopp, A.; Lavenus, J. Lateral root emergence in Arabidopsis is dependent on transcription factor LBD29 regulation of auxin influx carrier LAX3. Development 2016, 143, 3340–3349. [Google Scholar] [CrossRef] [PubMed]
- Dubrovsky, J.G.; Forde, B.G. Quantitative analysis of lateral root development: Pitfalls and how to avoid them. Plant Cell 2012, 24, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, S.; Li, J.; Choi, Y.H.; Seto, H.; Takatsuto, S.; Noguchi, T.; Watanabe, T.; Kuriyama, H.; Yokota, T.; Chory, J. The Arabidopsis deetiolated2 mutant is blocked early in brassinosteroid biosynthesis. Plant Cell 1997, 9, 1951–1962. [Google Scholar] [CrossRef] [PubMed]
- Ditengou, F.A.; Teale, W.D.; Kochersperger, P.; Flittner, K.A.; Kneuper, I.; van der Graaff, E.; Nziengui, H.; Pinosa, F.; Li, X.; Nitschke, R. Mechanical induction of lateral root initiation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2008, 105, 18818–18823. [Google Scholar] [CrossRef] [PubMed]
- Lucas, M.; Godin, C.; Jay-Allemand, C.; Laplaze, L. Auxin fluxes in the root apex co-regulate gravitropism and lateral root initiation. J. Exp. Bot. 2008, 59, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Péret, B.; Li, G.; Zhao, J.; Band, L.R.; Voss, U.; Postaire, O.; Luu, D.T.; da Ines, O.; Casimiro, I.; Lucas, M. Auxin regulates aquaporin function to facilitate lateral root emergence. Nat. Cell Biol. 2012, 14, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, H.; Hachez, C.; Bienert, M.D.; Beebo, A.; Swarup, K.; Voss, U.; Bouhidel, K.; Frigerio, L.; Schjoerring, J.K.; Bennett, M.J. Tonoplast aquaporins facilitate lateral root emergence. Plant Physiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Lucas, M.; Kenobi, K.; von Wangenheim, D.; Voss, U.; Swarup, K.; de Smet, I.; van Damme, D.; Lawrence, T.; Péret, B.; Moscardi, E. Lateral root morphogenesis is dependent on the mechanical properties of the overlaying tissues. Proc. Natl. Acad. Sci. USA 2013, 110, 5229–5234. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Lu, X.; Zi, Q.; Xun, Q.; Zhang, J.; Wu, Y.; Shi, H.; Wei, Z.; Zhao, B.; Zhang, X. RGF1 INSENSITIVE 1 to 5, a group of LRR receptor-like kinases, are essential for the perception of root meristem growth factor 1 in Arabidopsis thaliana. Cell Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Enstone, D.E.; Peterson, C.A.; Ma, F. Root endodermis and exodermis: Structure, function, and responses to the environment. J. Plant Growth Regul. 2002, 21, 335–351. [Google Scholar] [CrossRef]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Q.; Wang, X.; Zhang, D.; Wang, X.; Feng, C.; Xu, S. BRS1 Function in Facilitating Lateral Root Emergence in Arabidopsis. Int. J. Mol. Sci. 2017, 18, 1549. https://doi.org/10.3390/ijms18071549
Deng Q, Wang X, Zhang D, Wang X, Feng C, Xu S. BRS1 Function in Facilitating Lateral Root Emergence in Arabidopsis. International Journal of Molecular Sciences. 2017; 18(7):1549. https://doi.org/10.3390/ijms18071549
Chicago/Turabian StyleDeng, Qian, Xue Wang, Dongzhi Zhang, Xiaoming Wang, Cuizhu Feng, and Shengbao Xu. 2017. "BRS1 Function in Facilitating Lateral Root Emergence in Arabidopsis" International Journal of Molecular Sciences 18, no. 7: 1549. https://doi.org/10.3390/ijms18071549
APA StyleDeng, Q., Wang, X., Zhang, D., Wang, X., Feng, C., & Xu, S. (2017). BRS1 Function in Facilitating Lateral Root Emergence in Arabidopsis. International Journal of Molecular Sciences, 18(7), 1549. https://doi.org/10.3390/ijms18071549