1. Introduction
Cardiovascular diseases represent the leading causes of mortality and morbidity worldwide and contribute remarkably to the increase of health care expenditures [
1]. Based on these considerations, addressing cardiovascular diseases requires concrete actions and, in particular, a strategy, both globally and nationally, which includes prevention and reduction of risk factors, and adequate surveillance and monitoring [
2]. Epidemiological studies have established the benefit of reducing low-density lipoprotein cholesterol (LDL-C) and triglycerides (TG) in diminishing cardiovascular disease events [
2,
3,
4,
5,
6]. In light of these studies, the use of lipid-lowering drugs, mainly statins, may represent a primary prevention strategy of excellence. The significance of this approach is supported by a recent work by McConnachie et al. showing that a five-year primary prevention treatment with statins leads to long term benefits, reducing coronary-causes of death, and diminishing first hospital admission for cardiovascular events [
7]. Statins are the first choice LDL-lowering drugs with a recognized universal efficacy in preventing vascular events. The recognition of statin efficacy is highlighted by a recent suggestion to expand the use of statins even in individuals with a low-risk profile [
6,
7]. Despite the enthusiasm, we have to pay attention to some uncertainty related to the non-achievement of the desired LDL-C targets in about 50% of the high-risk treated population, and the occurrence of cardiovascular events even in patients who achieve the LDL-C levels [
8,
9,
10]. These two aspects suggest the existence of genetic traits that might explain the susceptibility to develop hypercholesterolemia and, secondly, the inter-individual response under statin regimen.
Several genome-wide studies have highlighted the involvement of numerous genes in lipid metabolism, in particular controlling serum High Density Lipoprotein Cholesterol (HDL-C), LDL-C and TG [
11,
12]. These loci include common variants, which have been consistently associated with lipid levels in candidate-gene studies reported over the past decade. Currently, the most studied variants are the G to A base change affecting the splicing site of exon 13 in the 3-hydroxy-3-methylglutaryl coenzyme A (
HMG-CoA) reductase (rs3846662) and the arginine to tryptophan substitution at codon 719 in the Kinesin-like protein 6 gene (
KIF6, rs20455). HMG-CoA reductase is the rate-limiting enzyme in cholesterol biosynthesis and the LDL-C-lowering effect of statins are mediated through its inhibition [
13]. KIF6 belongs to a superfamily of motor proteins (kinesins) that act upon microtubules as intracellular transport of cargo, including membrane organelles, protein complexes and mRNAs within cells [
14]. KIF6 is not directly involved in lipid metabolism; however, several studies reported the association between
KIF6 gene polymorphisms, coronary heart disease and events reduction from statin therapy [
15,
16,
17,
18,
19,
20,
21]. All these considerations prompted us to perform a pharmacogenetic study to determine whether individuals’ polymorphisms in
HMG-CoA reductase and
KIF6 genes are independently associated with hypercholesterolemia, other lipid-associated traits and statin response in a sample of unselected individuals enrolled in the Brisighella Heart Study (BHeS). Given the complexity of the investigated traits, we selected other polymorphisms, in addition to the two previously described, already associated with serum lipids levels and/or cardiovascular events [
19,
22]. In the present study, we also explored association between the investigated polymorphisms and obesity. This last association is driven by the fact that obesity is among the cardiovascular risk traits, together with lipid levels. Moreover, obesity is a well-established contributor to the development of dyslipidemia [
23].
3. Discussion
Serum lipids are important determinants of cardiovascular events, strongly related to morbidity [
11,
24,
25,
26,
27,
28]. Therefore, screening for increased circulating lipid levels and early treatment with statins are a key strategy for cardiovascular event prevention in the clinical practice. Besides this, several studies have suggested that lipid phenotypes are linked to genetic factors. In view of this, many efforts have been made to identify potential lipid-associated loci that could contribute to increase the armamentarium of available preventive strategies. The identification of a genetic feature, associated with the lipid phenotype, has a pivotal role in stepping up targeted medical controls and allows prompt intervention. The lipid phenotype is certainly recognized as a complex trait, involving multiple genes, and with a significant gene-environment interaction [
27]. In the attempt to overcome the genotype-phenotype gap in complex diseases, candidate gene/pathway approaches have traditionally been used. The success of these approaches strongly depends upon the correctness of the initial choice, which may lead to miss (i.e., unselected genes) significant results. In this context, genome-wide association studies (GWAS) represent a powerful tool. Starting from 2008, we witnessed an explosive rise in the number of GWAS identifying common genetic variants associated with LDL-C [
11,
12,
28,
29,
30,
31,
32,
33]. Among the discovered hits, three studies [
12,
32,
33] reported the significant association of a variant in the
HMG-CoA gene (rs3846662) and LDL-C level. The result was further replicated in gene-candidate studies [
34,
35,
36]. Stimulated by all these studies, we replicated the finding on the
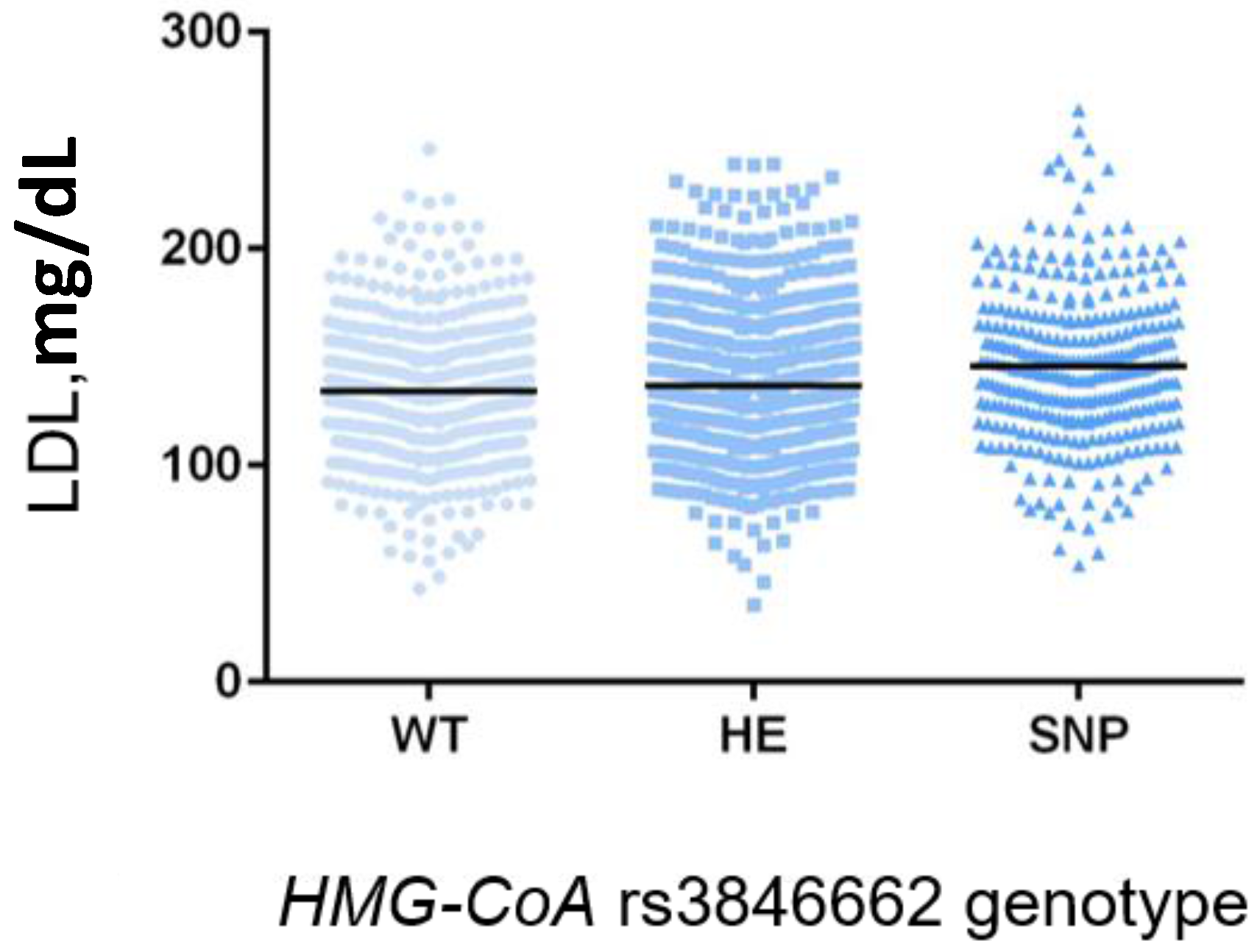
HMG-CoA polymorphisms rs3846662, in an independent and homogeneous Italian population. Overall, all the studies agree on the association of the
HMG-CoA-rs3846662-minor G allele with higher levels of LDL-C; in our study, in particular, the association was more evident in the homozygous GG genotype group. The polymorphism is a common variant located in intron 13 of the
HMG-CoA, affecting alternative splicing of the exon. Indeed, an in vitro study by Burkhardt and colleagues highlighted its functional role in modulating alternative splicing of HMG-CoA mRNA in human lymphoblastoid cells homozygous for the
HMG-CoA minor allele [
37]. In particular, the variant allele is associated with a significantly lower mRNA level of the alternatively spliced (Δ13 exon)
HMG-CoA. The authors speculated the polymorphism rs3846662 is located in a binding motif for a splice auxiliary protein, and allele status determines a change in the binding affinity of this protein. Consequently, a diminished HMG-CoA activity would promote a lower cellular cholesterol synthesis and, subsequently, an increase of cholesterol uptake from plasma through the LDL-receptor pathway to maintain intracellular cholesterol homeostasis [
37,
38]. The finding that the splice variant could not restore the enzyme activity when expressed in HMG-CoA deficient cells corroborates the functional role of the
HMG-CoA polymorphism (rs3846662). Despite its pharmacological interest, the exact molecular mechanism governing this process is largely unknown. Most probably, the presence of the A-major allele in homozygous increases the proportion of HMG-CoA mRNA lacking exon 13 [
37]. This alternative spliced variant is characterized by the lack of a portion of the catalytic domain (encoded by exon 13), and therefore may potentially affect stability of the enzyme [
37,
39,
40]. Most recently, it has been recognized the existence of specific splicing factors influencing stability, translation and structure of mRNAs, contributing to the observed phenotypic trait [
41]. In this context, the identification of HNRNPA1, a mRNA splicing regulator modulating the expression of HMG-CoA in an allele-related manner, and altering the RNA stability [
40], takes on a great importance in pharmacology. Specifically, HNRNPA1 promotes
HMG-CoA exon 13 skipping, more predominantly in the presence of the A-major allele, which also directly contributes to this exon-skipping phenomenon.
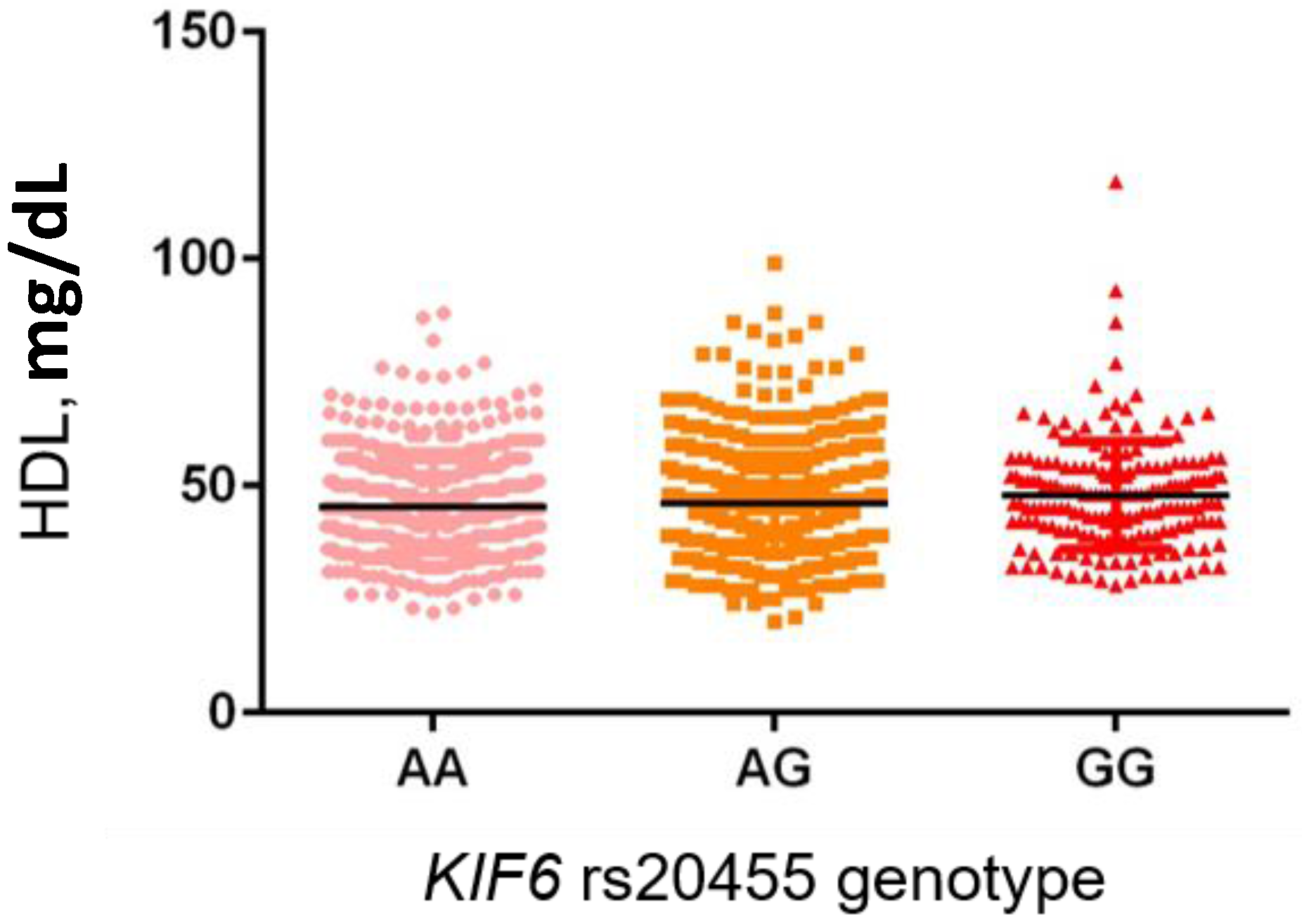
Besides the
HMG-CoA polymorphism, we also observed a correlation between
KIF6 rs20455 and HDL level; in particular, homozygous carriers of the G-minor allele showed higher serum HDL-C. The rs20455 variant is a non-synonymous polymorphism, leading to the replacement of a non-polar Trp with a basic Arg at codon 719, widely screened in relation to cardiovascular events, serum lipids and statin treatment outcome in various populations, with contrasting results [
20,
21,
42,
43,
44,
45,
46,
47]. To the best of our knowledge, our study is the first reporting an association with the KIF6 719Arg variant and HDL-C, while no apparent difference or opposite results were observed in previous studies [
20,
21,
42,
43,
44,
45,
46,
47]. Interestingly,
KIF6 polymorphisms (rs20455, rs9471077 and 9462535) were associated with risk of obesity, calculated as waist circumference, but in males only. Currently, obesity represents one of the major challenges and a public health problem worldwide. Adiposity, in particular abdominal obesity, is recognized as the key contributor to diabetes and other chronic diseases, and as a risk factor for hypercholesterolemia and further cardiovascular risk traits [
27,
28,
29]. Several studies have suggested that obesity is linked to genetic and environmental factors, and identifying genetic traits, including polymorphisms, may point to dysregulated genomic pathways. Recently, GWAS studies have identified several loci associated with obesity, mostly defined according to BMI values; however, none of them reported any association with
KIF6 variants [
48,
49,
50,
51,
52]. In our study, we found no correlation between any of the analyzed genetic variants and BMI, while we identified an interesting gene-gender interaction assessing obesity through waist circumference. Appropriate measures to prevent traits associated with cardiovascular events (as diabetes, dyslipidemia, hypertension, etc.) are mostly taken according to BMI anthropometric measures, aimed at identifying obese individuals. However, in the last decade waist circumference has emerged as a more accurate predictor of the metabolic risk of obesity compared to BMI [
53,
54,
55,
56,
57]. Based on these considerations, we analyzed men and women separately, as sex-specific cutoffs for waist circumference should be used to identify increased risk associated with abdominal fat. Our results showed that
KIF6 variants might confer risk of obesity in men only. This finding is interesting as the importance of gender in cardiovascular diseases has been widely described [
58,
59], and
KIF6 variants might potentially explain the gender-related differences in atherosclerosis development.
KIF6 encodes an intracellular protein involved in cellular cargo transportation, also expressed in coronary endothelial cells [
60], intuitively pointing to a possible role in cardiovascular function. Diverse variants within this gene may differently alter intracellular transport in endothelial cells in men and women, predisposing to coronary endothelial dysfunction and coronary heart disease. Recently, a study by Yoshino and co-workers reported a significant association between
KIF6 rs20456 and coronary epicardial endothelial dysfunction in men only [
61], while, in contrast, a previous study showed association between
KIF6 rs20455 and increased risk of coronary heart disease and myocardial infarction in Han Chinese women [
43]. Overall, to establish with certainty if our gender-specific finding reflects true-causal association, rather than a chance result, further studies deepening the underlying mechanism are required. Indeed, the homogeneous population and uniformity of life-style, minimizing population bias, points to a possible genetic locus for obesity in men. However, the association between
KIF6 polymorphisms and obesity needs confirmation or refutation in additional studies.
With regard to lipid-lowering therapy with statins, we did not find any association between
HMG-CoA or
KIF6 genotypes and achievement of <130 mg/dL LDL-C level. The data on KIF6 are in disagreement with the literature, as several studies have shown that
KIF6 719Arg allele carriers might have a greater lipid-lowering response from statin therapy compared to non-carriers [
15,
16,
17,
44,
62,
63,
64]. Furthermore, some (but not all) KIF6 719Arg carriers undergoing statin therapy have shown a greater reduction of coronary heart disease compared to non-carriers [
17,
18]. Although these findings have been debated, a
KIF6 (rs20455) genetic test is commercially available to physicians and could be routinely included in the clinical practice. A recent study has highlighted that provision of
KIF6-testing results and supporting information directly to patients was associated with significantly higher adherence and persistence to statin therapy over six months of follow-up [
46]. In the era of personalized medicine, this study suggests that pharmacogenetic testing approaches may represent a promising tool in the promotion of healthy patient behaviors, which is then translated into clinical utility (i.e., higher statin adherence and persistence). A different reason might be why we have not observed any association between
KIF6 genotypes and response to statin therapy. In particular, important limitations are: (i) sample size; (ii) statin responsiveness definition; (iii) kind of administered statin. With regard to the sample size, only 183 subjects out of 965 hypercholesterolemics were under statin therapy, greatly reducing the statistical power. However, this distribution is representative of treated subjects in the whole BHeS group where the decision to treat hypercholesterolemia is left to local General Physicians. Moreover, some cases of hypercholesterolemia were firstly diagnosed at the time of this population survey. Then, statin treatment is not considered as a first line for hypercholesterolemia in subjects at low global cardiovascular disease risk [
65]. Regarding responsiveness, we used achievement of LDL-C <130 mg/dL as a measure of response, while a more appropriate analysis would be to consider percentage reduction from baseline. However, the study started in 1972, and participants were evaluated every 4 years, so that a reliable baseline level is impossible to establish. An alternative analysis would be considering percentage reduction from the previous BheS survey. However, this approach is not properly correct, as in a four-year interval, it is difficult to establish the adherence to statin therapy and, therefore, the lipid levels observed. Alternatively, we could measure participants’ lipid levels 6 months prior to the BheS follow-up; however, this option was not scheduled, therefore we cannot calculate percentage reduction. Lastly, concerning the issue of the kind of administered statins, to date, benefit of statin therapy for
KIF6 carriers has been demonstrated for atorvastatin and pravastatin treatment only. In our study, it is difficult to draw a pharmacogenetic conclusion, as subjects under lipid-lowering treatment are heterogeneous in terms of the type of statin and dosage.
In conclusion, our results showed important associations between HMG-CoA rs3846662 and KIF6 rs20455 and lipid phenotypes, which may have an influence on dyslipidemia-related events. Next to this, to the best of our knowledge, this represents the first study implicating KIF6 variants with obesity in men only, and point to the possible involvement of this genetic locus in the known gender-related differences in coronary artery disease risk.

 ,
, 








{kind=link}
{kind=link}
{kind=link}