Emerging Roles of TWIK-1 Heterodimerization in the Brain


Abstract
:1. Introduction
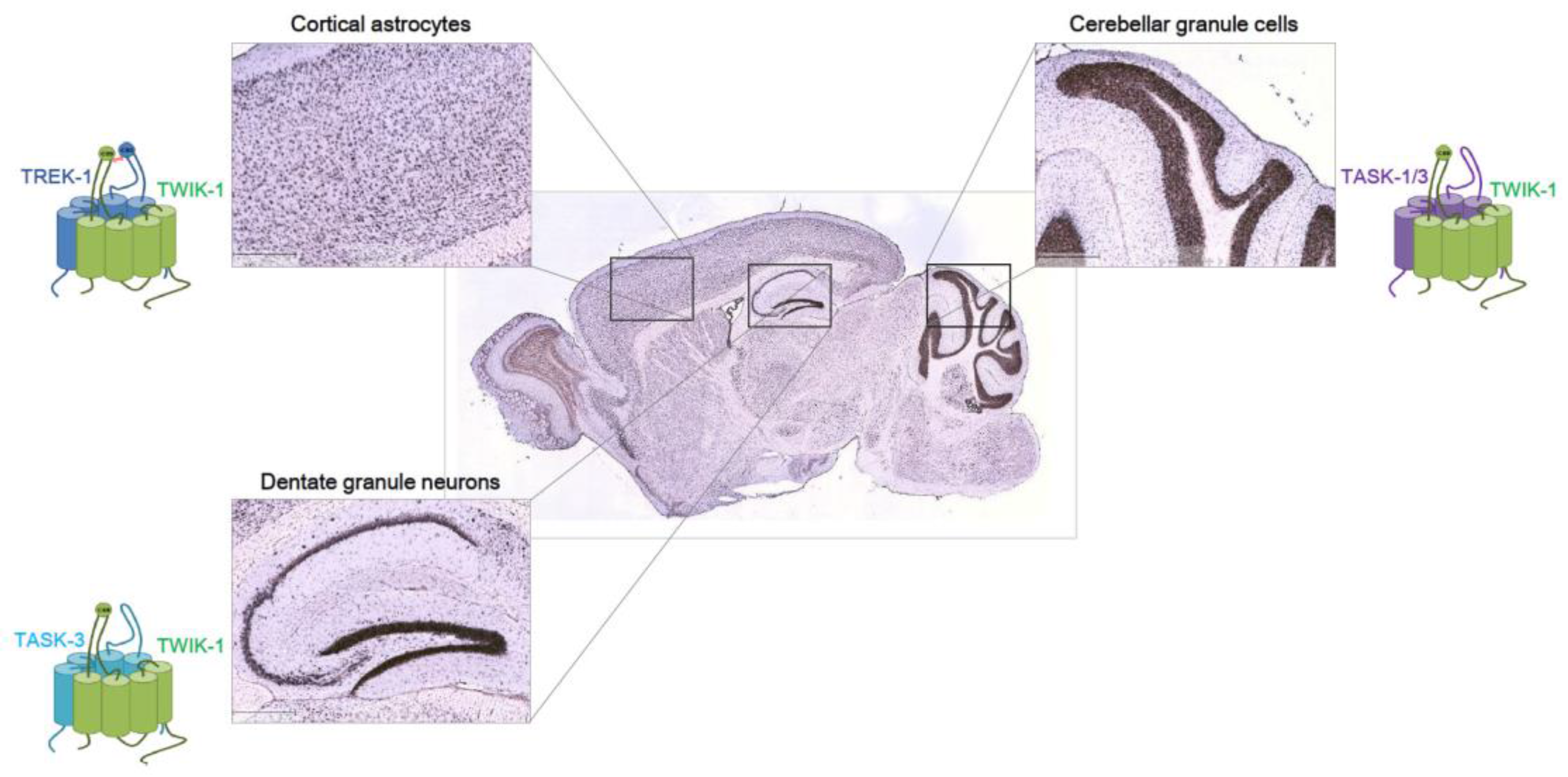
2. Distribution of TWIK-1 in the Brain
3. Heterodimerization between TWIK-1 and Other K2P Isotypes
4. Neuronal Function of TWIK-1
5. TWIK-1 in Astrocytic Passive Conductance
6. TWIK-1 in Astrocytic Glutamate Release
7. Implication of TWIK-1 in the central nervous system (CNS)/Brain Diseases
8. Perspectives
Acknowledgments
Conflicts of Interest
Abbreviations
| TWIK | tandem of pore domains in a weak inward rectifying K+ channel |
| TREK | TWIK-related K+ channel |
| TASK | TWIK-related acid-sensitive K+ channel |
| TRAAK | TWIK-related arachidonic acid-stimulated K+ channel |
| TALK | TWIK-related alkaline pH-activated K channel |
| THIK | tandem pore domain halothane-inhibited K+ channel |
| CHO | chinese hamster ovary |
| 5-HT1A | serotonin 1A receptor |
| CNS | central nervous system |
| Aβ | amyloid β |
References
- Enyedi, P.; Czirják, G. Molecular background of leak K+ currents: Two-pore domain potassium channels. Physiol. Rev. 2010, 90, 559–605. [Google Scholar] [CrossRef] [PubMed]
- Lesage, F.; Guillemare, E.; Fink, M.; Duprat, F.; Lazdunski, M.; Romey, G.; Barhanin, J. TWIK-1, a ubiquitous human weakly inward rectifying K+ channel with a novel structure. EMBO J. 1996, 15, 1004–1011. [Google Scholar] [PubMed]
- Rajan, S.; Plant, L.D.; Rabin, M.L.; Butler, M.H.; Goldstein, S.A. Sumoylation silences the plasma membrane leak K+ channel K2P1. Cell 2005, 121, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Feliciangeli, S.; Bendahhou, S.; Sandoz, G.; Gounon, P.; Reichold, M.; Warth, R.; Lazdunski, M.; Barhanin, J.; Lesage, F. Does sumoylation control K2P1/TWIK1 background K+ channels? Cell 2007, 130, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Plant, L.D.; Zuniga, L.; Araki, D.; Marks, J.D.; Goldstein, S.A. SUMOylation silences heterodimeric TASK potassium channels containing K2P1 subunits in cerebellar granule neurons. Sci. Signal. 2012, 5, ra84. [Google Scholar] [CrossRef] [PubMed]
- Plant, L.D.; Dementieva, I.S.; Kollewe, A.; Olikara, S.; Marks, J.D.; Goldstein, S.A. One SUMO is sufficient to silence the dimeric potassium channel K2P1. Proc. Natl. Acad. Sci. USA 2010, 107, 10743–10748. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Putra, A.; Schools, G.P.; Ma, B.; Chen, H.; Kaczmarek, L.K.; Barhanin, J.; Lesage, F.; Zhou, M. The contribution of TWIK-1 channels to astrocyte K+ current is limited by retention in intracellular compartments. Front. Cell. Neurosci. 2013, 7, 246. [Google Scholar] [CrossRef] [PubMed]
- Feliciangeli, S.; Tardy, M.P.; Sandoz, G.; Chatelain, F.C.; Warth, R.; Barhanin, J.; Bendahhou, S.; Lesage, F. Potassium channel silencing by constitutive endocytosis and intracellular sequestration. J. Biol. Chem. 2010, 285, 4798–4805. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Arrighi, I.; Kaissling, B.; Pfaff, I.; Mann, J.; Barhanin, J.; Vallon, V. Expression and insights on function of potassium channel TWIK-1 in mouse kidney. Pflugers Arch. Eur. J. Physiol. 2005, 451, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Chatelain, F.C.; Bichet, D.; Douguet, D.; Feliciangeli, S.; Bendahhou, S.; Reichold, M.; Warth, R.; Barhanin, J.; Lesage, F. TWIK1, a unique background channel with variable ion selectivity. Proc. Natl. Acad. Sci. USA 2012, 109, 5499–5504. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, X.; Chen, H. TWIK-1 two-pore domain potassium channels change ion selectivity and conduct inward leak sodium currents in hypokalemia. Sci. Signal. 2011, 4, ra37. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.H.; Chatelain, F.C.; Huttner, I.G.; Olesen, M.S.; Soka, M.; Feliciangeli, S.; Horvat, C.; Santiago, C.F.; Vandenberg, J.I.; Schmitt, N.; et al. The two-pore domain potassium channel, TWIK-1, has a role in the regulation of heart rate and atrial size. J. Mol. Cell. Cardiol. 2016, 97, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Lesage, F.; Lauritzen, I.; Duprat, F.; Reyes, R.; Fink, M.; Heurteaux, C.; Lazdunski, M. The structure, function and distribution of the mouse TWIK-1 K+ channel. FEBS Lett. 1997, 402, 28–32. [Google Scholar] [CrossRef]
- Arrighi, I.; Lesage, F.; Scimeca, J.C.; Carle, G.F.; Barhanin, J. Structure, chromosome localization, and tissue distribution of the TWIK K+ channel gene. FEBS Lett. 1998, 425, 310–316. [Google Scholar] [CrossRef]
- Talley, E.M.; Solorzano, G.; Lei, Q.; Kim, D.; Bayliss, D.A. CNS distribution of members of the two-pore-domain (KCNK) potassium channel family. J. Neurosci. 2001, 21, 7491–7505. [Google Scholar] [PubMed]
- Aller, M.I.; Wisden, W. Changes in expression of some two-pore domain potassium channel genes (KCNK) in selected brain regions of developing mice. Neuroscience 2008, 151, 1154–1172. [Google Scholar] [CrossRef] [PubMed]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A transcriptome database for astrocytes, neurons, and oligodendrocytes: A new resource for understanding brain development and function. J. Neurosci. 2008, 28, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Xu, G.; Xie, M.; Zhang, X.; Schools, G.P.; Ma, L.; Kimelberg, H.K.; Chen, H. TWIK-1 and TREK-1 are potassium channels contributing significantly to astrocyte passive conductance in rat hippocampal slices. J. Neurosci. 2009, 29, 8551–8564. [Google Scholar] [CrossRef] [PubMed]
- Benesova, J.; Rusnakova, V.; Honsa, P.; Pivonkova, H.; Dzamba, D.; Kubista, M.; Anderova, M. Distinct expression/function of potassium and chloride channels contributes to the diverse volume regulation in cortical astrocytes of GFAP/EGFP mice. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.M.; Kim, E.; Yarishkin, O.; Woo, D.H.; Han, K.S.; Park, N.; Bae, Y.; Woo, J.; Kim, D.; Park, M.; et al. A disulphide-linked heterodimer of TWIK-1 and TREK-1 mediates passive conductance in astrocytes. Nat. Commun. 2014, 5, 3227. [Google Scholar] [CrossRef] [PubMed]
- Lesage, F.; Reyes, R.; Fink, M.; Duprat, F.; Guillemare, E.; Lazdunski, M. Dimerization of TWIK-1 K+ channel subunits via a disulfide bridge. EMBO J. 1996, 15, 6400–6407. [Google Scholar] [PubMed]
- Miller, A.N.; Long, S.B. Crystal structure of the human two-pore domain potassium channel K2P1. Science 2012, 335, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.J.; Honoré, E. Properties and modulation of mammalian 2P domain K+ channels. Trends Neurosci. 2001, 24, 339–346. [Google Scholar] [CrossRef]
- Niemeyer, M.I.; Cid, L.P.; Valenzuela, X.; Paeile, V.; Sepúlveda, F.V. Extracellular conserved cysteine forms an intersubunit disulphide bridge in the KCNK5 (TASK-2) K+ channel without having an essential effect upon activity. Mol. Membr. Biol. 2003, 20, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Czirják, G.; Enyedi, P. Formation of functional heterodimers between the TASK-1 and TASK-3 two-pore domain potassium channel subunits. J. Biol. Chem. 2002, 277, 5426–5432. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.P.; Talley, E.M.; Manger, J.P.; Bayliss, D.A. Motoneurons express heteromeric TWIK-related acid-sensitive K+ (TASK) channels containing TASK-1 (KCNK3) and TASK-3 (KCNK9) subunits. J. Neurosci. 2004, 24, 6693–6702. [Google Scholar] [CrossRef] [PubMed]
- Blin, S.; Chatelain, F.C.; Feliciangeli, S.; Kang, D.; Lesage, F.; Bichet, D. Tandem Pore Domain Halothane-inhibited K+ Channel Subunits THIK1 and THIK2 Assemble and Form Active Channels. J. Biol. Chem. 2014, 289, 28202–28212. [Google Scholar] [CrossRef] [PubMed]
- Levitz, J.; Royal, P.; Comoglio, Y.; Wdziekonski, B.; Schaub, S.; Clemens, D.M.; Isacoff, E.Y.; Sandoz, G. Heterodimerization within the TREK channel subfamily produces a diverse family of highly regulated potassium channels. Proc. Natl. Acad. Sci. USA 2016, 113, 4194–4199. [Google Scholar] [CrossRef] [PubMed]
- Blin, S.; Ben Soussia, I.; Kim, E.J.; Brau, F.; Kang, D.; Lesage, F.; Bichet, D. Mixing and matching TREK/TRAAK subunits generate heterodimeric K2P channels with unique properties. Proc. Natl. Acad. Sci. USA 2016, 113, 4200–4205. [Google Scholar] [CrossRef] [PubMed]
- Lengyel, M.; Czirják, G.; Enyedi, P. Formation of Functional Heterodimers by TREK-1 and TREK-2 Two-pore Domain Potassium Channel Subunits. J. Biol. Chem. 2016, 291, 13649–13661. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Tsutsumi, K.; Miyamoto, T.; Yamamura, H.; Imaizumi, Y. Heterodimerization of two pore domain K+ channel TASK1 and TALK2 in living heterologous expression systems. PLoS ONE 2017, 12, e0186252. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.-Y.; Poudel, S.K.S.; Rojanathammanee, L.; Porter, J.E.; Lei, S. Serotonin inhibits neuronal excitability by activating two-pore domain K+ channels in the entorhinal cortex. Mol. Pharmacol. 2007, 72, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Brickley, S.G.; Revilla, V.; Cull-Candy, S.G.; Wisden, W.; Farrant, M. Adaptive regulation of neuronal excitability by a voltage-independent potassium conductance. Nature 2001, 409, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Yarishkin, O.; Lee, D.Y.; Kim, E.; Cho, C.H.; Choi, J.H.; Lee, C.J.; Hwang, E.M.; Park, J.Y. TWIK-1 contributes to the intrinsic excitability of dentate granule cells in mouse hippocampus. Mol. Brain 2014, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cotton, J.F.; Schuyler, J.A.; Fahlman, C.S.; Au, J.D.; Bickler, P.E.; Yost, C.S. Protective effects of TASK-3 (KCNK9) and related 2P K channels during cellular stress. Brain Res. 2005, 1031, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Marinc, C.; Derst, C.; Prüss, H.; Veh, R.W. Immunocytochemical localization of TASK-3 protein (K2P9.1) in the rat brain. Cell. Mol. Neurobiol. 2014, 34, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Woo, D.H.; Han, K.-S.; Shim, J.W.; Yoon, B.E.; Kim, E.; Bae, J.Y.; Oh, S.J.; Hwang, E.M.; Marmorstein, A.D.; Bae, Y.C.; et al. TREK-1 and Best1 channels mediate fast and slow glutamate release in astrocytes upon GPCR activation. Cell 2012, 151, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Badheka, D.; Yudin, Y.; Borbiro, I.; Hartle, C.M.; Yazici, A.; Mirshahi, T.; Rohacs, T. Inhibition of Transient Receptor Potential Melastatin 3 ion channels by G-protein βγ subunits. Elife 2017, 6, e26147. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.L.; Slesinger, P.A.; Casey, P.J.; Jan, Y.N.; Jan, L.Y. Evidence that direct binding of Gβγ to the GIRK1 G protein-gated inwardly rectifying K+ channel is important for channel activation. Neuron 1995, 15, 1133–1143. [Google Scholar] [CrossRef]
- Li, X.; Hümmer, A.; Han, J.; Herlitze, S. G protein β2 subunit-derived peptides for inhibition and induction of G protein pathways: Examination of voltage-gated Ca2+ and G protein inwardly rectifying K+ channels. J. Biol. Chem. 2005, 280, 23945–23959. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Orlandi, C.; Cao, Y.; Yang, S.; Choi, C.I.; Pagadala, V.; Birnbaumer, L.; Martemyanov, K.A.; Vardi, N. The TRPM1 channel in ON-bipolar cells is gated by both the α and the βγ subunits of the G-protein Go. Sci. Rep. 2016, 6, 20940. [Google Scholar] [CrossRef] [PubMed]
- Quallo, T.; Alkhatib, O.; Gentry, C.; Andersson, D.A.; Bevan, S. G protein βγ subunits inhibit TRPM3 ion channels in sensory neurons. Elife 2017, 6, e26138. [Google Scholar] [CrossRef] [PubMed]
- Dembla, S.; Behrendt, M.; Mohr, F.; Goecke, C.; Sondermann, J.; Schneider, F.M.; Schmidt, M.; Stab, J.; Enzeroth, R.; Leitner, M.G.; et al. Anti-nociceptive action of peripheral mu-opioid receptors by Gβγ protein-mediated inhibition of TRPM3 channels. Elife 2017, 6, e26280. [Google Scholar] [CrossRef] [PubMed]
- Matigian, N.; Windus, L.; Smith, H.; Filippich, C.; Pantelis, C.; McGrath, J.; Mowry, B.; Hayward, N. Expression profiling in monozygotic twins discordant for bipolar disorder reveals dysregulation of the WNT signalling pathway. Mol. Psychiatry 2007, 12, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Pindon, A.; Berry, M.; Hantaï, D. Thrombomodulin as a new marker of lesion-induced astrogliosis: Involvement of thrombin through the G-protein-coupled protease-activated receptor-1. J. Neurosci. 2000, 20, 2543–2550. [Google Scholar] [PubMed]
- Miyazaki, I.; Asanuma, M.; Murakami, S.; Takeshima, M.; Torigoe, N.; Kitamura, Y.; Miyoshi, K. Targeting 5-HT1A receptors in astrocytes to protect dopaminergic neurons in parkinsonian models. Neurobiol. Dis. 2013, 59, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Molinaro, G.; Battaglia, G. Targeting group II Metabotropic Glutamate (mGlu) Receptors for the Treatment of Psychosis Associated with Alzheimer’s Disease: Selective Activation of mGlu2 Receptors Amplifies β-Amyloid Toxicity in Cultured Neurons, Whereas Dual Activation of mGlu2 and mGlu3 Receptors is Neuroprotective. Mol. Pharmacol. 2011, 79, 618–626. [Google Scholar] [PubMed]
- Zhang, H.; Dong, H.; Cilz, N.I.; Kurada, L.; Hu, B.; Wada, E.; Bayliss, D.A.; Porter, J.E.; Lei, S. Neurotensinergic Excitation of Dentate Gyrus Granule Cells via Gαq-Coupled Inhibition of TASK-3 Channels. Cereb Cortex. 2016, 26, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Yarishkin, O.; Kim, E.; Bae, Y.; Kim, A.; Kim, S.C.; Ryoo, K.; Cho, C.H.; Hwang, E.M.; Park, J.Y. TWIK-1/TASK-3 heterodimeric channels contribute to the neurotensin-mediated excitation of hippocampal dentate gyrus granule cells. (unphublished: manuscript in preperation).


{kind=link}
| K2P Dimer | Validation | Disulfide Bond | Physiological Function | Ref. | |
|---|---|---|---|---|---|
| In Vitro | In Vivo | ||||
| TASK-1/TASK-3 | Co-IP | N.D. | The heterodimeric channels mediate the pH and isoflurane-sensitive K+ currents in hypoglossal motoneurons. | [26] | |
| TWIK-1/TASK-1 (or TASK-3) | FRET, Co-IP | N.D. | The heterodimeric channels comprise the acid-sensitive K+ currents and response to halothane in cerebellar granule cells. | [5] | |
| TWIK-1/TREK-1 | Co-IP, BiFC, MY2H | Co-IP, PLA | Dependent (TWIK-1 C69/ TREK-1 C93) | The heterodimeric channels mediate passive conductance and fast glutamate release in cortical astrocytes. | [20] |
| TWIK-1/TREK-2 | Co-IP | Dependent (TWIK-1 C69) | N.D. | [20] | |
| TWIK-1/TRAAK | Co-IP | Dependent (TWIK-1 C69) | N.D. | [20] | |
| THIK-1/THIK-2 | FRET, PLA | N.D. | N.D. | [27] | |
| TRAAK/TREK-1 (or TREK-2) | SiMPull TIRF imaging FRET PLA | N.D. | N.D. | [28,29] | |
| TREK-1/TREK-2 | Co-IP | Single channel recording | N.D. | N.D. | [30] |
| TASK-1/TALK-2 | BiFC, FRET, Co-IP | TIRF imaging | N.D. | N.D. | [31] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, C.-H.; Hwang, E.M.; Park, J.-Y. Emerging Roles of TWIK-1 Heterodimerization in the Brain. Int. J. Mol. Sci. 2018, 19, 51. https://doi.org/10.3390/ijms19010051
Cho C-H, Hwang EM, Park J-Y. Emerging Roles of TWIK-1 Heterodimerization in the Brain. International Journal of Molecular Sciences. 2018; 19(1):51. https://doi.org/10.3390/ijms19010051
Chicago/Turabian StyleCho, Chang-Hoon, Eun Mi Hwang, and Jae-Yong Park. 2018. "Emerging Roles of TWIK-1 Heterodimerization in the Brain" International Journal of Molecular Sciences 19, no. 1: 51. https://doi.org/10.3390/ijms19010051
APA StyleCho, C. -H., Hwang, E. M., & Park, J. -Y. (2018). Emerging Roles of TWIK-1 Heterodimerization in the Brain. International Journal of Molecular Sciences, 19(1), 51. https://doi.org/10.3390/ijms19010051