New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Multitemperature Single Stranded Conformational Polymorphism

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Phylogenetic Analysis
2.2. Granulin and Late Expression Factor-9 Nucleotide Sequence Alignment and Primers Design for PCR Reaction Prior to Multitemperature Single Stranded Conformational Polymorphism (MSSCP)
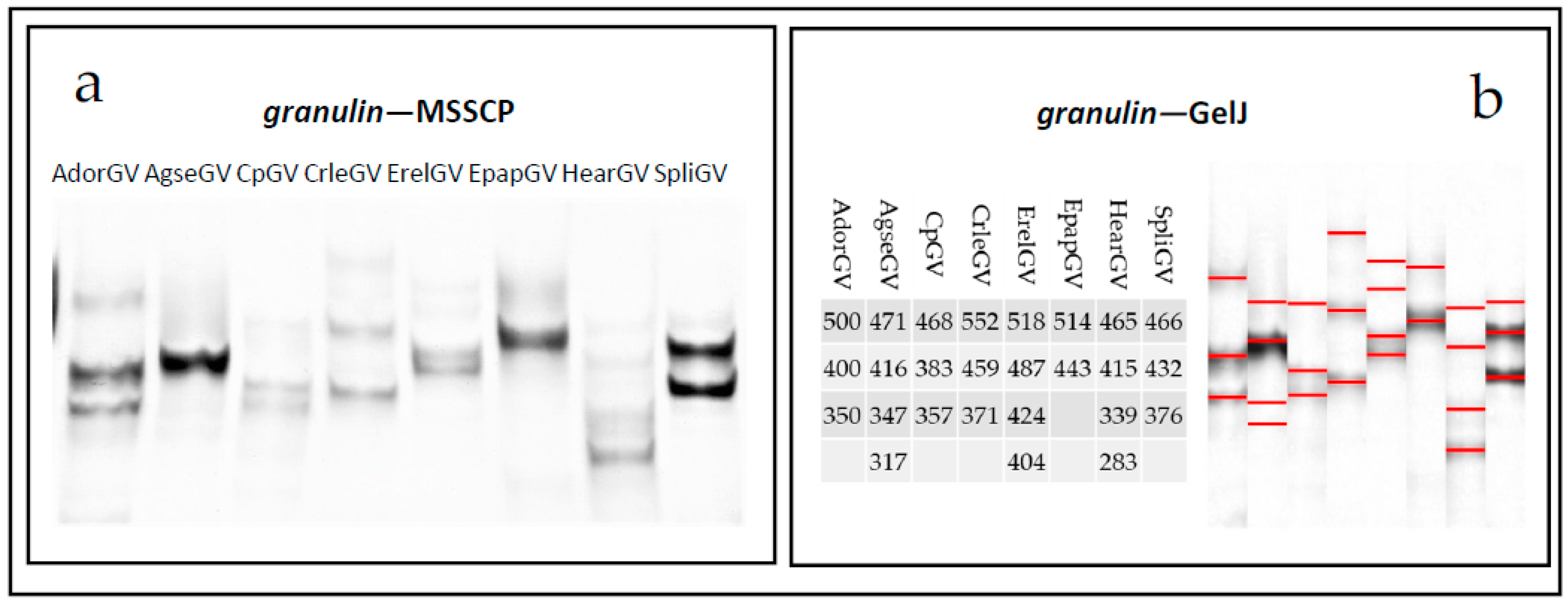
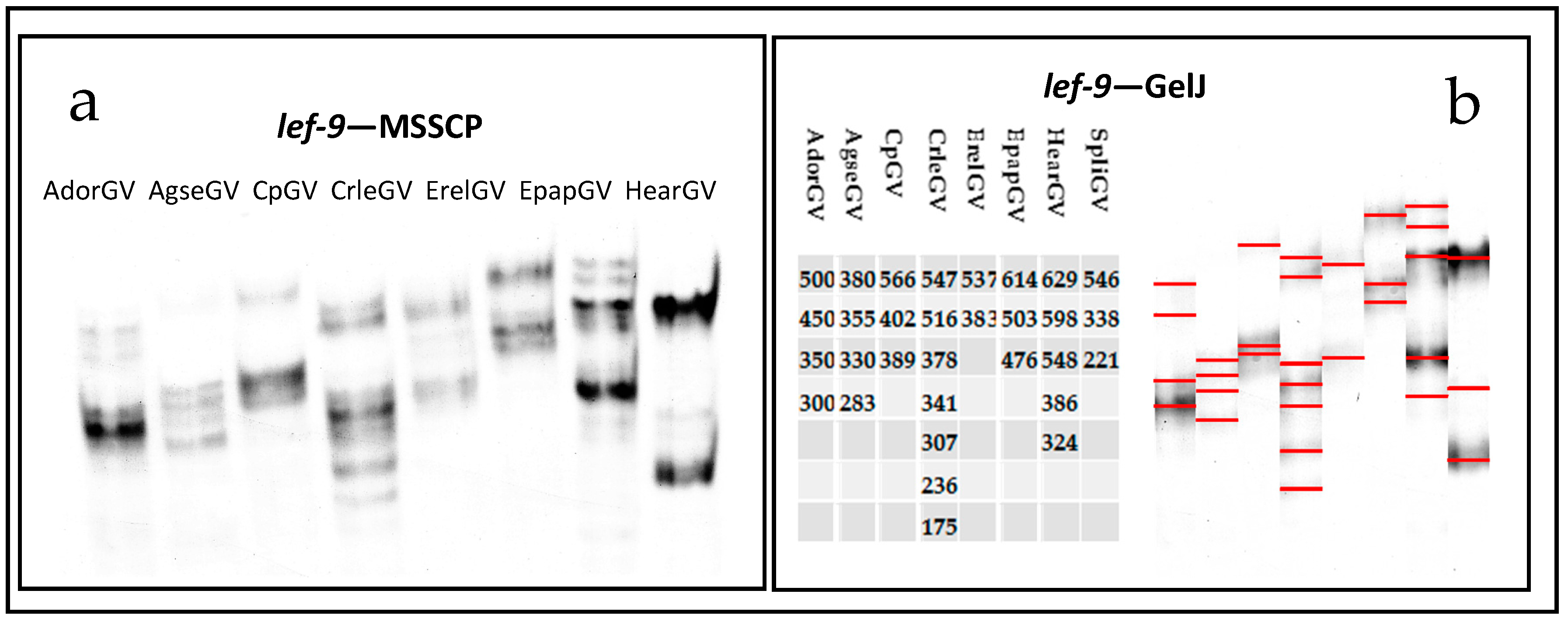
2.3. Multitemperature Single Stranded Conformational Polymorphism
3. Discussion
4. Materials and Methods
4.1. Phylogenetic Analyses of Betabaculovirus Full Genomes
4.2. Virus Purification and DNA Extraction
4.3. Granulin and Late Expression Factor-9 Nucleotide Sequences Alignment and Primer Design for PCR Reaction Prior to MSSCP
4.4. Polymerase Chain Reaction (PCR)
4.5. Multitemperature Single Stranded Conformational Polymorphism (MSSCP) Method
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| gran | granulin |
| lef-9 | late expression factor-9 |
| MSSCP | Multitemperature Single Stranded Conformation Polymorphism |
| NGS | Next Generation Sequencing |
| nt | nucleotide |
| OBs | occlusion bodies |
| PCR | Polymerase Chain Reaction |
| RFLP | Restriction Fragment Length Polymorphism |
| ssDNA | single stranded DNA |
References
- Szewczyk, B.; Hoyos-Carvajal, L.; Paluszek, M.; Skrzecz, I.; Lobo de Souza, M. Baculoviruses—Re-emerging biopesticides. Biotechnol. Adv. 2006, 24, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Beas-Catena, A.; Sanchez-Miron, A.; Garcia-Camacho, F.; Contreras-Gomez, A.; Molina-Grima, E. Baculoviruses biopesticides: An overview. J. Anim. Plant Sci. 2014, 24, 362–373. [Google Scholar]
- Blissard, G.W.; Rohrmann, G.F. Baculovirus diversity and molecular biology. Annu. Rev. Entomol. 1990, 35, 127–155. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Wang, H.; Zhihong, H.; Jehle, J.A. Towards a molecular identification and classification system of lepidopteran-specific baculoviruses. Virology 2004, 325, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, M.J.; Miele, S.A.B.; Iserte, J.A.; Belaich, M.N.; Ghiringhelli, P.D. The ac53, ac78, ac101, and ac103 genes are newly discovered core genes in the family Baculoviridae. J. Virol. 2012, 86, 12069–12079. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Miller, L.K. Identification of three late expression factor genes within the 33.8- to 43.4-map-unit region of Autographa californica nuclear polyhedrosis virus. J. Virol. 1994, 68, 6710–6718. [Google Scholar] [PubMed]
- Herniou, E.A.; Arif, B.M.; Becnel, J.J.; Blissard, G.W.; Bonning, B.C.; Harrison, R.; Jehle, J.A.; Theilmann, D.A.; Vlak, J.M. Baculoviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: New York, NY, USA, 2011. [Google Scholar]
- Miele, S.A.B.; Garavaglia, M.J.; Belaich, M.N.; Ghiringhelli, P.D. Baculovirus: Molecular Insights on Their Diversity and Conservation. Int. J. Evol. Biol. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Nakai, M.; Harrison, R.L.; Uchida, H.; Ukuda, R.; Hikihara, S.; Ishii, K.; Kunimi, Y. Isolation of an Adoxophyes orana granulovirus (AdorGV) occlusion body morphology mutant: Biological activity, genome sequence and relationship to other isolates of AdorGV. J. Gen. Virol. 2015, 96, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Dezianian, A.; Sajap, A.S.; Lau, W.H.; Omar, D.; Kadir, H.A.; Mohamed, R.; Yusoh, M.R.M. Morphological Characteristics of P. xylostella Granulovirus and Effects on Its Larval Host Diamondback Moth Plutella xylostella L. (Lepidoptera, Plutellidae). Am. J. Agric. Biol. Sci. 2010, 5, 43–49. [Google Scholar] [CrossRef]
- Kemp, E.M.; Woodward, D.T.; Cory, J.S. Detection of single and mixed covert baculovirus infections in eastern spruce budworm, Choristoneura fumiferana populations. J. Invertebr. Pathol. 2011, 107, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Cuartas, P.E.; Barrera, G.P.; Belaich, M.N.; Barreto, E.; Ghiringhelli, P.D.; Villamizar, L.F. The Complete Sequence of the First Spodoptera frugiperda Betabaculovirus Genome: A Natural Multiple Recombinant Virus. Viruses 2015, 7, 394–421. [Google Scholar] [CrossRef] [PubMed]
- Wennmann, J.T.; Jehle, J.A. Detection and quantitation of Agrotis baculoviruses in mixed infections. J. Virol. Methods 2014, 197, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Parola, A.D.; Sciocco-Cap, A.; Glikmann, G.; Romanowski, V. An immunochemical method for quantitation of Epinotia aporema granulovirus (EpapGV). J. Virol. Methods 2003, 112, 13–21. [Google Scholar] [CrossRef]
- Nadala, E.C.B., Jr.; Tapay, L.M.; Cao, S.; Loh, P.C. Detection of yellowhead virus and Chinese baculovirus in penaeid shrimp by the Western blot technique. J. Virol. Methods 1997, 69, 39–44. [Google Scholar] [CrossRef]
- Ebling, P.M.; Holmes, S.B. A refined method for the detection of baculovirus occlusion bodies in forest terrestrial and aquatic habitats. Pest Manag. Sci. 2002, 58, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Ramesh Kumar, D.; Sanjuktha, M.; Rajan, J.J.S.; Ananda Bharathi, R.; Santiago, T.C.; Alavandi, S.V.; Poornima, M. Development of SYBR Green based real time PCR assay for detection of monodon baculovirus in Penaeus monodon. J. Virol. Methods 2014, 205, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Murillo, R.; Muñoz, D.; Williams, T.; Mugeta, N.; Caballero, P. Application of the PCR-RFLP method for the rapid differentiation of Spodoptera exigua nucleopolyhedrovirus genotypes. J. Virol. Methods 2006, 135, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Christian, P.D.; Gibb, N.; Kasprzak, A.B.; Richards, A. A rapid method for the identification and differentiation of Helicoverpa nucleopolyhedroviruses (NPV Baculoviridae) isolated from the environment. J. Virol. Methods 2001, 96, 51–65. [Google Scholar] [CrossRef]
- Rabalski, L.; Krejmer-Rabalska, M.; Skrzecz, I.; Wasag, B.; Szewczyk, B. An alphabaculovirus isolated from dead Lymantria dispar larvae shows high genetic similarity to baculovirus previously isolated from Lymantria monacha—An example of adaptation to a new host. J. Invertebr. Pathol. 2016, 139, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, B.; Barski, P.; Sihler, W.; Rabalski, L.; Skrzecz, I.; Hoyos-Carvajal, L.; de Souza, M.L. Detection and identification of baculovirus pesticides by multitemperature single-strand conformational polymorphism. J. Environ. Sci. Health B 2008, 43, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Orita, M.; Iwahana, H.; Kanazawa, H.; Hayashi, K.; Sekiya, T. Detection of polymorphisms of human DNA by gel electrophoresis as single-strand conformation polymorphisms. Proc. Natl. Acad. Sci. USA 1989, 86, 2766–2770. [Google Scholar] [CrossRef] [PubMed]
- Kaczanowski, R.; Trzeciak, L.; Kucharczyk, K. Multitemperature single-strand conformation polymorphism. Electrophoresis 2001, 22, 3539–3545. [Google Scholar] [CrossRef]
- Heras, J.; Domínguez, C.; Mata, E.; Pascual, V.; Lozano, C.; Torres, C.; Zarazaga, M. GelJ—A tool for analyzing DNA fingerprint gel images. BMC Bioinform. 2015, 16, 270. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, B.; Lobo de Souza, M.; Batista de Castro, M.E.; Moscardi, F. Baculovirus biopesticides. In Pesticides—Formulations, Effects, Fate; Stoytcheva, M., Ed.; InTech Open Access Publisher: Rijeka, Croatia, 2011; Chapter 2; pp. 25–36. ISBN 978-953-307-532-7. [Google Scholar]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus Insecticides in Latin America: Historical Overview, Current Status and Future Perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Vijayendran, D.; Bonning, B.C. Next Generation Sequencing Technologies for Insect Virus Discovery. Viruses 2011, 3, 1849–1869. [Google Scholar] [CrossRef] [PubMed]
- Grzywacz, D. Basic and Applied Research: Baculovirus. In Microbial Control of Insect and Mite Pest. From Theory to Practice; Lacey, L.A., Ed.; Academic Press: London, UK, 2017; pp. 27–46. ISBN 978-0-12-803527-6. [Google Scholar]
- Huber, J. Western Europe. In Insect Viruses and Pest Management; Wiley and Sons: New York, NY, USA, 1998; pp. 201–215. [Google Scholar]
- Eberle, K.E.; Jehle, J.A. Field resistance of codling moth against Cydia pomonella granulovirus (CpGV) is autosomal and incompletely dominant inherited. J. Invertebr. Pathol. 2006, 93, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Eberle, K.E.; Asser-Kaiser, S.; Sayed, S.M.; Nguyen, H.T.; Jehle, J.A. Overcoming the resistance of codling moth against conventional Cydia pomonella granulovirus (CpGV-M) by a new isolate CpGV-I12. J. Invertebr. Pathol. 2008, 98, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Bisutti, I.L.; Ladurner, E.; Benuzzi, M.; Sauphanor, B.; Kienzle, J.; Zing, D.; Undorf-Spahn, D.; Fritsch, E.; Hüber, J.; et al. The occurrence and distribution of resistance of codling moth to Cydia pomonella granulovirus in Europe. J. Appl. Entomol. 2013, 137, 641–649. [Google Scholar] [CrossRef]
- Gwynn, R. Manual of Biocontrol Agents, 5th ed.; British Crop Protectin Council: Alton, UK, 2014; 520p. [Google Scholar]
- Moore, S.D.; Kirkman, W.; Richards, G.I.; Stephen, P.R. The Cryptophlebia Leucotreta Granulovirus—10 Years of Commercial Field Use. Viruses 2015, 7, 1284–1312. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.L.; Keena, M.A.; Rowley, D.L. Classification, genetic variation and pathogenicity of Lymantria dispar nuclepolyhedrovirus isolates from Asia, Europe and North America. J. Invertebr. Pathol. 2014, 116, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Licciardello, G.; Raspagliesi, D.; Bar-Joseph, M.; Catara, A. Characterization of isolates of Citrus tristeza virus by sequential analyses of enzyme immunoassays and capillary electrophoresis-single-strand conformation polymorphisms. J. Virol. Methods 2012, 181, 139–147. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baculovirus Designation | Betabaculovirus | Accession Number | Baculovirus Designation | Betabaculovirus | Accession Number |
|---|---|---|---|---|---|
| AdorGV | Adoxophyes orana GV | NC_005038 | ErelGV | Erynnyis ello GV | KJ406702 |
| AgseGV | Agrotis segetum GV | NC_005839 | HearGV | Helicoverpa armigera GV | NC_010240 |
| ChocGV | Choristoneura occidentalis GV | NC_008168 | PhopGV | Phthorimaea operculella GV | NC_004062 |
| ClanGV | Clostera anachoreta GV | NC_015398 | PrGV | Pieris rapae GV | NC_013797 |
| ClasGV | Clostera anastomosis GV | KR091910 | PlxyGV | Plutella xylosetella GV | NC_002593 |
| CaLGV-Henan | Clostera anastomosis GV (Henan isolate) | NC_022646 | PsunGV | Pseudaletia unipuncta GV | NC_013772 |
| CrleGV | Cryptophlebia leucotreta GV | NC_005068 | SfGV | Spodoptera frugiperda GV | NC_026511 |
| CpGV | Cydia pomonella GV | NC_002816 | SpliGV | Spodoptera litura GV | NC_009503 |
| EpapGV | Epinotia aporema GV | NC_005839 | XcGV | Xestia c-nigrum GV | NC_002331 |
| Gene | Forward | Reverse |
|---|---|---|
| gran | 5′TACATGGTBACNGARGA3′ | 5′AAYTCYTTNCCGCTCCAGTT3′ |
| lef-9 | 5′CARAACAARAAYGGRTAYGC3′ | 5′GGRTGNCGHGTGTTCCAYAC3′ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krejmer-Rabalska, M.; Rabalski, L.; Lobo de Souza, M.; Moore, S.D.; Szewczyk, B. New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Multitemperature Single Stranded Conformational Polymorphism. Int. J. Mol. Sci. 2018, 19, 83. https://doi.org/10.3390/ijms19010083
Krejmer-Rabalska M, Rabalski L, Lobo de Souza M, Moore SD, Szewczyk B. New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Multitemperature Single Stranded Conformational Polymorphism. International Journal of Molecular Sciences. 2018; 19(1):83. https://doi.org/10.3390/ijms19010083
Chicago/Turabian StyleKrejmer-Rabalska, Martyna, Lukasz Rabalski, Marlinda Lobo de Souza, Sean D. Moore, and Boguslaw Szewczyk. 2018. "New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Multitemperature Single Stranded Conformational Polymorphism" International Journal of Molecular Sciences 19, no. 1: 83. https://doi.org/10.3390/ijms19010083
APA StyleKrejmer-Rabalska, M., Rabalski, L., Lobo de Souza, M., Moore, S. D., & Szewczyk, B. (2018). New Method for Differentiation of Granuloviruses (Betabaculoviruses) Based on Multitemperature Single Stranded Conformational Polymorphism. International Journal of Molecular Sciences, 19(1), 83. https://doi.org/10.3390/ijms19010083