Regulation of Akt/FoxO3a/Skp2 Axis Is Critically Involved in Berberine-Induced Cell Cycle Arrest in Hepatocellular Carcinoma Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
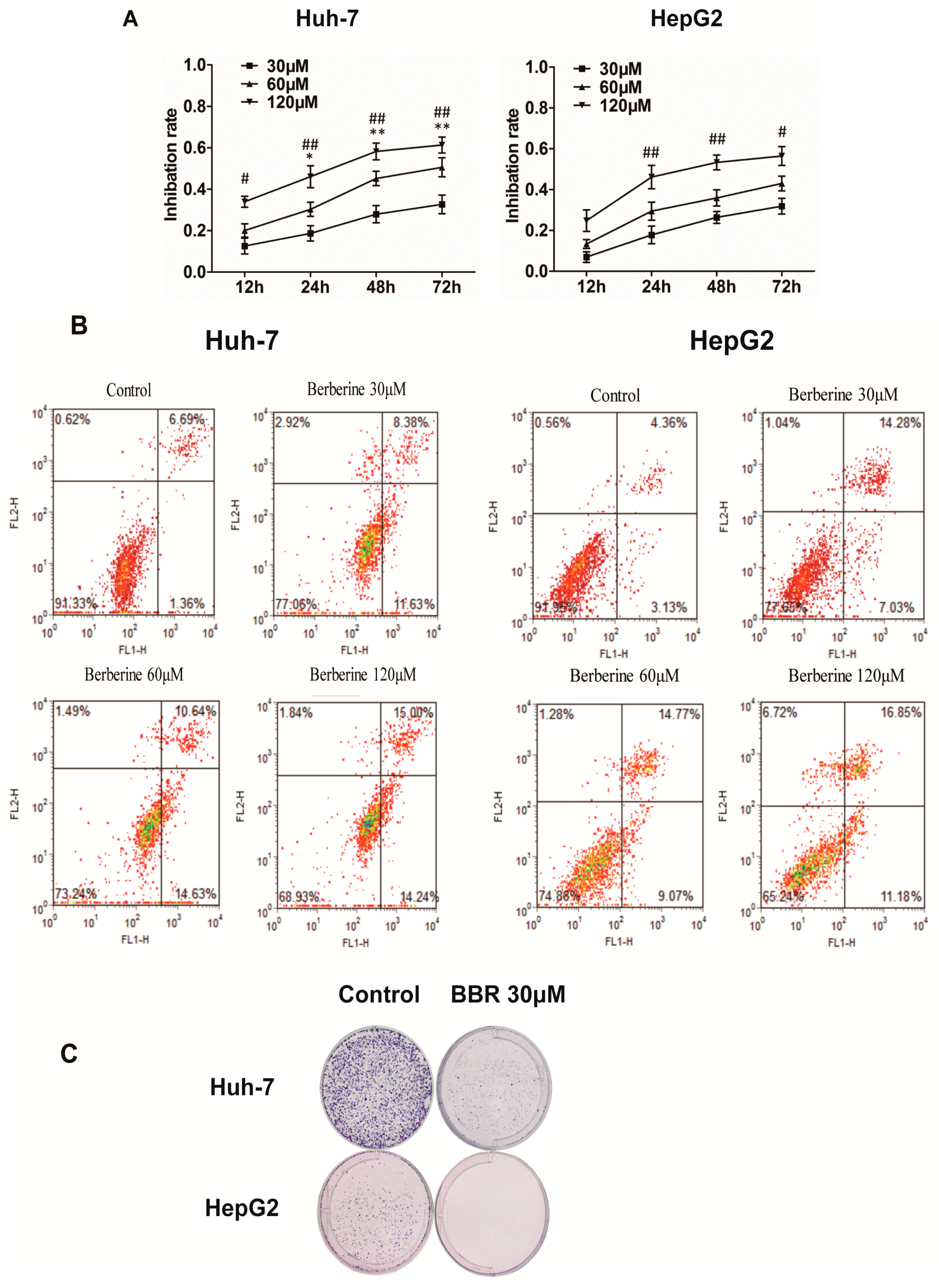
2.1. BBR Inhibits the Proliferation of Huh-7 and HepG2 Cells
2.2. BBR Inhibits Clonogenic Ability of Huh-7 and HepG2 Cells
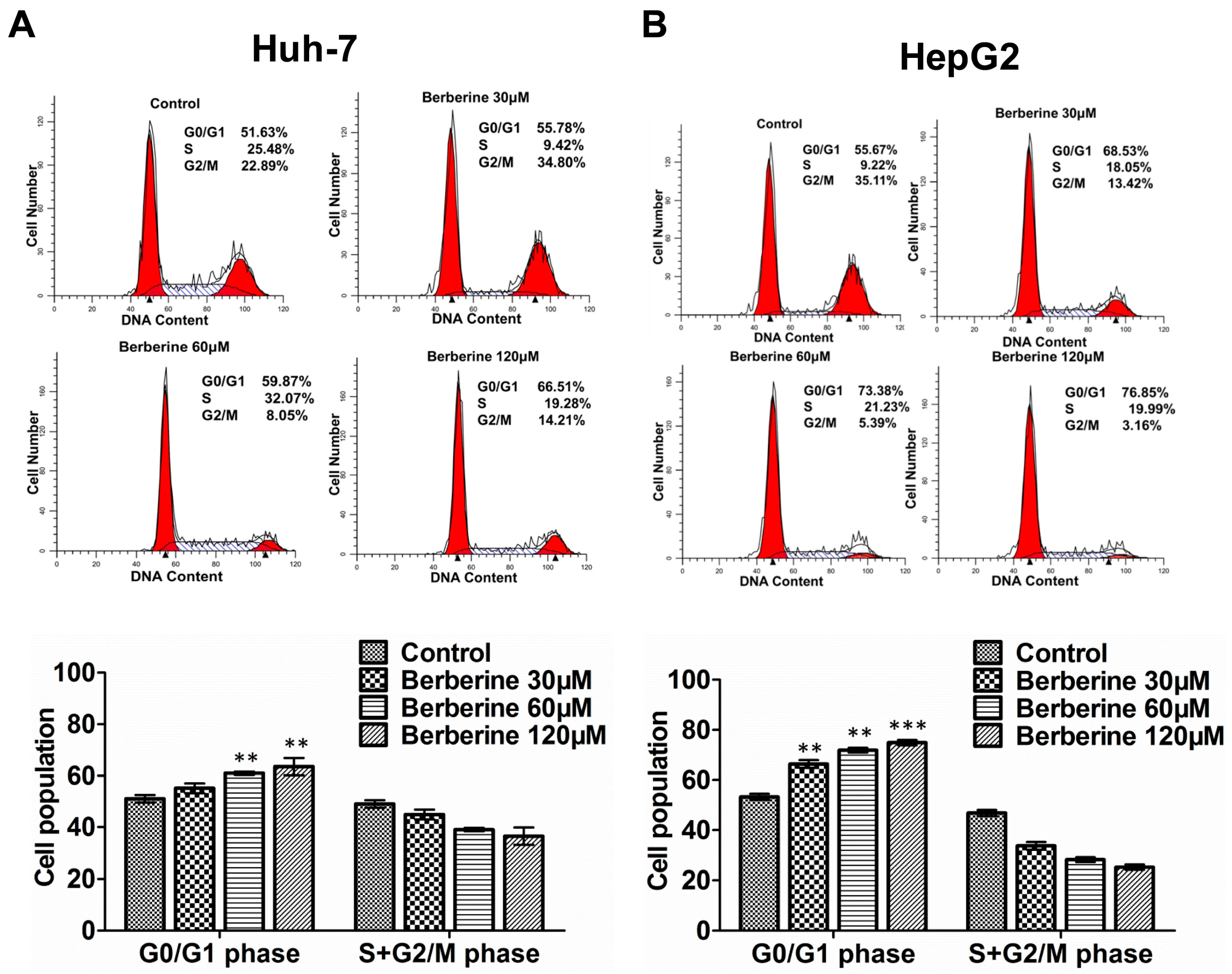
2.3. BBR Causes G0/G1 Phase Cell Cycle Arrest in Huh-7 and HepG2 Cells
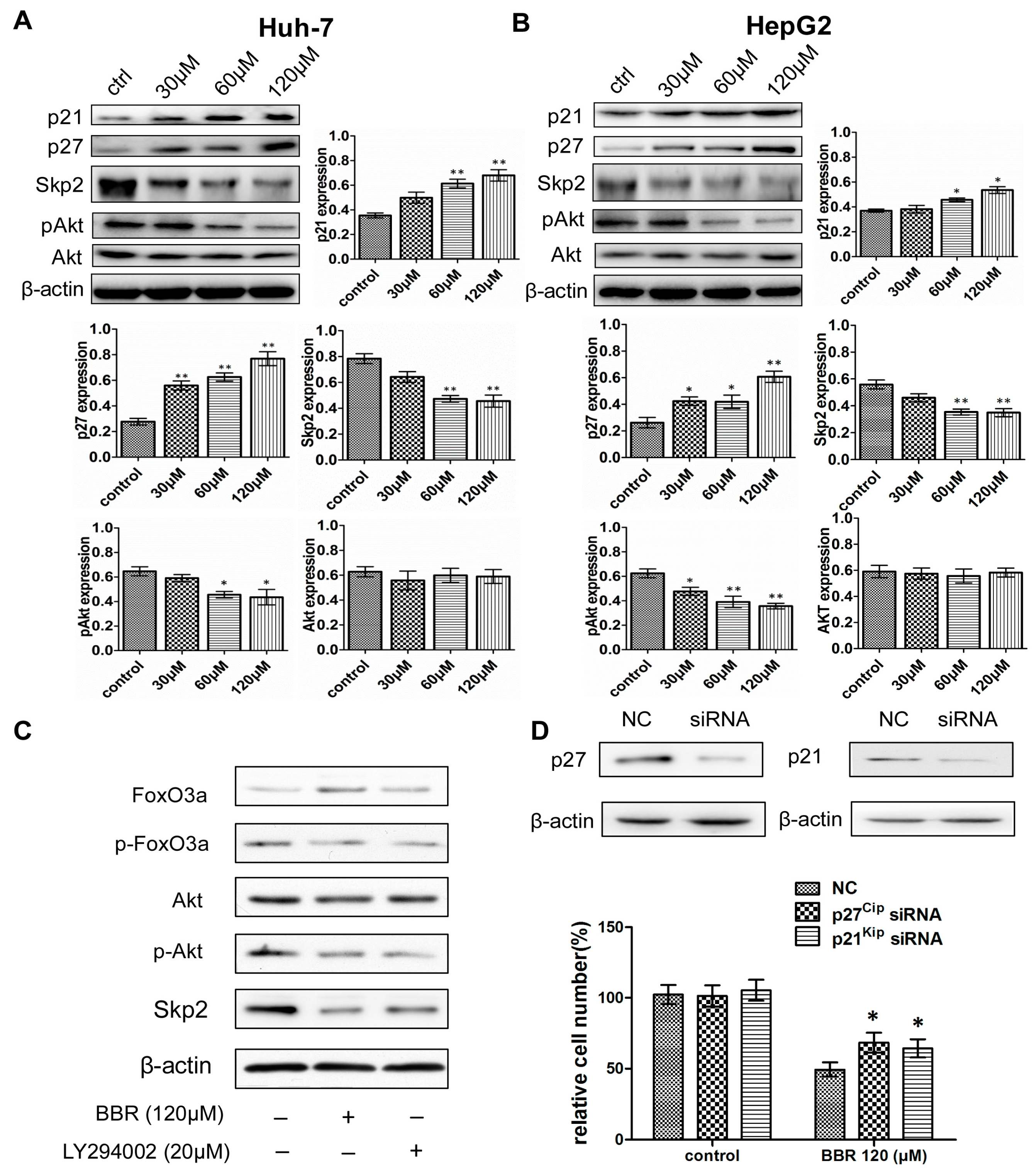
2.4. The Induction of BBR-Mediated G0/G1 Phase Cell Cycle Arrest Is Associated with the Regulation of p21Cip1, p27Kip1 and Skp2 Expression
2.5. Activation of Akt Is Involved in BBR-Mediated Skp2 Regulation
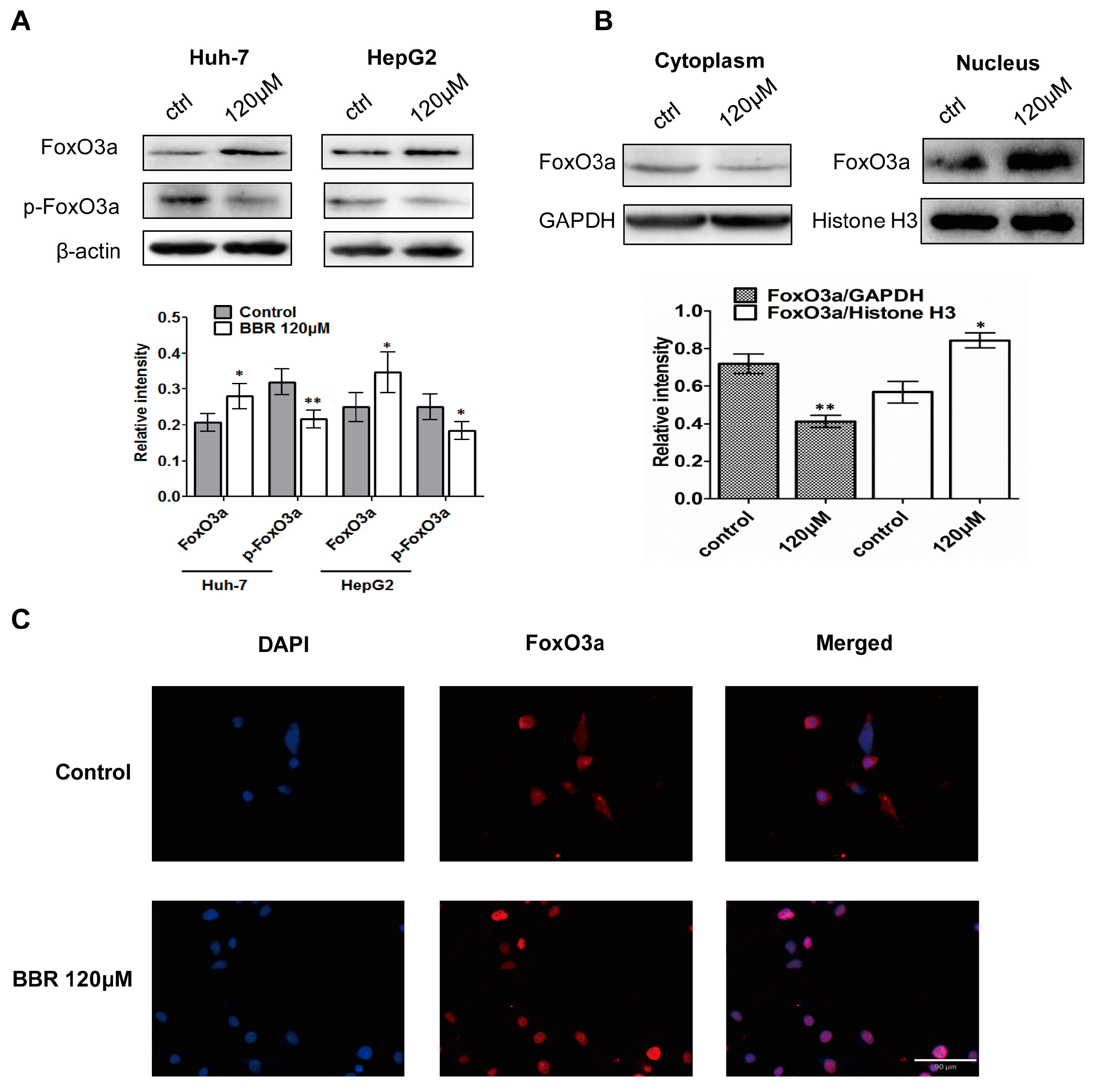
2.6. FoxO3a Translocates into Nucleus after BBR Treatment
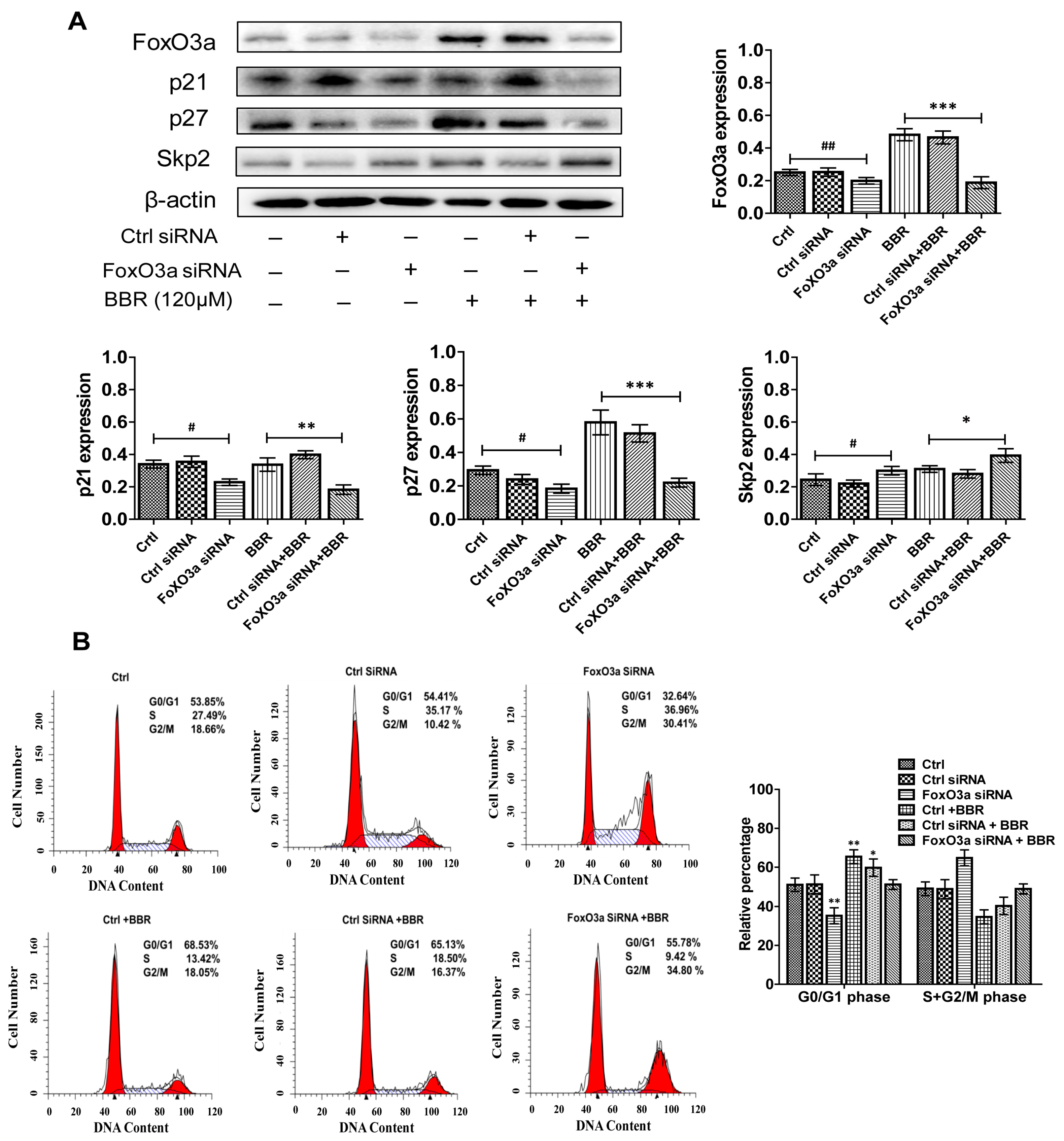
2.7. BBR-Induced G0/G1 Phase Cell Cycle Arrest Is Dependent on FoxO3a Expression
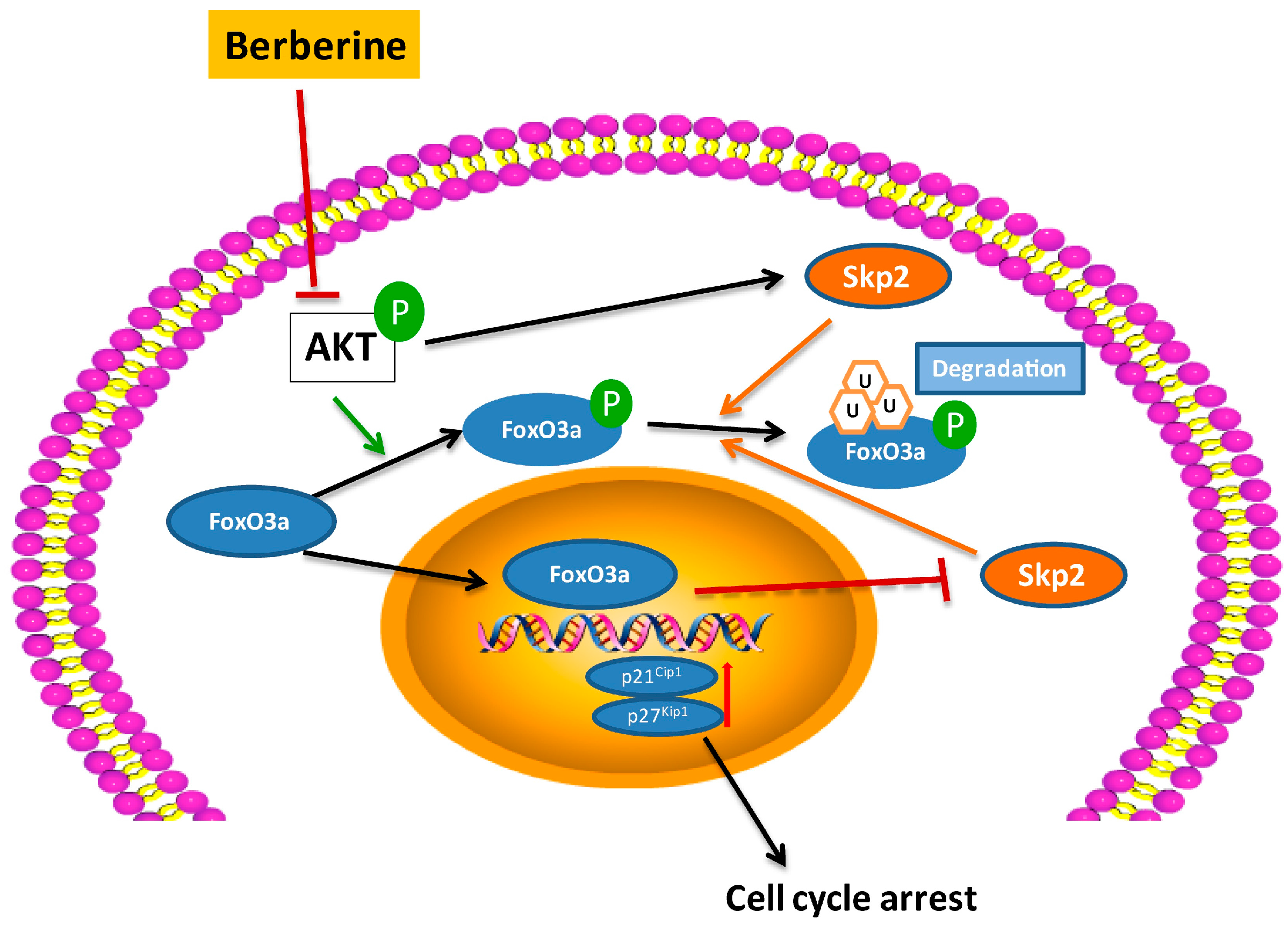
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture
4.3. Colony Formation Analysis
4.4. Cell Viability Analysis
4.5. Cell Cycle Analysis
4.6. Phosphatidylserine Externalization Analysis
4.7. Immunoflurescence Staining
4.8. Western Blot
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- El-Serag, H.B. Hepatocellular carcinoma. N. Engl. J. Med. 2011, 365, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Maluccio, M.; Covey, A. Recent progress in understanding, diagnosing, and treating hepatocellular carcinoma. CA Cancer J. Clin. 2012, 62, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Rizvi, F.; Raisuddin, S.; Kakkar, P. FoxO proteins’ nuclear retention and BH3-only protein Bim induction evoke mitochondrial dysfunction-mediated apoptosis in berberine-treated HepG2 cells. Free Radic. Biol. Med. 2014, 76, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Shi, Y.; Wang, X.; Zhou, J.; Qiu, S.; Fan, J. Translational medicine in hepatocellular carcinoma. Front. Med. 2012, 6, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, N.; Cheung, F.; Lao, L.; Li, C.; Feng, Y. Chinese medicines for prevention and treatment of human hepatocellular carcinoma: Current progress on pharmacological actions and mechanisms. J. Integr. Med. 2015, 13, 142–164. [Google Scholar] [CrossRef]
- Zou, K.; Li, Z.; Zhang, Y.; Zhang, H.Y.; Li, B.; Zhu, W.L.; Shi, J.Y.; Jia, Q.; Li, Y.M. Advances in the study of berberine and its derivatives: A focus on anti-inflammatory and anti-tumor effects in the digestive system. Acta Pharmacol. Sin. 2017, 38, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Chang, W. Non-coding RNAs and Berberine: A new mechanism of its anti-diabetic activities. Eur. J. Pharmacol. 2017, 795, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, Y.; Liu, K.; Zhao, Y.; Xu, B.; Xu, L.; Tan, L.; Tian, Y.; Li, C.; Zhang, W.; et al. Berberine inhibits colitis-associated tumorigenesis via suppressing inflammatory responses and the consequent EGFR signaling-involved tumor cell growth. Lab. Investig. 2017, 97, 1343–1353. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ekavali; Chopra, K.; Mukherjee, M.; Pottabathini, R.; Dhull, D.K. Current knowledge and pharmacological profile of berberine: An update. Eur. J. Pharmacol. 2015, 761, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Tang, X.; Liu, H.; Tang, J.; Yang, Y.; Jing, X.; Xiao, Q.; Wang, W.; Gou, X.; Wang, Z. Berberine induces cell death in human hepatoma cells in vitro by downregulating CD147. Cancer Sci. 2011, 102, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.M.; Cheung, K.C.; Cheung, Y.C.; Man, K.; Lui, V.W.; Tsao, S.W.; Feng, Y. Berberine suppresses Id-1 expression and inhibits the growth and development of lung metastases in hepatocellular carcinoma. Biochim. Biophys. Acta 2015, 1852, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.M.; Hyun, M.S.; Lim, S.Y.; Lee, W.Y.; Kim, D. The combination of berberine and irradiation enhances anti-cancer effects via activation of p38 MAPK pathway and ROS generation in human hepatoma cells. J. Cell. Biochem. 2009, 107, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, N.; Li, H.; Liu, M.; Cao, F.; Yu, X.; Zhang, J.; Tan, Y.; Xiang, L.; Feng, Y. Up-Regulation of PAI-1 and Down-Regulation of uPA Are Involved in Suppression of Invasiveness and Motility of Hepatocellular Carcinoma Cells by a Natural Compound Berberine. Int. J. Mol. Sci. 2016, 17, 577. [Google Scholar] [PubMed]
- Calnan, D.R.; Brunet, A. The FoxO code. Oncogene 2008, 27, 2276–2288. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.; Ma, X.; Li, H.Z.; Gao, Y.; Li, X.T.; Zhang, Y.; Ai, Q.; Zhang, P.; Song, E.L.; Huang, Q.B.; et al. Downregulation of FOXO3a promotes tumor metastasis and is associated with metastasis-free survival of patients with clear cell renal cell carcinoma. Clin. Cancer Res. 2014, 20, 1779–1790. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, L.; Shen, A.; Zhang, J.; Wang, Y.; Zhao, Y.; Zou, L.; Ke, Q.; He, F.; Wang, P.; et al. Clinical and biological significance of forkhead class box O 3a expression in glioma: Mediation of glioma malignancy by transcriptional regulation of p27kip1. J. Neuro-Oncol. 2010, 98, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.X.; Liu, X.M.; Wang, J.; Li, J.; Liu, Y.; Zhang, H.; Yu, X.W.; Wei, N. Inhibition of AKT/FoxO3a signaling induced PUMA expression in response to p53-independent cytotoxic effects of H1: A derivative of tetrandrine. Cancer Biol. Ther. 2015, 16, 965–975. [Google Scholar] [PubMed]
- Jia, T.; Zhang, L.; Duan, Y.; Zhang, M.; Wang, G.; Zhang, J.; Zhao, Z. The differential susceptibilities of MCF-7 and MDA-MB-231 cells to the cytotoxic effects of curcumin are associated with the PI3K/Akt-SKP2-Cip/Kips pathway. Cancer Cell Int. 2014, 14, 126. [Google Scholar] [CrossRef] [PubMed]
- Frescas, D.; Pagano, M. Deregulated proteolysis by the F-box proteins SKP2 and β-TrCP: Tipping the scales of cancer. Nat. Rev. Cancer 2008, 8, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Koo, K.H.; Kim, H.; Bae, Y.K.; Kim, K.; Park, B.K.; Lee, C.H.; Kim, Y.N. Salinomycin induces cell death via inactivation of Stat3 and downregulation of Skp2. Cell Death Dis. 2013, 4, e693. [Google Scholar] [CrossRef] [PubMed]
- Carrano, A.C.; Eytan, E.; Hershko, A.; Pagano, M. SKP2 is required for ubiquitin-mediated degradation of the CDK inhibitor p27. Nat. Cell Biol. 1999, 1, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tong, S.; Tai, A.W.; Hussain, M.; Lok, A.S. Hepatitis B virus core promoter mutations contribute to hepatocarcinogenesis by deregulating SKP2 and its target, p21. Gastroenterology 2011, 141, 1412–1421.e5. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Ninomiya, K.; Hamada, H.; Noda, M. Involvement of the SKP2-p27(KIP1) pathway in suppression of cancer cell proliferation by RECK. Oncogene 2012, 31, 4128–4138. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.K.; Wang, G.; Chen, Z.; Teruya-Feldstein, J.; Liu, Y.; Chan, C.H.; Yang, W.L.; Erdjument-Bromage, H.; Nakayama, K.I.; Nimer, S.; et al. Phosphorylation-dependent regulation of cytosolic localization and oncogenic function of Skp2 by Akt/PKB. Nat. Cell Biol. 2009, 11, 420–432. [Google Scholar]
- Yi, T.; Zhuang, L.; Song, G.; Zhang, B.; Li, G.; Hu, T. Akt signaling is associated with the berberine-induced apoptosis of human gastric cancer cells. Nutr. Cancer 2015, 67, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Lee, S.W.; Zhang, X.; Han, F.; Kwan, S.Y.; Yuan, X.; Yang, W.L.; Jeong, Y.S.; Rezaeian, A.H.; Gao, Y.; et al. Foxo3a transcription factor is a negative regulator of Skp2 and Skp2 SCF complex. Oncogene 2013, 32, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, M.; Khare, V.; Guturi, K.K.; Das, N.; Ghosh, M.K. The DEAD box protein p68: A crucial regulator of AKT/FOXO3a signaling axis in oncogenesis. Oncogene 2015, 34, 5843–5856. [Google Scholar] [CrossRef] [PubMed]
- Prindull, G. Final checkup of neoplastic DNA replication: Evidence for failure in decision-making at the mitotic cell cycle checkpoint G1/S. Exp. Hematol. 2008, 36, 1403–1416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bai, D.S.; Huang, X.Y.; Shi, G.M.; Ke, A.W.; Yang, L.X.; Yang, X.R.; Zhou, J.; Fan, J. Prognostic significance of Capn4 overexpression in intrahepatic cholangiocarcinoma. PLoS ONE 2013, 8, e54619. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Y.; Lv, Q.H.; Dong, Q.; Xu, R.Z.; Dong, Q.H. Berbamine induces Fas-mediated apoptosis in human hepatocellular carcinoma HepG2 cells and inhibits its tumor growth in nude mice. J. Asian Nat. Prod. Res. 2009, 11, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Piao, Y.; Thomas, C.; Fuller, G.N.; de Groot, J.F. Cdc2-like kinase 2 is a key regulator of the cell cycle via FOXO3a/p27 in glioblastoma. Oncotarget 2016, 7, 26793–26805. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wang, J.; Wu, M.; Zhang, X.; Zhang, H. The inhibition of activated hepatic stellate cells proliferation by arctigenin through G0/G1 phase cell cycle arrest: Persistent p27(Kip1) induction by interfering with PI3K/Akt/FOXO3a signaling pathway. Eur. J. Pharmacol. 2015, 747, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Dobson, M.; Ramakrishnan, G.; Ma, S.; Kaplun, L.; Balan, V.; Fridman, R.; Tzivion, G. Bimodal regulation of FoxO3 by AKT and 14-3-3. Biochim. Biophys. Acta 2011, 1813, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, V.V.; Allen, J.E.; Dicker, D.T.; El-Deiry, W.S. Small-Molecule ONC201/TIC10 Targets Chemotherapy-Resistant Colorectal Cancer Stem-like Cells in an Akt/Foxo3a/TRAIL-Dependent Manner. Cancer Res. 2015, 75, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.B.; Chung, Y.M.; Takahashi, Y.; Xu, Z.; Hu, M.C. Functional interaction between FOXO3a and ATM regulates DNA damage response. Nat. Cell Biol. 2008, 10, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Dong, F.; Luan, C.; Zhang, X.; Shao, H.; Liu, J.; Sun, C. Overexpression of CCAAT Enhancer-Binding Protein alpha Inhibits the Growth of K562 Cells via the Foxo3a-Bim Pathway. Acta Haematol. 2016, 136, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Ferber, E.C.; Peck, B.; Delpuech, O.; Bell, G.P.; East, P.; Schulze, A. FOXO3a regulates reactive oxygen metabolism by inhibiting mitochondrial gene expression. Cell Death Differ. 2012, 19, 968–979. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kobayashi, R.; Galaktionov, K.; Beach, D. p19Skp1 and p45Skp2 are essential elements of the cyclin A-CDK2 S phase kinase. Cell 1995, 82, 915–925. [Google Scholar] [CrossRef]
- Hao, Z.; Huang, S. E3 ubiquitin ligase Skp2 as an attractive target in cancer therapy. Front. Biosci. 2015, 20, 474–490. [Google Scholar] [CrossRef]
- Caraballo, J.M.; Acosta, J.C.; Cortes, M.A.; Albajar, M.; Gomez-Casares, M.T.; Batlle-Lopez, A.; Cuadrado, M.A.; Onaindia, A.; Bretones, G.; Llorca, J.; et al. High p27 protein levels in chronic lymphocytic leukemia are associated to low Myc and Skp2 expression, confer resistance to apoptosis and antagonize Myc effects on cell cycle. Oncotarget 2014, 5, 4694–4708. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chan, C.H.; Chen, K.; Guan, X.; Lin, H.K.; Tong, Q. Deacetylation of FOXO3 by SIRT1 or SIRT2 leads to Skp2-mediated FOXO3 ubiquitination and degradation. Oncogene 2012, 31, 1546–1557. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Dong, X.; Lin, P.; Jiang, J. Regulation of Akt/FoxO3a/Skp2 Axis Is Critically Involved in Berberine-Induced Cell Cycle Arrest in Hepatocellular Carcinoma Cells. Int. J. Mol. Sci. 2018, 19, 327. https://doi.org/10.3390/ijms19020327
Li F, Dong X, Lin P, Jiang J. Regulation of Akt/FoxO3a/Skp2 Axis Is Critically Involved in Berberine-Induced Cell Cycle Arrest in Hepatocellular Carcinoma Cells. International Journal of Molecular Sciences. 2018; 19(2):327. https://doi.org/10.3390/ijms19020327
Chicago/Turabian StyleLi, Fanni, Xiwen Dong, Peng Lin, and Jianli Jiang. 2018. "Regulation of Akt/FoxO3a/Skp2 Axis Is Critically Involved in Berberine-Induced Cell Cycle Arrest in Hepatocellular Carcinoma Cells" International Journal of Molecular Sciences 19, no. 2: 327. https://doi.org/10.3390/ijms19020327
APA StyleLi, F., Dong, X., Lin, P., & Jiang, J. (2018). Regulation of Akt/FoxO3a/Skp2 Axis Is Critically Involved in Berberine-Induced Cell Cycle Arrest in Hepatocellular Carcinoma Cells. International Journal of Molecular Sciences, 19(2), 327. https://doi.org/10.3390/ijms19020327