The Janus Face of NKT Cell Function in Autoimmunity and Infectious Diseases



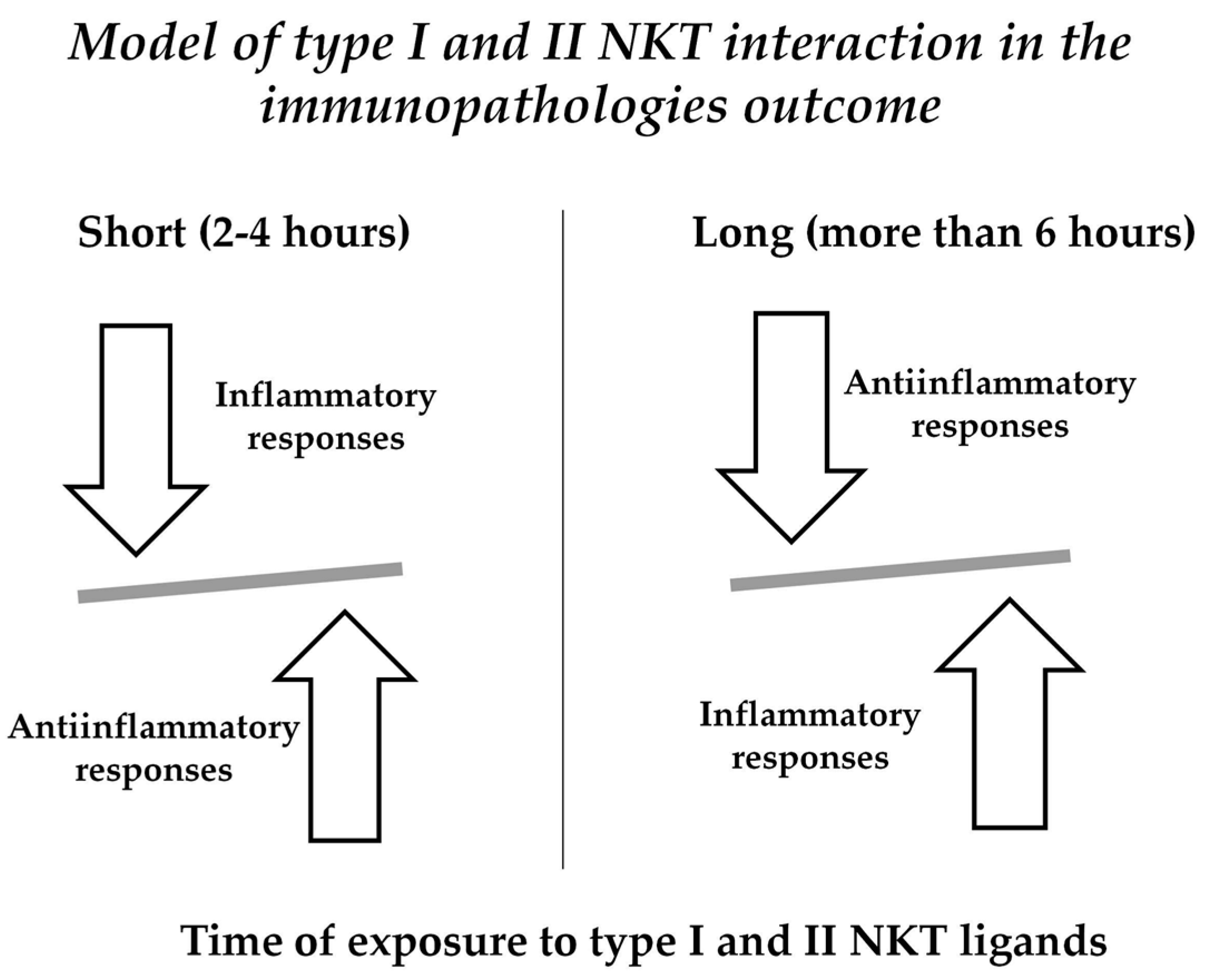{kind=link}
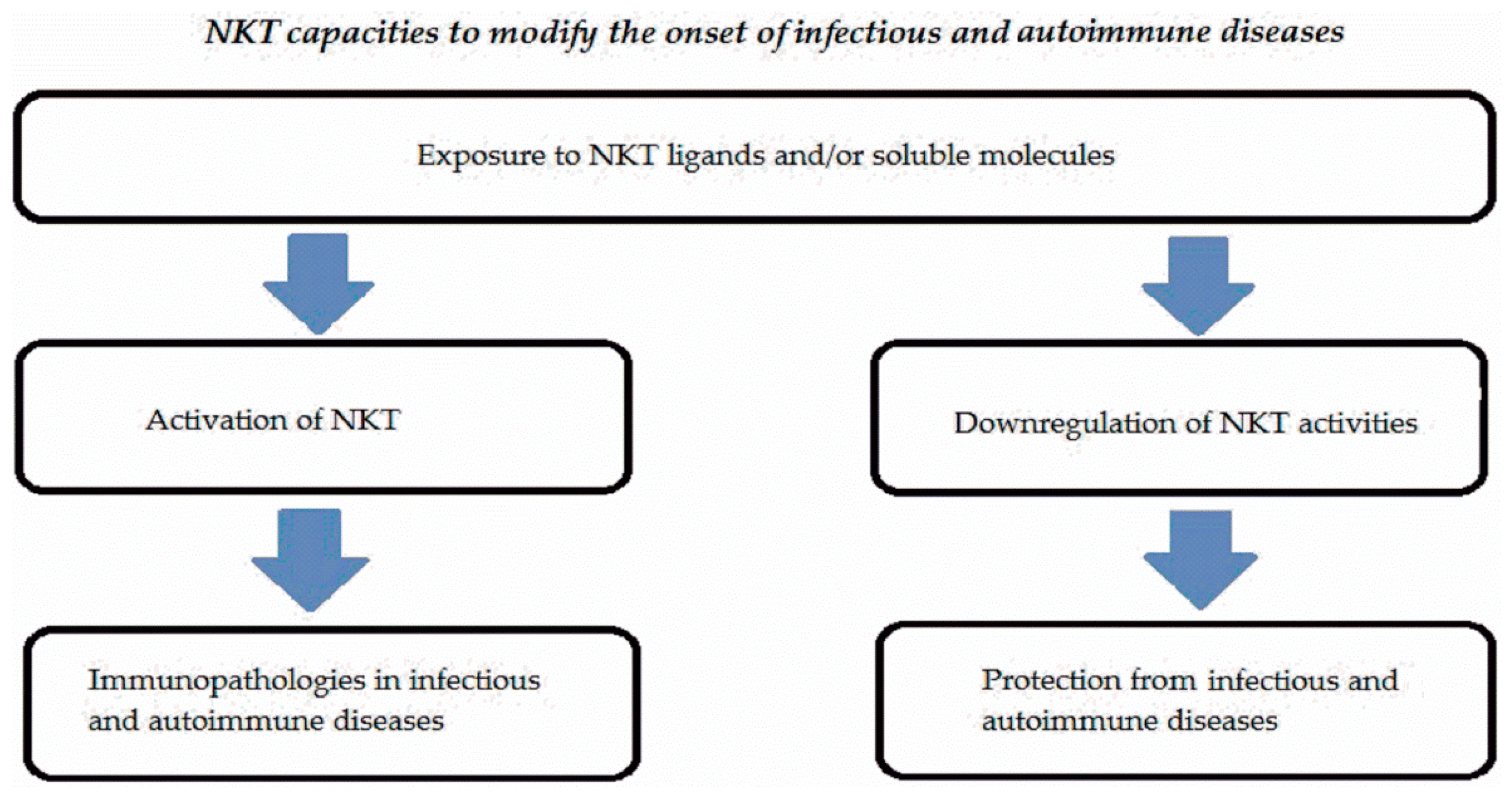{kind=link}
Abstract
:1. Distinctive Functional Activities of Types of NKT
2. Type I NKT in Response to Microbial Antigens
3. Role of Type II NKT in Immune Responses to Different Microorganisms
4. Type I NKT in Autoimmune and Chronic Inflammatory Diseases
5. Type II NKT in Autoimmune and Chronic Inflammatory Diseases
6. CIK Cells as Players of Antimicrobial Immune Response
7. Concluding Remarks
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Moody, D.B.; Sugita, M.; Peters, P.J.; Brenner, M.B.; Porcelli, S.A. The CD1-restricted T-cell response to mycobacteria. Res. Immunol. 1996, 147, 550–559. [Google Scholar] [CrossRef]
- Moody, D.B.; Reinhold, B.B.; Guy, M.R.; Beckman, E.M.; Frederique, D.E.; Furlong, S.T.; Ye, S.; Reinhold, V.N.; Sieling, P.A.; Modlin, R.L.; et al. Structural requirements for glycolipid antigen recognition by CD1b-restricted T cells. Science 1997, 278, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Rosat, J.P.; Grant, E.P.; Beckman, E.M.; Dascher, C.C.; Sieling, P.A.; Frederique, D.; Modlin, R.L.; Porcelli, S.A.; Furlong, S.T.; Brenner, M.B. CD1-restricted microbial lipid antigen-specific recognition found in the CD8+ αβ T cell pool. J. Immunol. 1999, 162, 366–371. [Google Scholar] [PubMed]
- Shamshiev, A.; Donda, A.; Carena, I.; Mori, L.; Kappos, L.; De Libero, G. Self-glycolipids as T-cell autoantigens. Eur. J. Immunol. 1999, 29, 1667–1675. [Google Scholar] [CrossRef]
- Moody, D.B.; Guy, M.R.; Grant, E.; Cheng, T.Y.; Brenner, M.B.; Besra, G.S.; Porcelli, S.A. CD1b-mediated T cell recognition of a glycolipid antigen generated from mycobacterial lipid and host carbohydrate during infection. J. Exp. Med. 2000, 192, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Moody, D.B.; Ulrichs, T.; Muhlecker, W.; Young, D.C.; Gurcha, S.S.; Grant, E.; Rosat, J.P.; Brenner, M.B.; Costello, C.E.; Besra, G.S.; et al. CD1c-mediated T-cell recognition of isoprenoid glycolipids in Mycobacterium tuberculosis infection. Nature 2000, 404, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Moody, D.B.; Young, D.C.; Cheng, T.Y.; Rosat, J.P.; Roura-Mir, C.; O’Connor, P.B.; Zajonc, D.M.; Walz, A.; Miller, M.J.; Levery, S.B.; et al. T cell activation by lipopeptide antigens. Science 2004, 303, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Birkinshaw, R.W.; Pellicci, D.G.; Cheng, T.Y.; Keller, A.N.; Sandoval-Romero, M.; Gras, S.; de Jong, A.; Uldrich, A.P.; Moody, D.B.; Godfrey, D.I.; et al. αβ T cell antigen receptor recognition of CD1a presenting self lipid ligands. Nat. Immunol. 2015, 16, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Arrenberg, P.; Halder, R.; Dai, Y.; Maricic, I.; Kumar, V. Oligoclonality and innate-like features in the TCR repertoire of type II NKT cells reactive to a beta-linked self-glycolipid. Proc. Natl. Acad. Sci. USA 2010, 107, 10984–10989. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, J.L.; Gapin, L.; Baron, J.L.; Sidobre, S.; Stetson, D.B.; Mohrs, M.; Locksley, R.M.; Kronenberg, M. Mouse V alpha 14i natural killer cells are resistant to cytokine polarization in vivo. Proc. Natl. Acad. Sci. USA 2003, 100, 8395–8400. [Google Scholar] [CrossRef] [PubMed]
- Sag, D.; Krause, P.; Hedrick, C.C.; Kronenberg, M.; Wingender, G. IL-10-producing NKT10 cells are a distinct regulatory invariant NKT cell subset. J. Clin. Investig. 2014, 124, 3725–3740. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Shin, T.; Kawano, T.; Sato, H.; Kondo, E.; Toura, I.; Kaneko, Y.; Koseki, H.; Kanno, M.; Taniguchi, M. Requirement for Vα14 NKT cells in IL-12-mediated rejection of tumors. Science 1997, 278, 1623–1626. [Google Scholar] [CrossRef] [PubMed]
- Jahng, A.; Maricic, I.; Aguilera, C.; Cardell, S.; Halder, R.C.; Kumar, V. Prevention of autoimmunity by targeting a distinct, noninvariant CD1d-reactive T cell population reactive to sulfatide. J. Exp. Med. 2004, 199, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Hodge, G.; Hodge, S. Steroid resistant CD8+CD28null NKT-like pro-inflammatory cytotoxic cells in chronic obstructive pulmonary disease. Front. Immunol. 2016, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Wolf, I.G.; Negrin, R.S.; Kiem, H.P.; Blume, K.G.; Weissman, I.L. Use of a SCID mouse/human lymphoma model to evaluate cytokine-induced killer cells with potent antitumor cell activity. J. Exp. Med. 1991, 174, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, D.I.; Kronenberg, M. Going both ways: Immune regulation via CD1d-dependent NKT cells. J. Clin. Investig. 2004, 114, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg, M.; Gapin, L. The unconventional lifestyle of NKT cells. Nat. Rev. Immunol. 2002, 2, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Tupin, E.; Kinjo, Y.; Kronenberg, M. The unique role of natural killer T cells in the response to microorganisms. Nat. Rev. Microbiol. 2007, 5, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Skold, M.; Behar, S.M. Role of CD1d-restricted NKT cells in microbial immunity. Infect. Immun. 2003, 71, 5447–5455. [Google Scholar] [CrossRef] [PubMed]
- Zajonc, M.D.; Girardi, E. Recognition of microbial glycolipids by natural killer T cells. Front. Immunol. 2015, 400, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Leite-De-Moraes, M.C.; Hameg, A.; Arnould, A.; Machavoine, F.; Koezuka, Y.; Schneider, E.; Herbelin, A.; Dy, M. A distinct IL-18-induced pathway to fully activate NKT lymphocytes independently from TCR engagement. J. Immunol. 1999, 163, 5871–5876. [Google Scholar] [PubMed]
- Nagarajan, N.A.; Kronenberg, M. Invariant NKT cells amplify the innate immune response to lipopolysaccharide. J. Immunol. 2007, 178, 2706–2713. [Google Scholar] [CrossRef] [PubMed]
- Wesley, J.D.; Tessmer, M.S.; Chaukos, D.; Brossay, L. NK cell-like behavior of Vα14i NK T cells during MCMV infection. PLoS Pathog. 2008, 4, e1000106. [Google Scholar] [CrossRef] [PubMed]
- Brigl, M.; Bry, L.; Kent, S.C.; Gumperz, J.E.; Brenner, M.B. Mechanism of CD1d restricted natural killer T cell activation during microbial infection. Nat. Immunol. 2003, 4, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Paget, C.; Mallevaey, T.; Speak, A.O.; Torres, D.; Fontaine, J.; Sheehan, K.C.; Capron, M.; Ryffel, B.; Faveeuw, C.; Leite de Moraes, M.; et al. Activation of invariant NKT cells by toll-like receptor 9-stimulated dendritic cells requires type I interferon and charged glycosphingolipids. Immunity 2007, 27, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Brigl, M.; Tatituri, R.V.; Watts, G.F.; Bhowruth, V.; Leadbetter, E.A.; Barton, N.; Cohen, N.R.; Hsu, F.F.; Besra, G.S.; Brenner, M.B. Innate and cytokine-driven signals, rather than microbial antigens, dominate in natural killer T cell activation during microbial infection. J. Exp. Med. 2011, 208, 1163–1177. [Google Scholar] [CrossRef] [PubMed]
- Tyznik, A.J.; Tupin, E.; Nagarajan, N.A.; Her, M.J.; Benedict, C.A.; Kronenberg, M. Cutting edge: The mechanism of invariant NKT cell responses to viral danger signals. J. Immunol. 2008, 181, 4452–4456. [Google Scholar] [CrossRef] [PubMed]
- Olson, C.M., Jr.; Bates, T.C.; Izadi, H.; Radolf, J.D.; Huber, S.A.; Boyson, J.E.; Anguita, J. Local production of IFN-gamma by invariant NKT cells modulates acute Lyme carditis. J. Immunol. 2009, 182, 3728–3734. [Google Scholar] [CrossRef] [PubMed]
- Kinjo, Y.; Tupin, E.; Wu, D.; Fujio, M.; Garcia-Navarro, R.; Benhnia, M.R.; Zajonc, D.M.; Ben-Menachem, G.; Ainge, G.D.; Painter, G.F.; et al. Natural killer T cells recognize diacylglycerol antigens from pathogenic bacteria. Nat. Immunol. 2006, 7, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Ben-Menachem, G.; Kubler-Kielb, J.; Coxon, B.; Yergey, A.; Schneerson, R. A newly discovered cholesteryl galactoside from Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2003, 100, 7913–7918. [Google Scholar] [CrossRef] [PubMed]
- Schröder, N.W.; Schombel, U.; Heine, H.; Gobel, U.B.; Zähringer, U.; Schumann, R.R. Acylated cholesteryl galactoside as a novel immunogenic motif in Borrelia burgdorferi sensu stricto. J. Biol. Chem. 2003, 278, 33645–33653. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Vela, J.L.; Matsumura, F.; Hoshino, H.; Tyznik, A.; Lee, H.; Girardi, E.; Zajonc, D.M.; Liddington, R.; Kobayashi, M.; et al. Helicobacter pylori cholesteryl alpha-glucosides contribute to its pathogenicity and immune response by natural killer T cells. PLoS ONE 2013, 8, e78191. [Google Scholar] [CrossRef] [PubMed]
- Lotter, H.; Gonzalez-Roldan, N.; Lindner, B.; Winau, F.; Isibasi, A.; Moreno-Lafont, M.; Ulmer, A.J.; Holst, O.; Tannich, E.; Jacobs, T. Natural killer T cells activated by a lipopeptidophosphoglycan from Entamoeba histolytica are critically important to control amebic liver abscess. PLoS Pathog. 2009, 5, e1000434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Manna, M.P.; Torina, A.; Agnone, A.; Blanda, V.; Caracappa, S.; Alongi, A.; Di Marco, V.; Giudice, E.; Dieli, F.; Sireci, G. Detection of natural killer T cells in mice infected with Rickettsia conorii. Transbound. Emerg. Dis. 2013, 60 (Suppl. 2), 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keino, H.; Watanabe, T.; Taki, W.; Okada, A.A. Effect of Infliximab on gene expression profiling in Behçet’s disease. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7681–7686. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Chimal, J.; Fernández-Figueroa, E.A.; Ruiz-Remigio, A.; Wilkins-Rodríguez, A.A.; Delgado-Domínguez, J.; Salaiza-Suazo, N.; Gutiérrez-Kobeh, L.; Becker, I. NKT cell activation by Leishmania mexicana LPG: Description of a novel pathway. Immunobiol 2017, 222, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Amprey, J.L.; Im, J.S.; Turco, S.J.; Murray, H.W.; Illarionov, P.A.; Besra, G.S.; Porcelli, S.A.; Späth, G.F. A subset of liver NK T cells is activated during Leishmania donovani infection by CD1d-bound lipophosphoglycan. J. Exp. Med. 2004, 200, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Albacker, L.A.; Chaudhary, V.; Chang, Y.-J.; Kim, H.Y.; Chuang, Y.T.; Pichavant, M.; DeKruyff, R.H.; Savage, P.B.; Umetsu, D.T. A Fungal Glycosphingolipid Directly Activates Natural Killer T Cells and Rapidly Induces Airways Disease. Nat. Med. 2013, 19, 1297–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duthie, M.S.; Kahn, M.; White, M.; Kapur, R.P.; Kahn, S.J. Critical proinflammatory and anti-inflammatory functions of different subsets of CD1d-restricted natural killer T cells during Trypanosoma cruzi infection. Infect. Immun. 2005, 73, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Mallevaey, T.; Zanetta, J.P.; Faveeuw, C.; Fontaine, J.; Maes, E.; Platt, F.; Capron, M.; de-Moraes, M.L.; Trottein, F. Activation of invariant NKT cells by the helminth parasite Schistosoma mansoni. J. Immunol. 2006, 176, 2476–2485. [Google Scholar] [CrossRef] [PubMed]
- Kwiecinski, J.; Rhost, S.; Lofbom, L.; Blomqvist, M.; Mansson, J.E.; Cardell, S.L.; Jin, T. Sulphatide attenuates experimental Staphylococcus aureus sepsis through a CD1d dependent pathway. Infect. Immun. 2013, 81, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Tatituri, R.V.; Watts, G.F.; Bhowruth, V.; Barton, N.; Rothchild, A.; Hsu, F.F.; Almeida, C.F.; Cox, L.R.; Eggeling, L.; Cardell, S.; et al. Recognition of microbial and mammalian phospholipid antigens by NKT cells with diverse TCRs. Proc. Natl. Acad. Sci. USA 2013, 110, 1827–1832. [Google Scholar] [CrossRef] [PubMed]
- Wolf, B.J.; Tatituri, R.V.; Almeida, C.F.; Le Nours, J.; Bhowruth, V.; Johnson, D.; Uldrich, A.P.; Hsu, F.F.; Brigl, M.; Besra, G.S.; et al. Identification of a potent microbial lipid antigen for diverse NKT cells. J. Immunol. 2015, 195, 2540–2551. [Google Scholar] [CrossRef] [PubMed]
- Zeissig, S.; Murata, K.; Sweet, L.; Publicover, J.; Hu, Z.; Kaser, A.; Bosse, E.; Iqbal, J.; Hussain, M.M.; Balschun, K.; et al. Hepatitis B virus-induced lipid alterations contribute to natural killer T cell-dependent protective immunity. Nat. Med. 2012, 18, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, C.S.; Kelleher, A.D.; Finlayson, R.; Godfrey, D.I.; Kent, S.J. NKT cell depletion in humans during early HIV infection. Immunol. Cell Biol. 2014, 92, 578–590. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.N.; Kee, S.-J.; Lee, S.-J.; Seo, S.-J.; Kim, T.-J.; Lee, S.-S.; Kim, M.-S.; Lee, W.-W.; Yoo, D.-H.; Kim, N.; et al. Numerical and functional deficiencies of natural killer T cells in systemic lupus erythematosus: Their deficiency related to disease activity. Rheumatology 2011, 50, 1054–1063. [Google Scholar] [CrossRef] [PubMed]
- Gutowska-Owsiak, D.; Birchall, M.A.; Moots, R.J.; Christmas, S.E.; Pazmany, L. Proliferatory defect of invariant population and accumulation of non-invariant CD1d-restricted natural killer T cells in the joints of RA patients. Mod. Rheumatol. 2014, 24, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Van der Vliet, B.M.; von Blomberg, E.; Nishi, N.; Reijm, M.; Voskuyl, A.E.; van Bodegraven, A.A.; Polman, C.H.; Rustemeyer, T.; Lips, P.; van den Eertwegh, A.J.; et al. Circulating Vα24+ Vβ11+ NKT cell numbers are decreased in a wide variety of diseases that are characterized by autoreactive tissue damage. Clin. Immunol. 2001, 100, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Mieza, M.A.; Itoh, T.; Cui, J.Q.; Makino, Y.; Kawano, T.; Tsuchida, K.; Koike, T.; Shirai, T.; Yagita, H.; Matsuzawa, A.; et al. Selective reduction of Vα14+NKTcells associated with disease development in autoimmune-prone mice. J. Immunol. 1996, 156, 4035–4040. [Google Scholar] [PubMed]
- Kojo, S.; Adachi, Y.; Keino, H.; Taniguchi, M.; Sumida, T. Dysfunction of T cell receptor AV24 AJ18+, BV11+ double negative regulatory natural killer T cells in autoimmune diseases. Arthritis Rheum. 2001, 44, 1127–1138. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, Y.; Yang, Q.; Hou, Y.; Hu, N.; Wang, D.; Sun, H. The aberrant expression of stimulatory and inhibitory killer immunoglobulin-like receptors in NK- and NKT-cells contributes to lupus. Clin. Lab. 2014, 60, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.T.Y.; Wong, C.K.; Tam, L.S.; Li, E.K.; Chen, D.P.; Lam, C.W.K. Decreased expression of T lymphocyte co-stimulatory molecule CD26 on invariant natural killer T cells in systemic lupus erythematosus. Immunol. Investig. 2009, 38, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhang, B.; Jarrell, J.A.; Price, J.V.; Dai, H.; Utz, P.J.; Strober, S. Ly108 expression distinguishes subsets of invariant NKT cells that help autoantibody production and secrete IL-21 from those that secrete IL-17 in lupus prone NZB/W mice. J. Autoimmun. 2014, 50, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Yoshiga, Y.; Goto, D.; Segawa, S.; Ohnishi, Y.; Matsumoto, I.; Ito, S.; Tsutsumi, A.; Taniguchi, M.; Sumida, T. Invariant NKT cells produce IL-17 through IL-23-dependent and -independent pathways with potential modulation of Th17 response in collagen induced arthritis. Int. J. Mol. Med. 2013, 31, 998. [Google Scholar] [CrossRef]
- Green, M.R.J.; Kennell, A.S.M.; Larche, M.J.; Seifert, M.H.; Isenberg, D.A.; Salaman, M.R. Natural killer T cells in families of patients with systemic lupus erythematosus: Their possible role in regulation of IgG production. Arthritis Rheum. 2007, 56, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Wermeling, F.; Lind, S.M.; Jordö, E.D.; Cardell, S.L.; Karlsson, M.C.I. Invariant NKT cells limit activation of autoreactive CD1d-positive B cells. J. Exp. Med. 2010, 207, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Q.; Kim, P.J.; Singh, R.R. Brief treatment with iNKT cell ligand α-galactosylceramide confers a long-term protection against lupus. J. Clin. Immunol. 2012, 32, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Tohn, R.; Blumenfeld, H.; Haeryfar, S.M.M.; Veerapen, N.; Besra, G.S.; Porcelli, S.A.; Delovitch, T.L. Stimulation of a shorter duration in the state of anergy of invariant natural killer T cell agonist enhances its efficiency of protection from type I diabetes. Clin. Exp. Immunol. 2011, 164, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Delovitch, T.L. Different subsets of natural killer T cells may vary in their roles in health and disease. Immunology 2014, 142, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, L.; Blumenfeld, H.; Tohn, R.; Ly, D.; Aguilera, C.; Maricic, I.; Mansson, J.E.; Buschard, K.; Kumar, V.; Delovitch, T.L. NKT cells stimulated by long fatty acyl chain sulphatides significantly reduce the incidence of type 1 diabetes in nonobese diabetic mice1. PLoS ONE 2012, 7, e37771. [Google Scholar] [CrossRef]
- Maricic, I.; Halder, R.; Bischof, F.; Kumar, V. Dendritic cells and anergic type I NKT cells play a crucial role in sulfatide-mediated immune regulation in experimental autoimmune encephalomyelitis. J. Immunol. 2014, 193, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Fuss, I.J.; Heller, F.; Boirivant, M.; Leon, F.; Yoshida, M.; Fichtner-Feigl, S.; Yang, Z.; Exley, M.; Kitani, A.; Blumberg, R.S.; et al. Nonclassical CD1d-restricted NKT cells that produce IL-13 characterize an atypical Th2 response in ulcerative colitis. J. Clin. Investig. 2004, 113, 1490–1497. [Google Scholar] [CrossRef] [PubMed]
- Fuss, I.J.; Joshi, B.; Yang, Z.; Degheidy, H.; Fichtner-Feigl, S.; de Souza, H.; Rieder, F.; Scaldaferri, F.; Schirbel, A.; Scarpa, M.; et al. IL-13Ralpha2-bearing, type II NKT cells reactive to sulfatide self-antigen populate the mucosa of ulcerative colitis. Gut 2014, 63, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.M.; Zimmer, M.I.; Shanmuganad, S.; Yu, H.T.; Cardell, S.L.; Wang, C.R. Dysregulation of CD1d-restricted type II natural killer T cells leads to spontaneous development of colitis in mice. Gastroenterology 2012, 142, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Pievani, A.; Borleri, G.; Pende, D.; Moretta, L.; Rambaldi, A.; Golay, J.; Introna, M. Dual-functional capability of CD3+CD56+ CIK cells, a T-cell subset that acquires NK function and retains TCR-mediated specific cytotoxicity. Blood 2011, 118, 3301–3310. [Google Scholar] [CrossRef] [PubMed]
- Mehta, B.A.; Schmidt-Wolf, I.G.; Weissman, I.L.; Negrin, R.S. Two pathways of exocytosis of cytoplasmic granule contents and target cell killing by cytokine-induced CD3+ CD56+ killer cells. Blood 1995, 86, 3493–3499. [Google Scholar] [PubMed]
- Vankayalapati, R.; Garg, A.; Porgador, A.; Griffith, D.E.; Klucar, P.; Safi, H.; Girard, W.M.; Cosman, D.; Spies, T.; Barnes, P.F. Role of NK cell-activating receptors and their ligands in the lysis of mononuclear phagocytes infected with an intracellular bacterium. J. Immunol. 2005, 175, 4611–4617. [Google Scholar] [CrossRef] [PubMed]
- De Libero, G.; Moran, A.P.; Gober, H.J.; Rossy, E.; Shamshiev, A.; Chelnokova, O.; Mazorra, Z.; Vendetti, S.; Sacchi, A.; Prendergast, M.M.; et al. Bacterial infections promote T cell recognition of self-glycolipids. Immunity 2005, 22, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Proft, T.; Fraser, J.D. Bacterial superantigens. Clin. Exp. Immunol. 2003, 133, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Ljunggren, H.G.; Sarvetnick, N. Innate immunity and autoimmunity: From self-protection to self-destruction. Trends Immunol. 2001, 22, 97–101. [Google Scholar] [CrossRef]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torina, A.; Guggino, G.; La Manna, M.P.; Sireci, G. The Janus Face of NKT Cell Function in Autoimmunity and Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 440. https://doi.org/10.3390/ijms19020440
Torina A, Guggino G, La Manna MP, Sireci G. The Janus Face of NKT Cell Function in Autoimmunity and Infectious Diseases. International Journal of Molecular Sciences. 2018; 19(2):440. https://doi.org/10.3390/ijms19020440
Chicago/Turabian StyleTorina, Alessandra, Giuliana Guggino, Marco Pio La Manna, and Guido Sireci. 2018. "The Janus Face of NKT Cell Function in Autoimmunity and Infectious Diseases" International Journal of Molecular Sciences 19, no. 2: 440. https://doi.org/10.3390/ijms19020440
APA StyleTorina, A., Guggino, G., La Manna, M. P., & Sireci, G. (2018). The Janus Face of NKT Cell Function in Autoimmunity and Infectious Diseases. International Journal of Molecular Sciences, 19(2), 440. https://doi.org/10.3390/ijms19020440