Can Plant Defence Mechanisms Provide New Approaches for the Sustainable Control of the Two-Spotted Spider Mite Tetranychus urticae?



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Biological Characteristics of T. urticae that Favour Its Development into a Devastating Pest of Crops
2.1. Life Cycle and Reproduction
2.2. High Ability to Adapt to Harsh Environmental Conditions
2.3. Detoxification and Adaptation to Pesticides
3. Current Management Tools to Control T. urticae
3.1. Secondary Pest Outbreaks Triggered by Unsuitable Management
4. Plant Defence Mechanisms that Contribute to a Sustainable Control of T. urticae
4.1. Pre-Existing Constitutive Defences against T. urticae
4.2. Inducible Defences against T. urticae
4.2.1. Hormone Signalling Pathways Activated Upon Mite Infestation
4.2.2. Plant Secondary Metabolites in Plant Defence against T. urticae
4.2.3. Defensive Proteins and Peptides against T. urticae
4.2.4. Induced Responses Triggering Direct Systemic Defence against T. urticae
4.2.5. Herbivore-Induced Plant Volatiles (HIPVs) in Plant Resistance
5. Top–Down and Indirect Defence Mechanisms against T. urticae
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Oakland, CA, USA, 1975. [Google Scholar]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: A review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Fleschner, C.A.; Badgley, M.E.; Ricker, D.W.; Hall, J.C. Air drift of spider mites. J. Econ. Entomol. 1956, 49, 624–627. [Google Scholar] [CrossRef]
- Saito, Y. Life types of spider mites. In Spider Mites. Their biology, Natural Enemies and Control; Helle, W., Sabelis, M.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; Volume 1, pp. 253–264. [Google Scholar]
- Saito, Y. Plant Mite and Sociality: Diversity and Evolution; Springer: Tokyo, Japan, 2010. [Google Scholar]
- Helle, W.; Sabelis, M.W. Spider Mites. Their biology, Natural Enemies and Control; Elsevier: Amsterdam, The Netherlands, 1985; Volume 1, p. 458. [Google Scholar]
- Bolland, H.R.; Gutierrez, J.; Flechtmann, C.H. World Catalogue of the Spider Mite Family (Acari: Tetranychidae); Brill: Leiden, The Netherlands, 1998. [Google Scholar]
- Migeon, A.; Nouguier, E.; Dorkeld, F. Spider Mites Web: A Comprehensive Database for the Tetranychidae. In Trends in Acarology; Springer: Amsterdam, The Netherlands, 2010; pp. 557–560. [Google Scholar]
- Grbić, M.; van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouzé, P.; Grbić, V.; Verdon, L. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attia, S.; Grissa, K.L.; Lognay, G.; Bitume, E.; Hance, T.; Mailleux, A.C. A review of the major biological approaches to control the worldwide pest Tetranychus urticae (Acari: Tetranychidae) with special reference to natural pesticides. J. Pest Sci. 2013, 86, 361–386. [Google Scholar] [CrossRef]
- Ohtsuka, K.; Osakabe, M. Deleterious effects of UV-B radiation on herbivorous spider mites: They can avoid it by remaining on lower leaf surfaces. Environ. Entomol. 2009, 38, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.T.; Lyon, H.H. Insects that Feed on Trees and Shrubs, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1991. [Google Scholar]
- Hoy, M.A. Agricultural Acarology: Introduction to Integrated Mite Management; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Huffaker, C.B.; Vrie, M.V.; McMurtry, J.A. The ecology of tetranychid mites and their natural control. Annu. Rev. Entomol. 1969, 14, 125–174. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Grbic, M.; Khila, A.; Lee, K.-Z.; Bjelica, A.; Grbic, V.; Whistlecraft, J.; Verdon, L.; Navajas, M.; Nagy, L. Mity model: Tetranychus urticae, a candidate for chelicerate model organism. Bioassays 2007, 29, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Helle, W.; Gutierrez, J.; Bolland, H.R. A study on sex-determination and karyotypic evolution in Tetranychidae. Genetica 1970, 41, 21–32. [Google Scholar] [CrossRef]
- Werren, J.H. Biology of wolbachia. Annu. Rev. Entomol. 1997, 42, 587–609. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and evolution of heritable bacterial symbionts. Ann. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Brownlie, J.C.; Johnson, K.N. Symbiont-mediated protection in insect hosts. Trends Microbiol. 2009, 17, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Gross, R.; Vavre, F.; Heddi, A.; Hurst, G.D.; Zchori-Fein, E.; Bourtzis, K. Immunity and symbiosis. Mol. Microbiol. 2009, 73, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Cook, P.E.; McGraw, E.A. Wolbachia pipientis: An expanding bag of tricks to explore for disease control. Trends Parasitol. 2010, 26, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Breeuwer, J.A.; Werren, J.H. Cytoplasmic incompatibility and bacterial density in Nasonia vitripennis. Genetics 1993, 135, 565–574. [Google Scholar] [PubMed]
- Hunter, M.D.; McNeil, J.N. Host-plant quality influences diapause and voltinism in a polyphagous insect herbivore. Ecology 1997, 78, 977–986. [Google Scholar] [CrossRef]
- Veerman, A. Diapause. In Spider Mites. Their biology, Natural Enemies and Control; Helle, W., Sabelis, M.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; Volume 1, pp. 279–316. [Google Scholar]
- Foott, W.H. Geotactic response of the two-spotted spider mite, Tetranychus urticae Koch (Acarina, Tetranychidae). Proc. Entomol. Soc. Ont. 1965, 95, 106–108. [Google Scholar]
- Bryon, A.; Wybouw, N.; Dermauw, W.; Tirry, L.; van Leeuwen, T. Genome wide gene-expression analysis of facultative reproductive diapause in the two-spotted spider mite Tetranychus urticae. BMC Genomics 2013, 14, 815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodayari, S.; Moharramipour, S.; Larvor, V.; Hidalgo, K.; Renault, D. Deciphering the metabolic changes associated with diapause syndrome and cold acclimation in the two-spotted spider mite Tetranychus urticae. PLoS ONE 2013, 8, e54025. [Google Scholar] [CrossRef]
- Dermauw, W.; Wybouw, N.; Rombauts, S.; Menten, B.; Vontas, J.; Grbic, M.; Clark, R.M.; Feyereisen, R.; Van, L.T. A link between host plant adaptation and pesticide resistance in the polyphagous spider mite Tetranychus urticae. Proc. Natl. Acad. Sci. USA 2013, 110, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Martel, C.; Zhurov, V.; Navarro, M.; Martinez, M.; Cazaux, M.; Auger, P.; Migeon, A.; Santamaria, M.E.; Wybouw, N.; Diaz, I.; et al. Tomato whole genome transcriptional response to Tetranychus urticae identifies divergence of spider mite-induced responses between tomato and Arabidopsis. Mol. Plant Microbe Interact. 2015, 28, 343–361. [Google Scholar] [CrossRef] [PubMed]
- Wybouw, N.; Zhurov, V.; Martel, C.; Bruinsma, K.A.; Hendrickx, F.; Grbić, V.; van Leeuwen, T. Adaptation of a polyphagous herbivore to a novel host plant extensively shapes the transcriptome of herbivore and host. Mol. Ecol. 2015, 24, 4647–4663. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Riquelme, J.; Zhurov, V.; Rioja, C.; Pérez-Moreno, P.; Badja, S.; van Leeuwen, T.; Martínez-Zapater, J.M.; Grbic, M.; Grbic, V. Comparative genome-wide transcriptome analysis of Vitis vinifera responses to adapted and non-adapted strains of two-spotted spider mite, Tetranuchus urticae. BMC Genomics 2016, 17, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaría, M.E.; González-Cabrera, J.; Martínez, M.; Grbic, V.; Castañera, P.; Díaz, I.; Ortego, F. Digestive proteases in bodies and faeces of the two-spotted spider mite, Tetranychus urticae. J. Insect. Phys. 2015, 78, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Hussey, N.W.; Parr, W.J. Dispersal of the glasshouse red spider mite Tetranychus urticae Koch (Acarina, Tetranychidae). Entomol. Exp. Appl. 1963, 6, 207–214. [Google Scholar] [CrossRef]
- Li, J.; Margolies, D.C. Effects of mite age, mite density, and host quality on aerial dispersal behavior in the two spotted spider mite. Entomol. Exp. Appl. 1993, 68, 79–86. [Google Scholar] [CrossRef]
- Osakabe, M.H.; Isobe, H.; Kasai, A.; Masuda, R.; Kubota, S.; Umeda, M. Aerodynamic advantages of upside down take-off for aerial dispersal in Tetranychus spider mites. Exp. Appl. Acarol. 2008, 44, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Clotuche, G.; Mailleux, A.C.; Fernández, A.A.; Deneubourg, J.L.; Detrain, C.; Hance, T. The formation of collective silk balls in the spider mite Tetranychus urticae Koch. PLoS ONE 2011, 6, e18854. [Google Scholar] [CrossRef] [PubMed]
- Knowles, C.O. Mechanisms of resistance to acaricides. In Molecular Mechanisms of Resistance to Agrochemicals; Springer Science & Business Media: Berlin, Germany, 1997; pp. 57–77. [Google Scholar]
- Van Leeuwen, T.; Vanholme, B.; van Pottelberge, S.; van Nieuwenhuyse, P.; Nauen, R.; Tirry, L.; Denholm, I. Mitochondrial heteroplasmy and the evolution of insecticide resistance: Non-Mendelian inheritance in action. Proc. Natl. Acad. Sci. USA 2008, 105, 5980–5985. [Google Scholar] [CrossRef] [PubMed]
- Michigan State University. 2017. Available online: https://www.pesticideresistance.org/search.php (accessed on 21 December 2017).
- IRAC. IRAC MoA Classification Scheme. Version 8.3. 2017. Available online: http://www.irac-online.org/documents/moa-classification/?ext=pdf (accessed on 21 December 2017).
- Rabinge, R. Aspects of damage assessment. In Spider Mites. Their Biology, Natural Enemies and Control; Helle, W., Sabelis, M.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; Volume B, pp. 261–272. [Google Scholar]
- Marcic, D. Acaricides in modern management of plant-feeding mites. J. Pest Sci. 2012, 85, 395–408. [Google Scholar] [CrossRef]
- Van Leeuven, T.; Dermauw, W.; Grbic, M.; Tirry, L.; Feyereisen, R. Spider mite control and resistance management: Does a genome help? Pest Manag. Sci. 2013, 69, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Dekeyser M, A. Acaricide mode of action. Pest Manag. Sci. 2005, 61, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Ruiz, S.; Aguilar-Fenollosa, E.; Ibáñez-Gual, V.; Hurtado-Ruiz, M.A.; Martínez-Ferrer, M.T.; Jacas, J.A. Economic threshold for Tetranychus urticae (Acari: Tetranychidae) in clementine mandarins Citrus clementina. Exp. Appl. Acarol. 2014, 62, 337–362. [Google Scholar] [CrossRef] [PubMed]
- García-Marí, F.; González-Zamora, J.E. Biological control of Tetranychus urticae (Acari: Tetranychidae) with naturally occurring predators in strawberry plantings in Valenica, Spain. Exp. Appl. Acarol. 1999, 23, 487–495. [Google Scholar] [CrossRef]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for unifying the terminology in biological control. BioControl 2001, 46, 46–387. [Google Scholar] [CrossRef]
- De Moraes, G.J.; McMurtry, J.A.; Denmark, H.A.; Cambos, C.B. A revised catalog of the mite family Phytoseiidae. Zootaxa 2004, 434, 1–494. [Google Scholar] [CrossRef]
- Mcmurtry, J.A.; de Moraes, G.J.; Sourassou, N.F. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol. 2013, 18, 297–320. [Google Scholar] [CrossRef]
- Croft, B.A.; Brown, A.W.A. Responses of arthropod natural enemies to insecticides. Annu. Rev. Entomol. 1975, 20, 285–335. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.D.; Vargas, R.; Banks, J.E. Incorporating ecologically relevant measures of pesticide effect for estimating the compatibility of pesticides and biocontrol agents. J. Econ. Entomol. 2007, 100, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Li, J.; Shao, Y.; Yang, B.; Liu, Z. Fipronil resistance in the whitebacked planthopper (Sogatella furcifera): Possible resistance mechanisms and cross-resistance. Pest Manag. Sci. 2010, 66, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Cloyd, R.A. Indirect Effects of Pesticides on Natural Enemies. Pesticides—Advances in Chemical and Botanical Pesticides; Intech: Rijeka, Croatia, 2012; pp. 127–150. [Google Scholar]
- Longley, M.; Jepson, P.C. Effects of honeydew and insecticide residues on the distribution of foraging aphid parasitoids under glasshouse and field conditions. Entomol. Exp. Appl. 1996, 81, 189–198. [Google Scholar] [CrossRef]
- Desneux, N.; Rafalimanana, H.; Kaiser, L. Dose–response relationship in lethal and behavioural effects of different insecticides on the parasitic wasp Aphidius ervi. Chemosphere 2004, 54, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Costa Comelles, J.; Bosch Serra, À.D.; Botargues, A.; Cabiscol, P.; Moreno, A.; Portillo, J.; Avilla Hernández, J. Acción de algunos acaricidas sobre los fítoseídos y la araña roja Panonychus ulmi (Koch) en manzano. Boletín de Sanidad Vegetal. Plagas 1997, 23, 93–103. [Google Scholar]
- Stanyard, M.J.; Foster, R.E.; Gibb, T.J. Population dynamics of Amblyseius fallacies (Acari: Phytoseiidae) and european red mite (Acari: Tetranychidae) in apple trees treated with selected acaricides. J. Econ. Entomol. 1998, 91, 217–225. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Magalhaes, L.C.; Cosme, L.V. Stimulatory sublethal response of a generalist predator to permethrin: Hormesis, hormoligosis, or homeostatic regulation? J. Econ. Entomol. 2009, 102, 170–176. [Google Scholar] [CrossRef] [PubMed]
- James, D.G.; Price, T.S. Fecundity in two spotted spider mite (Acari: Tetranychidae) is increased by direct and systemic exposure to imidacloprid. J. Econ. Entomol. 2002, 95, 729–732. [Google Scholar] [CrossRef] [PubMed]
- Marcic, D. The effects of clofentezine on life-table parameters in two-spotted spider mite Tetranychus urticae. Exp. Appl. Acarol. 2003, 30, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Garvey, M.K.I.; Li, B.; Carrillo, J. Domestication of tomato has reduced the attraction of herbivore natural enemies to pest-damaged plants. Agric. For. Entomol. 2017. [Google Scholar] [CrossRef]
- Brummer, C.E.; Barber, W.T.; Collier, S.M.; Cox, T.S.; Johnson, R.; Murray, S.C.; Olsen, R.T.; Pratt, R.C.; Thro, A.M. Plant breeding for harmony between agriculture and the environment. Front. Ecol. Environ. 2011, 9, 561–568. [Google Scholar] [CrossRef]
- Carmona, D.; Lajeunesse, M.J.; Johnson, M.T. Plant traits that predict resistance to herbivores. Funct. Ecol. 2011, 25, 358–367. [Google Scholar] [CrossRef]
- Stowe, K.A.; Marquis, R.J.; Hochwender, C.G.; Simms, E.L. The evolutionary ecology of tolerance to consumer damage. Annu. Rev. Ecol. Syst. 2000, 31, 565–595. [Google Scholar] [CrossRef]
- Schaller, A.; Howe, G.A. Direct Defenses in Plants and Their Induction by Wounding and Insect Herbivores. In Induced Plant Resistance to Herbivory; Schaller, A., Ed.; Springer Science & Business Media: Berlin, Germany, 2008; pp. 7–29. [Google Scholar]
- Wittstock, U.; Gershenzon, J. Constitutive plant toxins and their role in defense against herbivores and pathogens. Curr. Opin. Plant Biol. 2002, 5, 300–307. [Google Scholar] [CrossRef]
- Huchelmann, A.; Boutry, M.; Hachez, C. Plant glandular trichomes: Natural cell factories of high biotechnological interest. Plant Physiol. 2017, 175, 6–22. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Tooker, J.; Peiffer, M.; Chung, S.H.; Felton, G.W. Role of trichomes in defense against herbivores: comparison of herbivore response to woolly and hairless trichome mutants in tomato (Solanum lycopersicum). Planta 2012, 236, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Karley A., J.; Mitchell, C.; Brookes, C.; McNicol, J.; O’Neill, T.; Roberts, H.; Graham, J.; Johnson, S.N. Exploiting physical defence traits for crop protection: leaf trichomes of Rubus idaeus have deterrent effects on spider mites but not aphids. Annu. Appl. Biol. 2016, 168, 159–172. [Google Scholar] [CrossRef]
- Eigenbrode, S.D. The effects of plant epicuticular waxy blooms on attachment and effectiveness of predatory insects. Arthropod. Struct. Dev. 2004, 33, 91–102. [Google Scholar] [CrossRef] [PubMed]
- VanEtten, H.D.; Mansfield, J.W.; Bailey, J.A.; Farmer, E.E. Two classes of plant antibiotics: Phytoalexins versus “phytoanticipins”. Plant Cell 1994, 6, 1191–1192. [Google Scholar] [CrossRef] [PubMed]
- Stauber, E.J.; Kuczka, P.; van Ohlen, M.; Vogt, B.; Janowitz, T.; Piotrowski, M.; Beuerle, T.; Wittstock, U. Turning the “mustard oil bomb” into a “cyanide bomb”: Aromatic glucosinolate metabolism in a specialist insect herbivore. PLoS ONE 2012, 7, e35545. [Google Scholar] [CrossRef] [PubMed]
- Miresmailli, S.; Isman, M.B. Botanical insecticides inspired by plant–herbivore chemical interactions. Trends Plant Sci. 2014, 19, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Rioja, C.; Zhurov, V.; Bruinsma, K.; Grbic, M.; Grbic, V. Plant-herbivore interactions: A case of an extreme generalist, the two-spotted spider mite Tetranychus urticae. MPMI 2017, 30, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Alba, J.M.; Schimmel, B.C.J.; Glas, J.J.; Ataide, L.M.S.; Pappas, M.L.; Villarroel, C.A.; Schuurink, R.C.; Sabbelis, M.W.; Kant, M.R. Spider mites suppress tomato defenses downstream of jasmonate and salicylate independently of hormonal crosstalk. New Phytol. 2015, 205, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Sarmento, R.A.; Lemos, F.; Bleeker, P.M.; Schuurink, R.C.; Pallini, A.; Oliveira, M.G.A.; Lima, E.R.; Kant, M.; Sabelis, M.W.; Janssen, A. A herbivore that manipulates plant defence. Ecol. Lett. 2011, 14, 229–236. [Google Scholar]
- Pieterse, C.M.; Leon-Reyes, A.; van der Ent, S.; van Wees, S.C. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Villarroel, C.A.; Jonckheere, W.; Alba, J.M.; Glas, J.J.; Dermauw, W.; Haring, M.A.; van Leeuwen, T.; Schuurink, R.C.; Kant, M.R. Salivary proteins of spider mites suppress defenses in Nicotiana benthamiana and promote mite reproduction. Plant J. 2016, 86, 119–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensoussan, N.; Santamaria, M.E.; Zhurov, V.; Diaz, J.; Grbić, M.; Grbić, V. Plant–herbivore interaction: dissection of the cellular pattern of Tetranychus urticae feeding on the host plant. Front. Plant Sci. 2016, 7, 1105. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Mithöfer, A.; Boland, W. Recognition of herbivory-Associated Molecular Patterns. Plant Phys. 2008, 146, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.I.; Ozawa, R.; Nishioka, T.; Boland, W.; Koch, T.; Kühnemann, F.; Takabayashi, J. Herbivore-induced volatiles induce the emission of ethylene in neighboring lima bean plants. Plant J. 2002, 29, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Leitner, M.; Boland, W.; Mithöfer, A. Direct and indirect defenses induced by piercing-sucking and chewing herbivores in Medicago truncatula. New Phytol. 2005, 167, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Ament, K.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Differential timing of spider mite-induced direct and indirect defenses in tomato plants. Plant Phys. 2004, 135, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Kawazu, K.; Mochizuki, A.; Sato, Y.; Sugeno, W.; Murata, M.; Seo, S.; Mitsuhara, I. Different expression profiles of jasmonic acid and salicylic acid inducible genes in the tomato plant against herbivores with various feeding modes. Arthropod-Plant Interact. 2012, 6, 221–230. [Google Scholar] [CrossRef]
- Zhurov, V.; Navarro, M.; Bruinsma, K.A.; Arbona, V.; Santamaria, M.E.; Cazaux, M.; Wybouw, N.; Osborne, E.J.; Ens, C.; Rioja, C.; et al. Reciprocal responses in the interaction between Arabidopsis and the cell-content-feeding chelicerate herbivore spider mite. Plant Physiol. 2014, 164, 384–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maserti, B.E.; del Carratore, R.; Della Croce, C.M. Comparative analysis of proteome changes induced by the two-spotted spider mite Tetranychus urticae and methyl jasmonate in citrus leaves. J. Plant Phys. 2011, 168, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Agut, B.; Gamir, J.; Jacas, J.A.; Hurtado, M.; Flors, V. Different metabolic and genetic responses in citrus may explain relative susceptibility to Tetranychus urticae. Pest Manag. Sci. 2014, 70, 1728–1741. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Intraspecific variation in a generalist herbivore accounts for differential induction and impact of host plant defenses. Proc. R Soc. B: Biol. Sci. 2008, 275, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Jonckheere, W.; Dermauw, W.; Khalighi, M.; Pavlidi, N.; Reubens, W.; Baggerman, G.; Tirry, L.; Menschaert, G.; Kant, M.R.; Vanholeme, B.; et al. A gene family coding for salivary proteins (SHOT) of the polyphagous spider mite Tetranychus urticae exhibits fast host-dependent transcriptional plasticity. MPMI 2018, 31, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Glas, J.J.; Alba, J.M.; Simoni, S.; Villarroel, C.A.; Stoops, M.; Schimmel, B.C.J.; Schuurink, R.C.; Sabelis, M.W.; Kant, M.R. Defense suppression benefits herbivores that have a monopoly on their feeding site but can backfire within natural communities. BMC Biol. 2014, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Castañé, C.; Arnó, J.; Gabarra, R.; Alomar, O. Plant damage to vegetable crops by zoophytophagus mirid predators. Biol. Control 2011, 59, 22–29. [Google Scholar] [CrossRef]
- Pappas, M.L.; Steppuhn, A.; Geuss, D.; Topalidou, N.; Aografou, A.; Sabelis, M.W.; Broufas, G.D. Beyond predation: The zoophytophagous predator macrolophus pygmaeus induces tomato resistance against spider mites. PLoS ONE 2015, 10, e0127251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.J.; Zheng, S.J.; van Loon, J.J.; Boland, W.; David, A.; Mumm, R.; Dicke, M. Whiteflies interfere with indirect plant defense against spider mites in Lima bean. Proc. Nat. Acad. Sci. USA 2009, 106, 21202–21207. [Google Scholar] [CrossRef] [PubMed]
- Bleeker, P.M.; Mirabella, R.; Diergaarde, P.J.; VanDoorn, A.; Tissier, A.; Kant, M.R.; Schuurink, R.C. Improved herbivore resistance in cultivated tomato with the sesquiterpene biosynthetic pathway from a wild relative. Proc. Nat. Acad. Sci. USA 2012, 109, 20124–20129. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, S.C.H.; Niculau, E.D.S.; Blank, A.F.; Câmara, C.A.G.; Araújo, I.N.; Alves, P.B. Composition and acaricidal activity of Lippia sidoides essential oil against two-spotted spider mite (Tetranychus urticae Koch). Bioresour Technol. 2010, 101, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Bleeker, P.M.; Diergaarde, P.J.; Ament, K.; Schütz, S.; Johne, B.; Dijkink, J.; Hiemstra, H.; de Gelder, R.; de Both, M.T.J.; Sabelis, M.W.; et al. Tomato-produced 7-epizingiberene and R-curcumene act as repellents to whiteflies. Phytochemistry 2011, 72, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Gordon-Weeks, R.U.T.H.; Pickett, J.; Ton, J. Natural variation in priming of basal resistance: from evolutionary origin to agricultural exploitation. Mol. Plant Pathol. 2010, 11, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Schlaeppi, K.; Abou-Mansour, E.; Buchala, A.; Mauch, F. Disease resistance of Arabidopsis to Phytophthora brassicae is established by the sequential action of indole glucosinolates and camalexin. Plant J. 2010, 62, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Winde, I.; Wittstock, U. Insect herbivore counteradaptations to the plant glucosinolate-myrosinase system. Phytochemistry 2011, 72, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Badenes-Perez, F.R.; Reichelt, M.; Gershenzon, J.; Heckel, D.G. Interaction of glucosinolate content of Arabidopsis thaliana mutant lines and feeding and oviposition by generalist and specialist lepidopterans. Phytochemistry 2013, 86, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, M.E.; Martínez, M.; Arnaiz, A.; Ortego, F.; Grbic, V.; Diaz, I. MATI, a novel protein involved in the regulation of herbivore-associated signaling pathways. Front. Plant Sci. 2017, 8, 975. [Google Scholar] [CrossRef] [PubMed]
- Pourcel, L.; Irani, N.G.; Koo, A.J.; Bohorquez-Restrepo, A.; Howe, G.A.; Grotewold, E. A chemical complementation approach reveals genes and interactions of flavonoids with other pathways. Plant J. 2013, 74, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Onkokesung, N.; Reichelt, M.; van Doorn, A.; Schuurink, R.C.; van Loon, J.J.; Dicke, M. Modulation of flavonoid metabolites in Arabidopsis thaliana through overexpression of the MYB75 transcription factor: role of kaempferol-3, 7-dirhamnoside in resistance to the specialist insect herbivore Pieris brassicae. J. Exp. Bot. 2014, 65, 2203–2217. [Google Scholar] [CrossRef] [PubMed]
- Agut, B.; Gamir, J.; Jaques, J.A.; Flors, V. Tetranychus urtichae-triggered responses promote genotype-dependent conspecific repellence or attractiveness in citrus. New Phyt. 2015, 207, 790–804. [Google Scholar] [CrossRef] [PubMed]
- Mauch-Mani, B.; Bacelli, I.; Luna, E.; Flors, V. Defense priming: An adaptative part of induced resistance. Annu. Rev. Plant. Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.; Pozo, M.J.; Ton, J.; van Dam N., M.; Conrath, U. Recognizing plant defense priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Alba, J.M.; Montserrat, M.; Fernández-Muñoz, R. Resistance to the two-spotted spider mite (Tetranychus urticae) by acylsucroses of wild tomato (Solanum pimpinellifolium) trichomes studied in a recombinant inbred line population. Exp. Appl. Acarol. 2009, 47, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Hare, J.D.; Walling, L.L. Constitutive and jasmonate-inducible traits of Datura wrightii. J. Chem. Ecol. 2006, 32, 29–47. [Google Scholar] [CrossRef] [PubMed]
- Green, T.R.; Ryan, C.A. Wound-induced proteinase inhibitor in plant leaves: a possible defense mechanism against insects. Science 1972, 175, 776–777. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.A. Protease inhibitors in plants: Genes for improving defenses against insects and pathogens. Annu. Rev. Phytopathol. 1990, 28, 425–449. [Google Scholar] [CrossRef]
- Lison, P.; Rodrigo, I.; Conejero, V. A novel function for the cathepsin D inhibitor in tomato. Plant Phys. 2006, 142, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Zavala, J.A.; Patankar, A.G.; Gase, K.; Hui, D.Q.; Baldwin, I.T. Manipulation of endogenous trypsin proteinase inhibitor production in Nicotiana attenuata demonstrates their function as antiherbivore defenses. Plant Phys. 2004, 134, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Talyzina, N.M.; Ingvarsson, P.K. Molecular evolution of a small gene family of wound inducible Kunitz trypsin inhibitors in Populus. J. Mol. Evol. 2006, 63, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, L.; Martinez, M.; Álvarez-Alfageme, F.; Castañera, P.; Smagghe, G.; Diaz, I.; Ortego, F. A barley cysteine-proteinase inhibitor reduces the performance of two aphid species in artificial diets and transgenic Arabidopsis plants. Transgenic Res 2011, 20, 305–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaria, M.E.; Cambra, I.; Martinez, M.; Pozancos, C.; Gonzalez-Melendi, P.; Grbic, V.; Castañera, P.; Ortego, F.; Diaz, I. Gene pyramiding of peptidase inhibitors enhances plant resistance to the spider mite Tetranychus urticae. PLoS ONE 2012, 7, e43011. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; van Wees, S.C.; Bakker, P.A. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Köllner, T.G.; Degenhardt, J.; Zwahlen, C.; Hibbard, B.E.; Turlings, T.C. The role of abscisic acid and water stress in root herbivore-induced leaf resistance. New Phytologist 2011, 189, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Karban, R.; Carey, J.R. Induced resistance of cotton seedlings to mites. Science 1984, 225, 53–54. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.A.; Chauvin, A.; Pascaud, F.; Kellenberger, S.; Farmer, E.E. GLUTAMATE RECEPTOR-LIKE genes mediate leaf-to-leaf wound signalling. Nature 2013, 500, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Agut, B.; Gamir, J.; Jaques, J.A.; FLors, V. Systemic resistance in citrus to Tetranychus urticae induced by conspecifics is transmitted by grafting mediated by mobile amino acids. J. Exp. Bot. 2016, 67, 5711–5723. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M. Volatile spider-mite pheromone and host-plant kairomone, involved in spaced-out gregariousness in the spider mite Tetranychus urticae. Physiol. Entomol. 1986, 11, 251–262. [Google Scholar] [CrossRef]
- Pallini, A.; Janssen, A.; Sabelis, M.W. Odour-mediated responses of phytophagous mites to conspecific and heterospecific competitors. Oecologia 1997, 110, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Frost, C.J.; Mescher, M.C.; Carlson, J.E.; De Moraes, C.M. Plant defense priming against herbivores: getting ready for a different battle. Plant Physiol. 2008, 146, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.; Schultz, J.C. Talking trees. Science 1983, 221, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proc. Nat. Acad. Sci. USA 2004, 101, 1781–1785. [Google Scholar] [CrossRef] [PubMed]
- Muroi, A.; Ramadan, A.; Nishihara, M.; Yamamoto, M.; Ozawa, R.; Takabayashi, J.; Arimura, G.I. The composite effect of transgenic plant volatiles for acquired immunity to herbivory caused by inter-plant communications. PLoS ONE 2011, 6, e24594. [Google Scholar] [CrossRef] [PubMed]
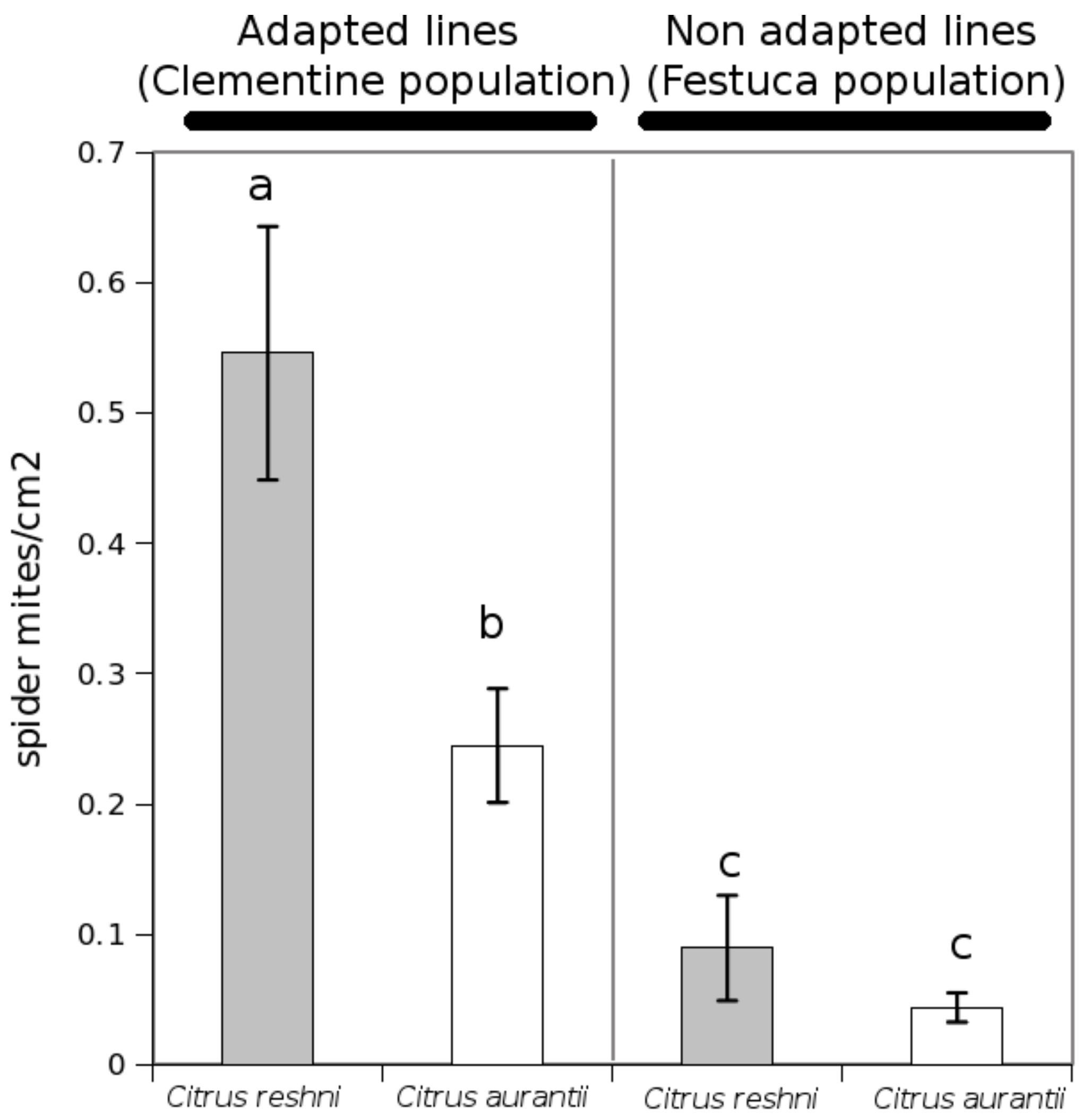
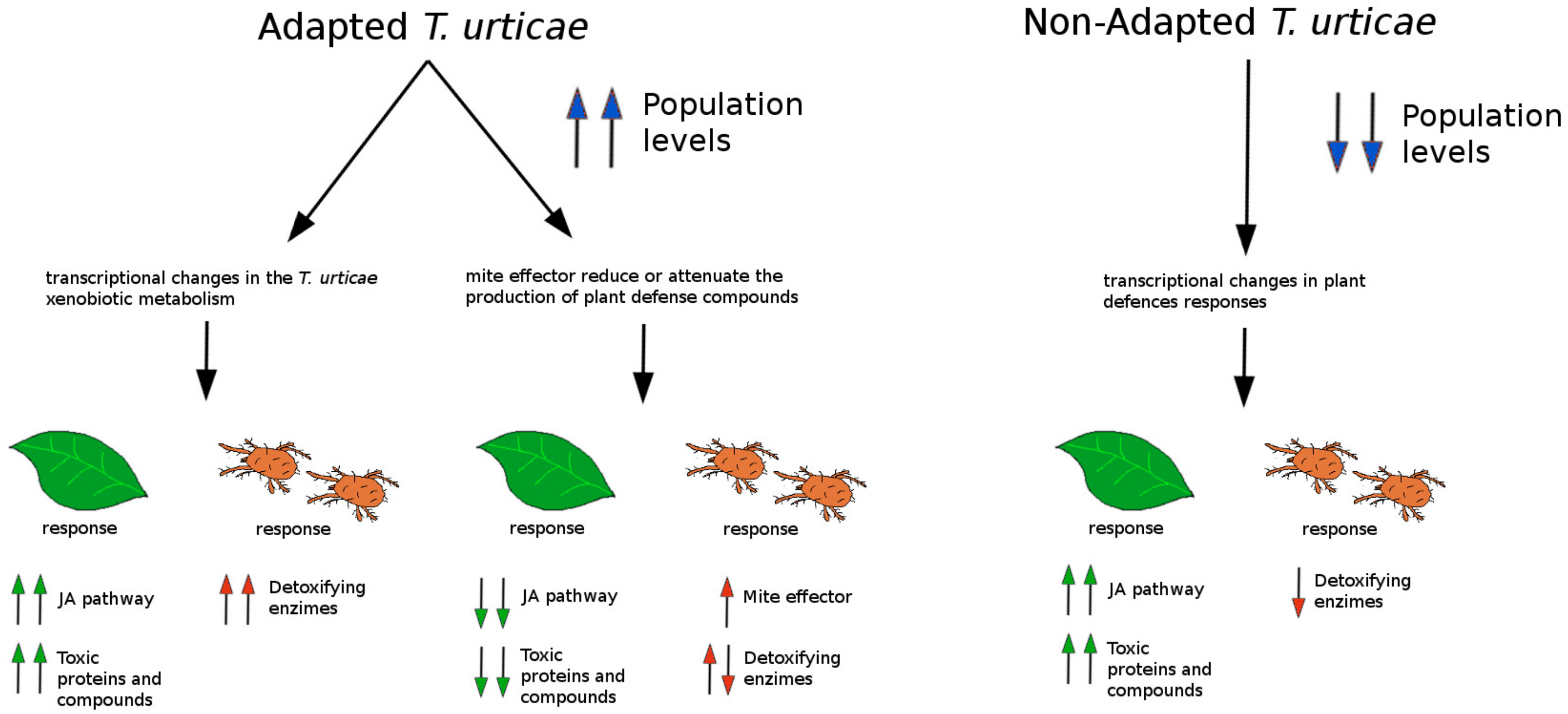
- Aguilar-Fenollosa, E.; Pina, T.; Gómez-Martínez, M.A.; Hurtado, M.A.; Jacas, J.A. Does host adaptation of Tetranychus urticae populations in clementine orchards with a Festuca arundinacea cover contribute to a better natural regulation of this pest mite? Entomol. Exp. App. 2012, 144, 181–190. [Google Scholar] [CrossRef]
- Van Wijk, M.; de Bruijn, P.J.; Sabelis, M.W. Predatory mite attraction to herbivore-induced plant odors is not a consequence of attraction to individual herbivore-induced plant volatiles. J. Chem. Ecol. 2008, 34, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Fenollosa, E.; Ibáñez-Gual, M.V.; Pascual-Ruiz, S.; Hurtado, M.; Jacas, J.A. Effect of ground-cover management on spider mites and their phytoseiid natural enemies in clementine mandarin orchards (I): Bottom-up regulation mechanisms. Biol. Control 2011, 59, 158–170. [Google Scholar] [CrossRef]
- De Boer, J.G.; Dicke, M. The role of methyl salicylate in prey searching behavior of the predatory mite Phytoseiulus persimilis. J. Chem. Ecol. 2004, 30, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Ament, K.; Krasikov, V.; Allmann, S.; Rep, M.; Takken, F.L.; Schuurink, R.C. Methyl salicylate production in tomato affects biotic interactions. Plant J. 2010, 62, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Kappers, I.F.; Hoogerbrugge, H.; Bouwmeester, H.J.; Dicke, M. Variation in herbivory-induced volatiles among cucumber (Cucumis sativus L.) varieties has consequences for the attraction of carnivorous natural enemies. J. Chem. Ecol. 2011, 37, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Brillada, C.; Nishihara, M.; Shimoda, T.; Garms, S.; Boland, W.; Maffei, M.E.; Arimura, G.I. Metabolic engineering of the C16 homoterpene TMTT in Lotus japonicus through overexpression of (E,E)-geranyllinalool synthase attracts generalist and specialist predators in different manners. New Phytol. 2013, 200, 1200–1211. [Google Scholar] [CrossRef] [PubMed]
- Pozo, M.J.; Jung, S.C.; Martínez-Medina, A.; López-Ráez, J.A.; Azcón-Aguilar, C.; Barea, J.M. Root allies: Arbuscular mycorrhizal fungi help plants to cope with biotic stresses. In Symbiotic Endophytes; Springer Science & Business Media: Berlin, Germany, 2013; pp. 289–307. [Google Scholar]
- Schausberger, P.; Peneder, S.; Juerschik, S.; Hoffmann, D. Mycorrhiza changes plant volatiles to attract spider mite enemies. Func. Ecol. 2012, 26, 441–449. [Google Scholar] [CrossRef]
- Rashid, M.H.; Chung, Y.R. Induction of systemic resistance against insect herbivores in plants by beneficial soil microbes. Front. Plant Sci. 2017, 8, 1816. [Google Scholar] [CrossRef] [PubMed]
- Sharma, E.; Anand, G.; Kapoor, R. Terpenoids in plant and arbuscular mycorrhiza-reinforced defence against herbivorous attack. Ann. Bot. 2017, 119, 791–801. [Google Scholar] [PubMed]
- Mérey, G.V.; Veyrat, N.; Mahuku, G.; Valdez, R.L.; Turlings, T.C.; D’Alessandro, M. Dispensing synthetic green leaf volatiles in maize fields increases the release of sesquiterpenes by the plants, but has little effect on the attraction of pest and beneficial insects. Phytochemistry 2011, 72, 1838–1847. [Google Scholar] [CrossRef] [PubMed]
- Uefune, M.; Choh, Y.; Abe, J.; Shiojiri, K.; Sano, K.; Takabayashi, J. Application of synthetic herbivore-induced plant volatiles causes increased parasitism of herbivores in the field. J. App. Entomol. 2012, 136, 561–567. [Google Scholar] [CrossRef]
- Sun, X.L.; Wang, G.C.; Gao, Y.; Chen, Z.M. Screening and field evaluation of synthetic volatile blends attractive to adults of the tea weevil, Myllocerinus aurolineatus. Chemoecology 2012, 22, 229–237. [Google Scholar] [CrossRef]
- Simpson, M.; Gurr, G.M.; Simmons, A.T.; Wratten, S.D.; James, D.G.; Leeson, G.; Nicol, H.I. Insect attraction to synthetic herbivore-induced plant volatile-treated field crops. Agr. For. Entomol. 2011, 13, 45–57. [Google Scholar] [CrossRef]
- Rodriguez-Saona, C.; Kaplan, I.; Braasch, J.; Chinnasamy, D.; Williams, L. Field responses of predaceous arthropods to methyl salicylate: A meta-analysis and case study in cranberries. Biol. Control 2011, 59, 294–303. [Google Scholar] [CrossRef]
- Adar, E.; Inbar, M.; Gal, S.; Gan-Mor, S.; Palevsky, E. Pollen on-twine for food provisioning and oviposition of predatory mites in protected crops. BioControl 2014, 59, 307–317. [Google Scholar] [CrossRef]
- Aguilar-Fenollosa, E.; Jacas, J.A. Effect of ground cover management on Thysanoptera (thrips) in clementine mandarin orchards. J. Pest Sci. 2013, 86, 468–481. [Google Scholar] [CrossRef]
- Pina, T.; Argolo, P.S.; Urbaneja, A.; Jacas, J.A. Effect of pollen quality on the efficacy of two different life-style predatory mites against Tetranychus urticae in citrus. Biol. Control 2012, 61, 176–183. [Google Scholar] [CrossRef]
- Gómez-Martínez, M.A.; Aguilar-Fenollosa, E.; Jaques, J.A.; Pina, T. Ecobiology of Anaphothrips obscurus, a new dweller of citrus orchards brought in by more sustainable pest management practices. Agric. For. Entomol. 2018, 20, 93–103. [Google Scholar] [CrossRef]
- Jaques, J.A.; Aguilar-Fenollosa, E.; Hurtado, M.A.; Pina, M.T. Food web engineering to enhance biological control of Tetranychus urticae by phytoseiid mites (Tetranychidae: Phytoseiidae) in Citrus. In Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms. Progress in Biological Control; Carrillo, D., de Moraes, G.J., Peña, J.E., Eds.; Springer: Berlin, Germany, 2015; Volume 15, pp. 251–269. [Google Scholar]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agut, B.; Pastor, V.; Jaques, J.A.; Flors, V. Can Plant Defence Mechanisms Provide New Approaches for the Sustainable Control of the Two-Spotted Spider Mite Tetranychus urticae? Int. J. Mol. Sci. 2018, 19, 614. https://doi.org/10.3390/ijms19020614
Agut B, Pastor V, Jaques JA, Flors V. Can Plant Defence Mechanisms Provide New Approaches for the Sustainable Control of the Two-Spotted Spider Mite Tetranychus urticae? International Journal of Molecular Sciences. 2018; 19(2):614. https://doi.org/10.3390/ijms19020614
Chicago/Turabian StyleAgut, Blas, Victoria Pastor, Josep A. Jaques, and Victor Flors. 2018. "Can Plant Defence Mechanisms Provide New Approaches for the Sustainable Control of the Two-Spotted Spider Mite Tetranychus urticae?" International Journal of Molecular Sciences 19, no. 2: 614. https://doi.org/10.3390/ijms19020614
APA StyleAgut, B., Pastor, V., Jaques, J. A., & Flors, V. (2018). Can Plant Defence Mechanisms Provide New Approaches for the Sustainable Control of the Two-Spotted Spider Mite Tetranychus urticae? International Journal of Molecular Sciences, 19(2), 614. https://doi.org/10.3390/ijms19020614