Mesotheliomas in Genetically Engineered Mice Unravel Mechanism of Mesothelial Carcinogenesis


Abstract
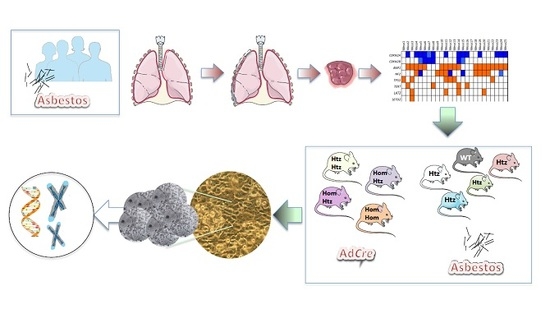
:
1. Introduction
2. Human Malignant Mesothelioma
2.1. Human Malignant Pleural Mesothelioma
2.1.1. Natural History
2.1.2. Histological Classification
2.1.3. Physiopathology
2.1.4. Molecular Alterations in MPM
2.1.5. Alterations in Regulatory Pathways
2.1.6. Molecular Classification of MPM
2.1.7. MPM Response to Treatments
2.2. Malignant Peritoneal Mesothelioma
2.3. Conclusions on Human Malignant Mesothelioma Biology
3. Models of Malignant Mesothelioma
3.1. Spontaneous Mesotheliomas in Wild Type Rodents
3.2. Mesothelioma in Animal Experiments
3.2.1. Asbestos-Induced Mesotheliomas in WT Animals
3.2.2. Mesothelioma in GEM
Spontaneous MuMM
MuMM in Mice Exposed to Carcinogenic Fibres
3.2.3. Mutation Profiles in MuMM of Mice Exposed to Fibres
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| CNT | Carbon Nanotubes |
| GEM | Genetically Engineered Mice |
| HMM | Human Malignant Mesothelioma |
| MM | Malignant Mesothelioma |
| MPeM | Malignant Peritoneal Mesothelioma |
| MuMM | Murine Malignant Mesothelioma |
References
- Boyer, A.; Pasquier, E.; Tomasini, P.; Ciccolini, J.; Greillier, L.; Andre, N.; Barlesi, F.; Mascaux, C. Drug repurposing in malignant pleural mesothelioma: A breath of fresh air? Eur. Respir. Rev. 2018, 27, 170098. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Allo, G.; John, T.; Li, M.; Tagawa, T.; Opitz, I.; Anraku, M.; Yun, Z.; Pintilie, M.; Pitcher, B.; et al. Patient-Derived Xenograft Establishment from Human Malignant Pleural Mesothelioma. Clin. Cancer Res. 2017, 23, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Pruett, N.; Hoang, C.D. In vitro experimental models of mesothelioma revisited. Transl. Lung Cancer Res. 2017, 6, 248–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzman, D.; Sterman, D.H. Updates in the diagnosis and treatment of malignant pleural mesothelioma. Curr. Opin. Pulm. Med. 2018, 24, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Lacourt, A.; Leveque, E.; Goldberg, M.; Leffondre, K. Dose-time response association between occupational asbestos exposure and pleural mesothelioma: Authors’ response. Occup. Environ. Med. 2018, 75, 161–162. [Google Scholar] [CrossRef] [PubMed]
- McCambridge, A.J.; Napolitano, A.; Mansfield, A.S.; Fennell, D.A.; Sekido, Y.; Nowak, A.K.; Reungwetwattana, T.; Mao, W.; Pass, H.I.; Carbone, M.; et al. Progress in the Management of Malignant Pleural Mesothelioma in 2017. J. Thorac. Oncol. 2018, 13, 606–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serio, G.; Pagliarule, V.; Marzullo, A.; Pezzuto, F.; Gentile, M.; Pennella, A.; Nazzaro, P.; Buonadonna, A.L.; Covelli, C.; Lettini, T.; et al. Molecular changes of malignant mesothelioma in the testis and their impact on prognosis: Analyses of two cases. Int. J. Clin. Exp. Pathol. 2016, 9, 7658–7667. [Google Scholar]
- Goswami, E.; Craven, V.; Dahlstrom, D.L.; Alexander, D.; Mowat, F. Domestic asbestos exposure: A review of epidemiologic and exposure data. Int. J. Environ. Res. Public Health 2013, 10, 5629–5670. [Google Scholar] [CrossRef] [PubMed]
- Noonan, C.W. Environmental asbestos exposure and risk of mesothelioma. Ann. Transl. Med. 2017, 5, 234. [Google Scholar] [CrossRef] [PubMed]
- Serio, G.; Pezzuto, F.; Marzullo, A.; Scattone, A.; Cavone, D.; Punzi, A.; Fortarezza, F.; Gentile, M.; Buonadonna, A.L.; Barbareschi, M.; et al. Peritoneal Mesothelioma with Residential Asbestos Exposure. Report of a Case with Long Survival (Seventeen Years) Analyzed by Cgh-Array. Int. J. Mol. Sci. 2017, 18, 1818. [Google Scholar] [CrossRef] [PubMed]
- Vimercati, L.; Cavone, D.; Lovreglio, P.; De Maria, L.; Caputi, A.; Ferri, G.M.; Serio, G. Environmental asbestos exposure and mesothelioma cases in Bari, Apulia region, southern Italy: A national interest site for land reclamation. Environ. Sci. Pollut. 2018, 25, 15692–15701. [Google Scholar] [CrossRef] [PubMed]
- Husain, A.N.; Colby, T.V.; Ordonez, N.G.; Allen, T.C.; Attanoos, R.L.; Beasley, M.B.; Butnor, K.J.; Chirieac, L.R.; Churg, A.M.; Dacic, S.; et al. Guidelines for Pathologic Diagnosis of Malignant Mesothelioma 2017 Update of the Consensus Statement From the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2018, 142, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Mutsaers, S.E.; Jaurand, M.C.; GaryLee, Y.C.; Prele, C.M. Mesothelial cells and pleural immunology. In Textbook of Pleural Diseases, 3nd ed.; Light, R.W., GaryLee, Y.C., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 27–44. [Google Scholar]
- Wang, N.S. Anatomy of the pleura. Clin. Chest Med. 1998, 19, 229–240. [Google Scholar] [CrossRef]
- Oshiro, H.; Miura, M.; Iobe, H.; Kudo, T.; Shimazu, Y.; Aoba, T.; Okudela, K.; Nagahama, K.; Sakamaki, K.; Yoshida, M.; et al. Lymphatic Stomata in the Adult Human Pulmonary Ligament. Lymphat. Res. Biol. 2015, 13, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broaddus, V.C.; Everitt, J.I.; Black, B.; Kane, A.B. Non-neoplastic and neoplastic pleural endpoints following fiber exposure. J. Toxicol. Environ. Health B Crit. Rev. 2011, 14, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Fleury Feith, J.; Jaurand, M.C. Pleural lymphatics and pleural diseases related to fibres. Rev. Pneumol. Clin. 2013, 69, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Oehl, K.; Vrugt, B.; Opitz, I.; Meerang, M. Heterogeneity in Malignant Pleural Mesothelioma. Int. J. Mol. Sci. 2018, 19, 1603. [Google Scholar] [CrossRef] [PubMed]
- Hylebos, M.; Van Camp, G.; Vandeweyer, G.; Fransen, E.; Beyens, M.; Cornelissen, R.; Suls, A.; Pauwels, P.; van Meerbeeck, J.P.; Op de Beeck, K. Large-scale copy number analysis reveals variations in genes not previously associated with malignant pleural mesothelioma. Oncotarget 2017, 8, 113673–113686. [Google Scholar] [CrossRef] [PubMed]
- Jean, D.; Daubriac, J.; Le Pimpec-Barthes, F.; Galateau-Salle, F.; Jaurand, M.C. Molecular changes in mesothelioma with an impact on prognosis and treatment. Arch. Pathol. Lab. Med. 2012, 136, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Tranchant, R.; Quetel, L.; Tallet, A.; Meiller, C.; Renier, A.; de Koning, L.; de Reynies, A.; Le Pimpec-Barthes, F.; Zucman-Rossi, J.; Jaurand, M.C.; et al. Co-occurring Mutations of Tumor Suppressor Genes, LATS2 and NF2, in Malignant Pleural Mesothelioma. Clin. Cancer Res. 2017, 23, 3191–3202. [Google Scholar] [CrossRef] [PubMed]
- Bueno, R.; Stawiski, E.W.; Goldstein, L.D.; Durinck, S.; De Rienzo, A.; Modrusan, Z.; Gnad, F.; Nguyen, T.T.; Jaiswal, B.S.; Chirieac, L.R.; et al. Comprehensive genomic analysis of malignant pleural mesothelioma identifies recurrent mutations, gene fusions and splicing alterations. Nat. Genet. 2016, 48, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Jean, D.; Thomas, E.; Renier, A.; de Reynies, A.; Lecomte, C.; Andujar, P.; Fleury-Feith, J.; Giovannini, M.; Zucman-Rossi, J.; Stern, M.H.; et al. Syntenic relationships between genomic profiles of fiber-induced murine and human malignant mesothelioma. Am. J. Pathol. 2011, 176, 881–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borczuk, A.C.; Pei, J.; Taub, R.N.; Levy, B.; Nahum, O.; Chen, J.; Chen, K.; Testa, J.R. Genome-wide analysis of abdominal and pleural malignant mesothelioma with DNA arrays reveals both common and distinct regions of copy number alteration. Cancer Biol. Ther. 2016, 17, 328–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tallet, A.; Nault, J.C.; Renier, A.; Hysi, I.; Galateau-Salle, F.; Cazes, A.; Copin, M.C.; Hofman, P.; Andujar, P.; Le Pimpec-Barthes, F.; et al. Overexpression and promoter mutation of the TERT gene in malignant pleural mesothelioma. Oncogene 2014, 33, 3748. [Google Scholar] [CrossRef] [PubMed]
- Barone, E.; Gemignani, F.; Landi, S. Overexpressed genes in malignant pleural mesothelioma: Implications in clinical management. J. Thorac. Dis. 2018, 10, S369–S382. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, E.; Suraokar, M.B.; Rodriguez, J.; Mino, B.; Lin, H.Y.; Rice, D.C.; Tsao, A.; Wistuba, I.I. Frequent coamplification and cooperation between C-MYC and PVT1 oncogenes promote malignant pleural mesothelioma. J. Thorac. Oncol. 2014, 9, 998–1007. [Google Scholar] [CrossRef] [PubMed]
- Tsao, A.S.; Harun, N.; Fujimoto, J.; Devito, V.; Lee, J.J.; Kuhn, E.; Mehran, R.; Rice, D.; Moran, C.; Hong, W.K.; et al. Elevated PDGFRB gene copy number gain is prognostic for improved survival outcomes in resected malignant pleural mesothelioma. Ann. Diagn. Pathol. 2014, 18, 140–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, L.; Finn, S.P.; Cuffe, S.; Gray, S.G. Non-coding RNA repertoires in malignant pleural mesothelioma. Lung Cancer 2015, 90, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Vannini, I.; Fanini, F.; Fabbri, M. Emerging roles of microRNAs in cancer. Curr. Opin. Genet. Dev. 2018, 48, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.C.; Houseman, E.A.; Godleski, J.J.; Marsit, C.J.; Longacker, J.L.; Roelofs, C.R.; Karagas, M.R.; Wrensch, M.R.; Yeh, R.F.; Nelson, H.H.; et al. Epigenetic profiles distinguish pleural mesothelioma from normal pleura and predict lung asbestos burden and clinical outcome. Cancer Res. 2009, 69, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.C.; Godleski, J.J.; Marsit, C.J.; Houseman, E.A.; Lopez-Fagundo, C.Y.; Longacker, J.L.; Bueno, R.; Sugarbaker, D.J.; Nelson, H.H.; Kelsey, K.T. Asbestos exposure predicts cell cycle control gene promoter methylation in pleural mesothelioma. Carcinogenesis 2008, 29, 1555–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, R.; Ali, G.; Giannini, R.; Proietti, A.; Lucchi, M.; Chella, A.; Melfi, F.; Mussi, A.; Fontanini, G. Malignant pleural mesothelioma and mesothelial hyperplasia: A new molecular tool for the differential diagnosis. Oncotarget 2017, 8, 2758–2770. [Google Scholar] [CrossRef] [PubMed]
- Busacca, S.; Germano, S.; De Cecco, L.; Rinaldi, M.; Comoglio, F.; Favero, F.; Murer, B.; Mutti, L.; Pierotti, M.; Gaudino, G. MicroRNA signature of malignant mesothelioma with potential diagnostic and prognostic implications. Am. J. Respir. Cell Mol. Biol. 2010, 42, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Reid, G. MicroRNAs in mesothelioma: From tumour suppressors and biomarkers to therapeutic targets. J. Thorac. Dis. 2015, 7, 1031–1040. [Google Scholar] [PubMed]
- De Santi, C.; Melaiu, O.; Bonotti, A.; Cascione, L.; Di Leva, G.; Foddis, R.; Cristaudo, A.; Lucchi, M.; Mora, M.; Truini, A.; et al. Deregulation of miRNAs in malignant pleural mesothelioma is associated with prognosis and suggests an alteration of cell metabolism. Sci. Rep. 2017, 7, 3140. [Google Scholar] [CrossRef] [PubMed]
- Jaurand, M.C.; Jean, D. Biomolecular Pathways and Malignant Pleural Mesothelioma. In Malignant Pleural Mesothelioma: Present Status and Future Directions; Mineo, T.C., Ed.; Bentham Science Publishers: Sharjah, UAE, 2015; pp. 173–196. [Google Scholar]
- Sekido, Y. Molecular pathogenesis of malignant mesothelioma. Carcinogenesis 2013, 34, 1413–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felley-Bosco, E.; Opitz, I.; Meerang, M. Hedgehog Signaling in Malignant Pleural Mesothelioma. Genes 2015, 6, 500–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gradilla, A.C.; Sanchez-Hernandez, D.; Brunt, L.; Scholpp, S. From top to bottom: Cell polarity in Hedgehog and Wnt trafficking. BMC Biol. 2018, 16, 37. [Google Scholar] [CrossRef] [PubMed]
- Leprieur, E.G.; Jablons, D.M.; He, B. Old Sonic Hedgehog, new tricks: A new paradigm in thoracic malignancies. Oncotarget 2018, 9, 14680–14691. [Google Scholar] [CrossRef] [PubMed]
- Suraokar, M.B.; Nunez, M.I.; Diao, L.; Chow, C.W.; Kim, D.; Behrens, C.; Lin, H.; Lee, S.; Raso, G.; Moran, C.; et al. Expression profiling stratifies mesothelioma tumors and signifies deregulation of spindle checkpoint pathway and microtubule network with therapeutic implications. Ann. Oncol. 2014, 25, 1184–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Reynies, A.; Jaurand, M.C.; Renier, A.; Couchy, G.; Hysi, I.; Elarouci, N.; Galateau-Salle, F.; Copin, M.C.; Hofman, P.; Cazes, A.; et al. Molecular classification of malignant pleural mesothelioma: Identification of a poor prognosis subgroup linked to the epithelial-to-mesenchymal transition. Clin. Cancer Res. 2014, 20, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Churg, A.; Attanoos, R.; Borczuk, A.C.; Chirieac, L.R.; Galateau-Salle, F.; Gibbs, A.; Henderson, D.; Roggli, V.; Rusch, V.; Judge, M.J.; et al. Dataset for Reporting of Malignant Mesothelioma of the Pleura or Peritoneum: Recommendations From the International Collaboration on Cancer Reporting (ICCR). Arch. Pathol. Lab. Med. 2016, 140, 1104–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridda, A.; Padoan, I.; Mencarelli, R.; Frego, M. Peritoneal mesothelioma: A review. MedGenMed 2007, 9, 32. [Google Scholar] [PubMed]
- Joseph, N.M.; Chen, Y.Y.; Nasr, A.; Yeh, I.; Talevich, E.; Onodera, C.; Bastian, B.C.; Rabban, J.T.; Garg, K.; Zaloudek, C.; et al. Genomic profiling of malignant peritoneal mesothelioma reveals recurrent alterations in epigenetic regulatory genes BAP1, SETD2, and DDX3X. Mod. Pathol. 2017, 30, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.P.; Dong, F.; Watkins, J.C.; Nardi, V.; Bueno, R.; Dal Cin, P.; Godleski, J.J.; Crum, C.P.; Chirieac, L.R. Identification of ALK Rearrangements in Malignant Peritoneal Mesothelioma. JAMA Oncol. 2018, 4, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Tanigawa, H.; Onodera, H.; Maekawa, A. Spontaneous Mesotheliomas in Fischer Rats—A Histological and Electron Microscopic Study. Toxicol. Pathol. 1987, 15, 157–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maronpot, R.R.; Nyska, A.; Foreman, J.E.; Ramot, Y. The legacy of the F344 rat as a cancer bioassay model (a retrospective summary of three common F344 rat neoplasms). Crit. Rev. Toxicol. 2016, 46, 641–675. [Google Scholar] [CrossRef] [PubMed]
- Bomhard, E. Frequency of spontaneous tumours in NMRI mice in 21-month studies. Exp. Toxicol. Pathol. 1993, 45, 269–289. [Google Scholar] [CrossRef]
- Peters, R.L.; Rabstein, L.S.; Spahn, G.J.; Madison, R.M.; Huebner, R.J. Incidence of spontaneous neoplasms in breeding and retired breeder BALB-cCr mice throughout the natural life span. Int. J. Cancer 1972, 10, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Shirai, M.; Maejima, T.; Tanimoto, T.; Kumagai, K.; Makino, T.; Kai, K.; Teranishi, M.; Sanbuissho, A. Mixed Type of Malignant Mesothelioma in an Aged Male ICR Mouse. J. Toxicol. Pathol. 2011, 24, 169–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IARC. Man-made vitreous fibres and Radon. IARC Monogr. Eval. Carcinog. Risks Hum. 1988, 43, 1–300. [Google Scholar]
- Boulanger, G.; Andujar, P.; Pairon, J.C.; Billon-Galland, M.A.; Dion, C.; Dumortier, P.; Brochard, P.; Sobaszek, A.; Bartsch, P.; Paris, C.; et al. Quantification of short and long asbestos fibers to assess asbestos exposure: A review of fiber size toxicity. Environ. Health 2014, 13, 59. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.C.; Berry, G. Mesotheliomas in rats following inoculation with asbestos. Br. J. Cancer 1969, 23, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, S.; Patch, A.M.; Dick, I.M.; Kazakoff, S.; Pearson, J.V.; Waddell, N.; Allcock, R.J.N.; Holt, R.A.; Robinson, B.W.S.; Creaney, J. Whole exome sequencing of an asbestos-induced wild-type murine model of malignant mesothelioma. BMC Cancer 2017, 17, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernova, T.; Murphy, F.A.; Galavotti, S.; Sun, X.M.; Powley, I.R.; Grosso, S.; Schinwald, A.; Zacarias-Cabeza, J.; Dudek, K.M.; Dinsdale, D.; et al. Long-Fiber Carbon Nanotubes Replicate Asbestos-Induced Mesothelioma with Disruption of the Tumor Suppressor Gene Cdkn2a (Ink4a/Arf). Curr. Biol. 2017, 27, 3302–3314.e6. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chirieac, L.R.; Bueno, R.; Pass, H.; Wu, W.; Malinowska, I.A.; Kwiatkowski, D.J. Tsc1-Tp53 loss induces mesothelioma in mice, and evidence for this mechanism in human mesothelioma. Oncogene 2014, 33, 3151–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongsma, J.; van Montfort, E.; Vooijs, M.; Zevenhoven, J.; Krimpenfort, P.; van der Valk, M.; van de Vijver, M.; Berns, A. A conditional mouse model for malignant mesothelioma. Cancer Cell 2008, 13, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Kadariya, Y.; Cheung, M.; Xu, J.; Pei, J.; Sementino, E.; Menges, C.W.; Cai, K.Q.; Rauscher, F.J.; Klein-Szanto, A.J.; Testa, J.R. Bap1 Is a Bona Fide Tumor Suppressor: Genetic Evidence from Mouse Models Carrying Heterozygous Germline Bap1 Mutations. Cancer Res. 2016, 76, 2836–2844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sementino, E.; Menges, C.W.; Kadariya, Y.; Peri, S.; Xu, J.; Liu, Z.; Wilkes, R.G.; Cai, K.Q.; Rauscher, F.J., 3rd; Klein-Szanto, A.J.; et al. Inactivation of Tp53 and Pten drives rapid development of pleural and peritoneal malignant mesotheliomas. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Altomare, D.A.; Menges, C.W.; Pei, J.; Zhang, L.; Skele-Stump, K.L.; Carbone, M.; Kane, A.B.; Testa, J.R. Activated TNF-α/NF-κB signaling via down-regulation of Fas-associated factor 1 in asbestos-induced mesotheliomas from Arf knockout mice. Proc. Natl. Acad. Sci. USA 2009, 106, 3420–3425. [Google Scholar] [CrossRef] [PubMed]
- Altomare, D.A.; Menges, C.W.; Xu, J.; Pei, J.; Zhang, L.; Tadevosyan, A.; Neumann-Domer, E.; Liu, Z.; Carbone, M.; Chudoba, I.; et al. Losses of both products of the Cdkn2a/Arf locus contribute to asbestos-induced mesothelioma development and cooperate to accelerate tumorigenesis. PLoS ONE 2011, 6, e18828. [Google Scholar] [CrossRef] [PubMed]
- Altomare, D.A.; Vaslet, C.A.; Skele, K.L.; De Rienzo, A.; Devarajan, K.; Jhanwar, S.C.; McClatchey, A.I.; Kane, A.B.; Testa, J.R. A mouse model recapitulating molecular features of human mesothelioma. Cancer Res. 2005, 65, 8090–8095. [Google Scholar] [CrossRef] [PubMed]
- Andujar, P.; Lecomte, C.; Renier, A.; Fleury-Feith, J.; Kheuang, L.; Daubriac, J.; Janin, A.; Jaurand, M.C. Clinico-pathological features and somatic gene alterations in refractory ceramic fibre-induced murine mesothelioma reveal mineral fibre-induced mesothelioma identities. Carcinogenesis 2007, 28, 1599–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleury-Feith, J.; Lecomte, C.; Renier, A.; Matrat, M.; Kheuang, L.; Abramowski, V.; Levy, F.; Janin, A.; Giovannini, M.; Jaurand, M.C. Hemizygosity of Nƒ2 is associated with increased susceptibility to asbestos-induced peritoneal tumours. Oncogene 2003, 22, 3799–3805. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, A.; Pellegrini, L.; Dey, A.; Larson, D.; Tanji, M.; Flores, E.G.; Kendrick, B.; Lapid, D.; Powers, A.; Kanodia, S.; et al. Minimal asbestos exposure in germline BAP1 heterozygous mice is associated with deregulated inflammatory response and increased risk of mesothelioma. Oncogene 2016, 35, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Vaslet, C.A.; Messier, N.J.; Kane, A.B. Accelerated progression of asbestos-induced mesotheliomas in heterozygous p53+/− mice. Toxicol. Sci. 2002, 68, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Menges, C.W.; Kadariya, Y.; Altomare, D.; Talarchek, J.; Neumann-Domer, E.; Wu, Y.; Xiao, G.H.; Shapiro, I.M.; Kolev, V.N.; Pachter, J.A.; et al. Tumor suppressor alterations cooperate to drive aggressive mesotheliomas with enriched cancer stem cells via a p53-miR-34a-c-Met axis. Cancer Res. 2014, 74, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Takagi, A.; Hirose, A.; Nishimura, T.; Fukumori, N.; Ogata, A.; Ohashi, N.; Kitajima, S.; Kanno, J. Induction of mesothelioma in p53+/− mouse by intraperitoneal application of multi-wall carbon nanotube. J. Toxicol. Sci. 2008, 33, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Kadariya, Y.; Menges, C.W.; Talarchek, J.; Cai, K.Q.; Klein-Szanto, A.J.; Pietrofesa, R.A.; Christofidou-Solomidou, M.; Cheung, M.; Mossman, B.T.; Shukla, A.; et al. Inflammation-Related IL1beta/IL1R Signaling Promotes the Development of Asbestos-Induced Malignant Mesothelioma. Cancer Prev. Res. 2016, 9, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.; van Bruggen, I.; Segal, A.; Dunham, M.; Sherwood, A.; Koentgen, F.; Robinson, B.W.; Lake, R.A. A novel SV40 TAg transgenic model of asbestos-induced mesothelioma: Malignant transformation is dose dependent. Cancer Res. 2006, 66, 10786–10794. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.; Dick, I.M.; Wise, M.J.; Holloway, A.; Diyagama, D.; Robinson, B.W.; Creaney, J.; Lake, R.A. Consistent gene expression profiles in MexTAg transgenic mouse and wild type mouse asbestos-induced mesothelioma. BMC Cancer 2015, 15, 983. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, C.; Andujar, P.; Renier, A.; Kheuang, L.; Abramowski, V.; Mellottee, L.; Fleury-Feith, J.; Zucman-Rossi, J.; Giovannini, M.; Jaurand, M.C. Similar tumor suppressor gene alteration profiles in asbestos-induced murine and human mesothelioma. Cell Cycle 2005, 4, 1862–1869. [Google Scholar] [CrossRef] [PubMed]
- Rehrauer, H.; Wu, L.; Blum, W.; Pecze, L.; Henzi, T.; Serre-Beinier, V.; Aquino, C.; Vrugt, B.; de Perrot, M.; Schwaller, B.; et al. How asbestos drives the tissue towards tumors: YAP activation, macrophage and mesothelial precursor recruitment, RNA editing, and somatic mutations. Oncogene 2018, 37, 2645–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Kadariya, Y.; Cheung, M.; Pei, J.; Talarchek, J.; Sementino, E.; Tan, Y.; Menges, C.W.; Cai, K.Q.; Litwin, S.; et al. Germline mutation of Bap1 accelerates development of asbestos-induced malignant mesothelioma. Cancer Res. 2014, 74, 4388–4397. [Google Scholar] [CrossRef] [PubMed]
- Bignell, G.R.; Greenman, C.D.; Davies, H.; Butler, A.P.; Edkins, S.; Andrews, J.M.; Buck, G.; Chen, L.; Beare, D.; Latimer, C.; et al. Signatures of mutation and selection in the cancer genome. Nature 2010, 463, 893–898. [Google Scholar] [CrossRef] [PubMed]
- Moller, P.; Danielsen, P.H.; Jantzen, K.; Roursgaard, M.; Loft, S. Oxidatively damaged DNA in animals exposed to particles. Crit. Rev. Toxicol. 2013, 43, 96–118. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Gene(s) Affected | Gene(s) Status | MuMM % | Epi 2 % | Sarco 2 % | Mixed % | Survival 3 Weeks | Reference |
|---|---|---|---|---|---|---|---|
| Nf2 Ink4a/Arf | Htz Hom | 34 4 | 28.6 | 21.4 | 50 | 58 5 | [59] |
| Nf2 Ink4a/Arf | Hom Htz | 34.6 4 | 22.2 | 27.8 | 50 | 71 5 | [59] |
| Nf2 Ink4a/Arf | Hom Hom | 79 4 | 2.2 | 68.9 | 28.9 | 31 5 | [59] |
| Nf2 Rb | Htz Hom | 5.9 4 | 0 | Primarily sarco | Some mixed | ND 7 | [59] |
| Nf2 Rb | Hom Htz | 13.3 4 | 0 | Primarily sarco | Some mixed | ND | [59] |
| Nf2 Rb | Hom Hom | 26.3 4 | 0 | Primarily sarco | Some mixed | ND | [59] |
| Nf2 Rb | Htz Hom | 6.75 6 | 0 | Primarily sarco | Some mixed | ND | [59] |
| Nf2 Rb | Hom Htz | 13.3 6 | 0 | Primarily sarco | Some mixed | ND | [59] |
| Nf2 Rb | Hom Hom | 20 6 | 0 | Primarily sarco | Some mixed | ND | [59] |
| Nf2 Trp53 | Htz Hom | 59 4 | 25 | 25 | 50 | 29 5 | [59] |
| Nf2 Trp53 | Hom Htz | 25 4 | 60 | 40 | 0 | 86 5 | [59] |
| Nf2 Trp53 | Hom Hom | 82 4 | 15.5 | 46.7 | 37.8 | 19 5 | [59] |
| Nf2 Trp53 Ink4a | Hom Hom Htz | 93.7 4 | 0 | 40 | 60 | ND | [59] |
| Nf2 Trp53 Ink4a | Hom Hom Hom | 91.1 4 | 0 | 76.6 | 23.4 | 11 | [59] |
| Tsc1 Tp53 | Hom Hom | 85 6 | Mostly | 37 | [58] | ||
| Tsc1 Tp53 | Hom WT | 0 6 | NA 7 | >57 | [58] | ||
| Tsc1 Tp53 | WT Hom | 0 6 | NA | >57 | [58] | ||
| Tsc1 Tp53 | WT WT | 0 6 | NA | >57 | [58] | ||
| Tsc1 Tp53 | Hom Hom | 73 8 | Mostly | 44 | [58] | ||
| Tsc1 Tp53 | Hom WT | 0 8 | NA | >57 | [58] | ||
| Tsc1 Tp53 | WT Hom | 0 8 | NA | >57 | [58] | ||
| Tsc1 Tp53 | WT WT | 0 8 | NA | >57 | [58] |
| Mice Strain | Gene(s) Affected | Gene(s) Status | Treatment | MuMM % | Epi 2 % | Sarco 2 % | Mixed % | Survival 3 Weeks | Reference |
|---|---|---|---|---|---|---|---|---|---|
| FVB/N | Nf2 | Htz | Asbestos | 47 | 30.4 Htz + WT | 65.2 Htz + WT | 4.3 Htz + WT | [66] | |
| FVB/N | None | WT | Asbestos | 15 | 30.4 Htz + WT | 65.2 Htz + WT | 4.3 Htz + WT | [66] | |
| FVB/N | Nf2 | Htz | Saline | 0 | NA | NA | NA | [66] | |
| FVB/N | None | WT | Saline | 0 | NA | NA | NA | [66] | |
| 129Sv/Jae | Nf2 | Htz | Asbestos | 85 | 6.25 | 18.75 | 75 | 43 | [64] |
| 129Sv/Jae | None | WT | Asbestos | 59 | 31 | 27.6 | 41.4 | 52 | [64] |
| FVB/N | Nf2 | Htz | RCF | 55 | 27 | 38.4 | 34.6 | 68 | [65] |
| FVB/N | None | WT | RCF | 7.1 | 0 | 0 | 100 | 80 | [65] |
| C57/Bl6 | Nf2 | Htz | Asbestos | 10 | ND | ND | ND | ND | [75] |
| C57/Bl6 | Arf | Htz | Asbestos | 96.2 | 68 | 12 | 20 | 42 | [62] |
| C57/Bl6 | None | WT | Asbestos | 81.5 | 68.2 | 18.2 | 13.6 | 56 | [62] |
| Hybrids | Ink4a/Arf | Htz | Asbestos | 88 | Occasional | Prevalent | Occasional | 29.6 | [63] |
| Hybrids | Ink4a/Arf | Htz | TiO2 | 0 | NA | NA | NA | NA | [63] |
| Hybrids | Ink4a | Htz | Asbestos | 66 | Occasional | Prevalent | Occasional | 34.6 | [63] |
| Hybrids | Arf | Htz | Asbestos | 65 | Occasional | Prevalent | Occasional | 38 | [63] |
| Hybrids | None | WT | Asbestos | 50 | Occasional | Prevalent | Occasional | 49.4 | [63] |
| Hybrids | Nf2 | Htz | Asbestos | ND | ND | ND | ND | 38 | [69] |
| Hybrids | Nf2Ink4a/Arf | HtzHtz | Asbestos | ND | ND | Most sarcomatous | ND | 24 | [69] |
| Hybrids | None | WT | Asbestos | ND | ND | ND | ND | 45 | [69] |
| 129/Sv on a 75% C57/Bl6 background | Trp53 | Htz | Asbestos | 76 (after 44 weeks) | ND | ND | ND | [68] | |
| 129/Sv on a 75% C57/Bl6 background | Trp53 | Hom | Asbestos | ND | ND | ND | ND | [68] | |
| 129/Sv on a 75% C57/Bl6 background | None | WT | Asbestos | 32 (after 67 weeks) | ND | ND | ND | [68] | |
| FVB | Bap1 | Htz | Asbestos | 73 | ND | ND | ND | 43 | [76] |
| FVB | None | WT | Asbestos | 32 | ND | ND | ND | 55 | [76] |
| FVB | Bap1 | Htz (L) | Asbestos | 71 | ND | ND | ND | 46 | [60] |
| FVB | Bap1 | Htz (W) | Asbestos | 74 | ND | ND | ND | 48 | [60] |
| FVB | None | WT | Asbestos | 35 | ND | ND | ND | 60 | [60] |
| C57BL/6 | Bap1 | Htz | Asbestos low dose | 36 | all or part | 57 | [67] | ||
| C57BL/6 | None | WT | Asbestos low dose | 10 | all or part | 57 | [67] | ||
| C57BL/6 | None | WT | Saline | 0 | NA | NA | [67] | ||
| C57BL/6 | Bap1 | Htz | Asbestos std dose | 60 | all or part | 39 | [67] | ||
| C57BL/6 | Bap1 | WT | Asbestos std dose | 28 | all or part | 57 | [67] | ||
| C57BL/6 | Asc | Hom | Asbestos | 55 | 0 | 75 | 25 | 66.2 | [71] |
| C57BL/6 | Asc | Htz | Asbestos | 65 | 0 | 68 | 32 | 69.4 | [71] |
| C57BL/6 | None | WT | Asbestos | 80 | 0 | 67 | 33 | OK | [71] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jean, D.; Jaurand, M.-C. Mesotheliomas in Genetically Engineered Mice Unravel Mechanism of Mesothelial Carcinogenesis. Int. J. Mol. Sci. 2018, 19, 2191. https://doi.org/10.3390/ijms19082191
Jean D, Jaurand M-C. Mesotheliomas in Genetically Engineered Mice Unravel Mechanism of Mesothelial Carcinogenesis. International Journal of Molecular Sciences. 2018; 19(8):2191. https://doi.org/10.3390/ijms19082191
Chicago/Turabian StyleJean, Didier, and Marie-Claude Jaurand. 2018. "Mesotheliomas in Genetically Engineered Mice Unravel Mechanism of Mesothelial Carcinogenesis" International Journal of Molecular Sciences 19, no. 8: 2191. https://doi.org/10.3390/ijms19082191
APA StyleJean, D., & Jaurand, M. -C. (2018). Mesotheliomas in Genetically Engineered Mice Unravel Mechanism of Mesothelial Carcinogenesis. International Journal of Molecular Sciences, 19(8), 2191. https://doi.org/10.3390/ijms19082191