Omics Approaches for Engineering Wheat Production under Abiotic Stresses


Abstract
:1. Introduction
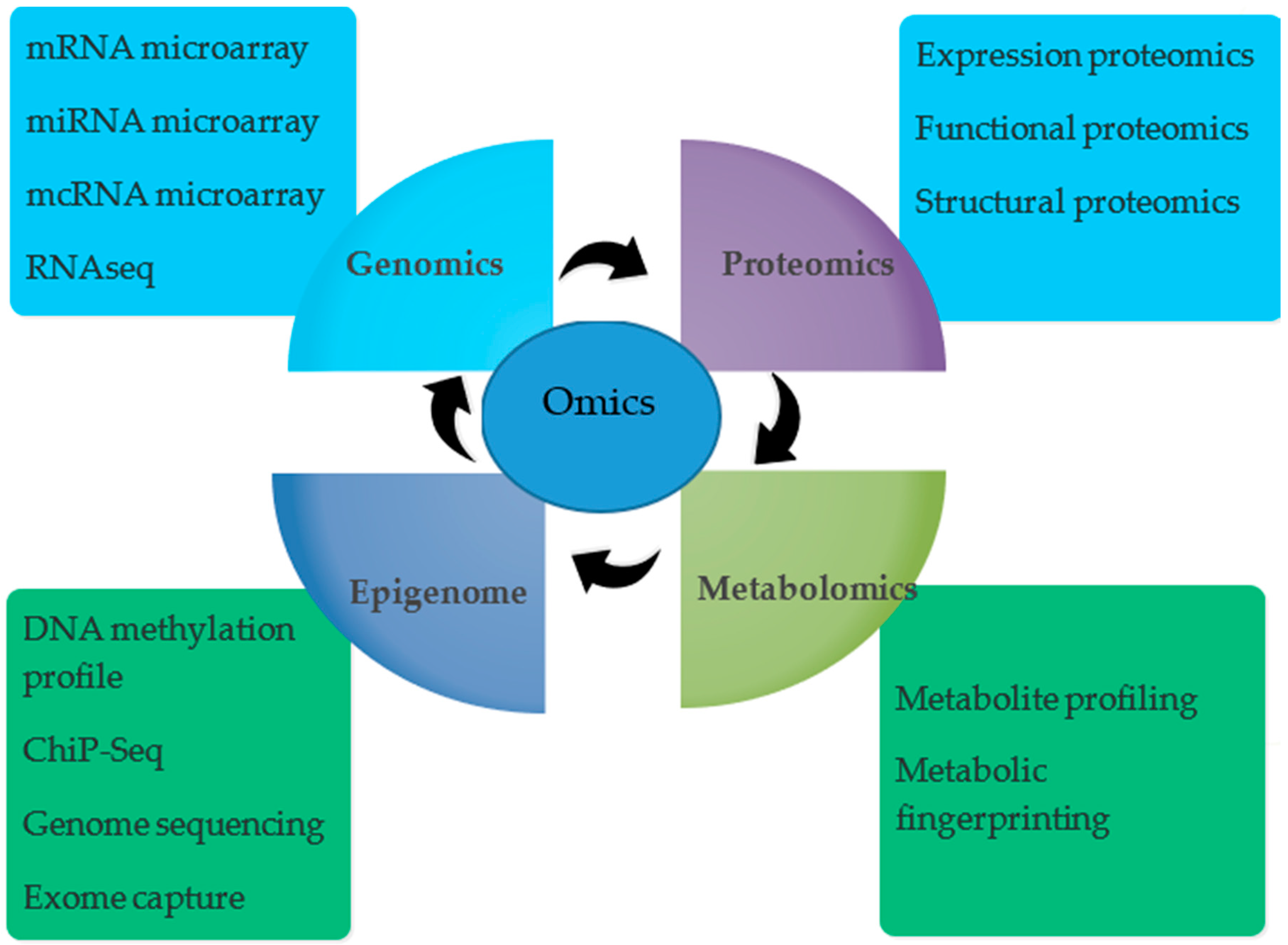
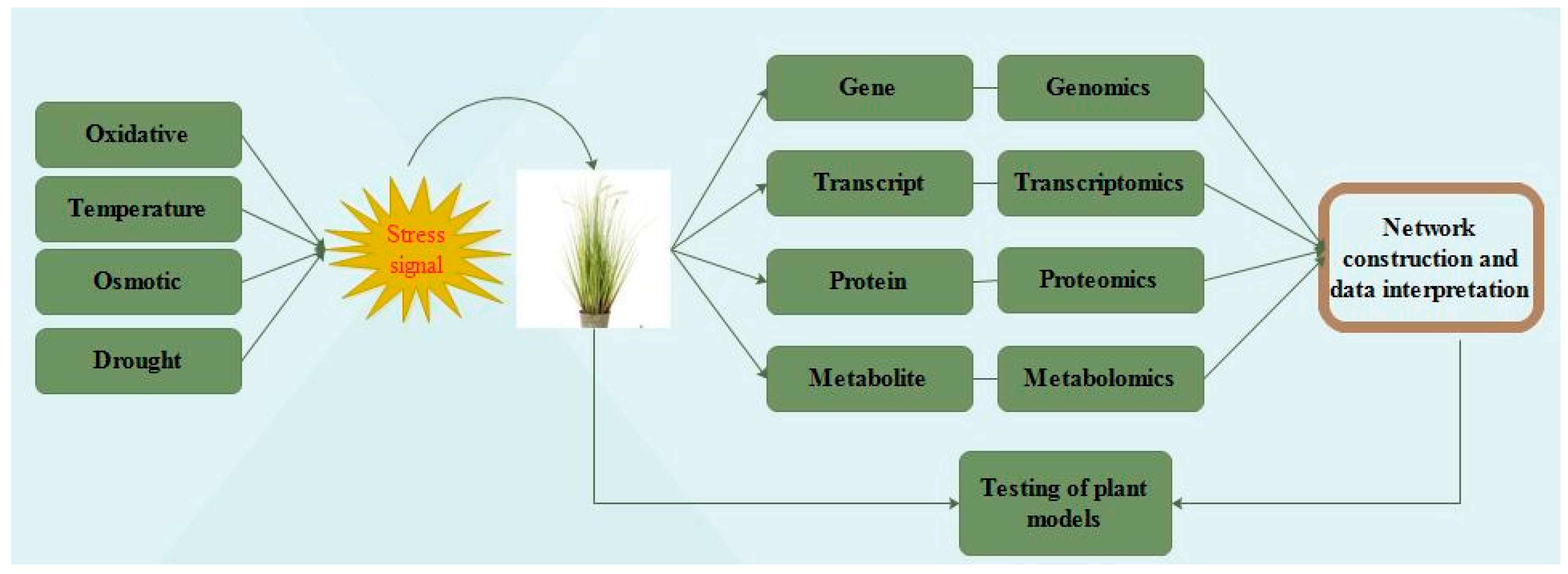
2. Omics Approaches in the Technological Era
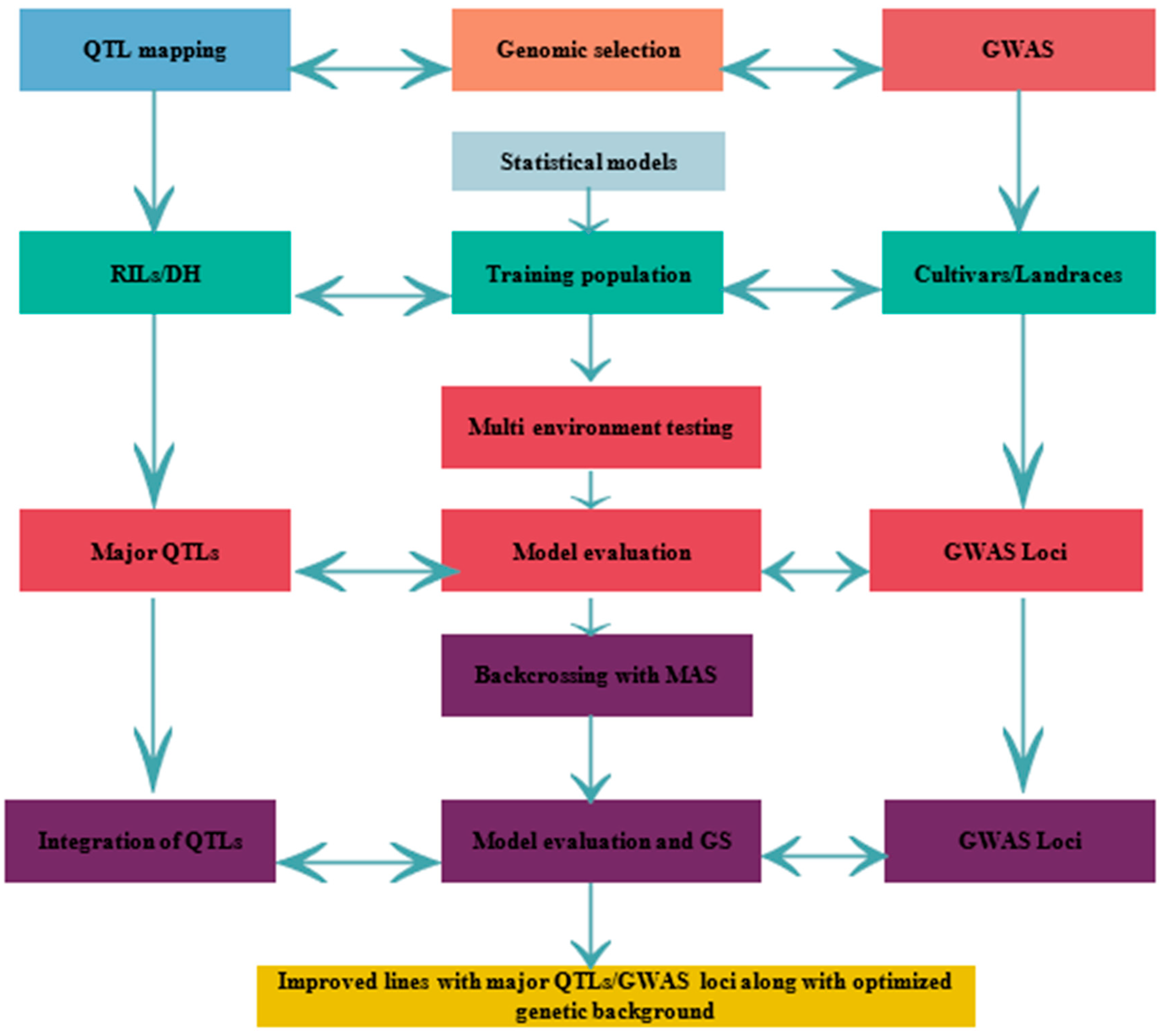
3. Genomics Progresses for Abiotic Stress Tolerance in Wheat
3.1. Molecular Marker Resources
3.2. Quantitative Trait Loci (QTL) Mapping for Abiotic Stress
3.3. Genome Wide Association Studies
3.4. Genomic Selection
3.5. Transcriptome Profiling for Abiotic Stress Tolerance
4. Proteomics in Wheat
5. Metabolomics Advances for Abiotic Stress
6. Ionomics for Wheat
7. Phenomics Prospective in Wheat
8. Role of Online Databases for Effective Integration of Omics Platforms
9. Conclusions and Future Perspectives
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Leonard, W.H.; Martin, J.H. Cereal Crop; No. SB189 L37; The Macmillan Company: New York, NY, USA, 1963. [Google Scholar]
- Santra, D.; Santra, M.; Allan, R.; Campbell, K.; Kidwell, K. Genetic and molecular characterization of vernalization genes Vrn-A1, Vrn-B1 and Vrn-D1 in spring wheat germplasm from the Pacific Northwest region of the U.S.A. Plant Breed. 2009, 28, 576–584. [Google Scholar] [CrossRef]
- Gomez, D.; Vanzetti, L.; Helguera, M.; Lombardo, L.; Fraschina, J.; Miralles, D.J. Effect of Vrn-1, Ppd-1 genes and earliness per se on heading time in Argentinean bread wheat cultivars. Field Crop. Res. 2014, 158, 73–81. [Google Scholar] [CrossRef]
- Goncharov, N.P. Genetics of growth habit (spring vs. winter) in common wheat: Confirmation of the existence of dominant gene Vrn4. Theor. Appl. Genet. 2003, 107, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Grainger, C.M.; Rajcan, I. Characterization of the genetic changes in a multi-generational pedigree of an elite Canadian soybean cultivar. Theor. Appl. Genet. 2014, 127, 211–229. [Google Scholar] [CrossRef] [PubMed]
- Manavalan, L.P.; Guttikonda, S.K.; Phan Tran, L.S.; Nguyen, H.T. Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, X.; Ge, S.; Jensen, J.D.; Hu, F.; Li, X.; Dong, Y.; Gutenkunst, R.N.; Fang, L.; Huang, L.; et al. Resequencing 50 accessions of cultivated and wild rice yields markers for identifying agronomically important genes. Nat. Biotechnol. 2012, 30, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Tomar, R.S.S.; Deshmukh, R.K.; Naik, K.; Tomar, S.M.S.; Vinod. Development of chloroplast−specific microsatellite markers for molecular characterization of alloplasmic lines and phylogenetic analysis in wheat. Plant Breed. 2014, 133, 12–18. [Google Scholar] [CrossRef]
- Song, Q.; Jia, G.; Zhu, Y.; Grant, D.; Nelson, R.T.; Hwang, E.Y.; Hythen, D.L.; Cregan, P.B. Abundance of SSR motifs and development of candidate polymorphic SSR markers (BARCSOYSSR_1. 0) in soybean. Crop Sci. 2010, 50, 1950–1960. [Google Scholar] [CrossRef]
- Tardivel, A.; Sonah, H.; Belzile, F.; O’Donoughue, L.S. Rapid identification of alleles at the soybean maturity gene E3 using genotyping by sequencing and a haplotype-based approach. Plant Genome 2014, 7, 1–9. [Google Scholar] [CrossRef]
- Shi, A.; Chen, P.; Li, D.; Zheng, C.; Zhang, B.; Hou, A. Pyramiding multiple genes for resistance to soybean mosaic virus in soybean using molecular markers. Mol. Breed. 2009, 23, 113–124. [Google Scholar] [CrossRef]
- Jun, T.H.; Mian, M.R.; Kang, S.T.; Michel, A.P. Genetic mapping of the powdery mildew resistance gene in soybean PI 567301B. Theor. Appl. Genet. 2012, 125, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Palloix, A.; Ayme, V.; Moury, B. Durability of plant major resistance genes to pathogens depends on the genetic background, experimental evidence and consequences for breeding strategies. New Phytol. 2009, 183, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrell, P.L.; Buckler, E.S.; Ross-Ibarra, J. Crop genomics: Advances and applications. Nat. Rev. Genet. 2011, 13, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, J.E. Peer-reviewed surveys indicate positive impact of commercialized GM crops. Nat. Biotechnol. 2010, 28, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Hyten, D.L.; Jia, G.; Quigley, C.V.; Fickus, E.W.; Nelson, R.L.; Cregen, B.P. Development and evaluation of SoySNP50K, a high-density genotyping array for soybean. PLoS ONE 2013, 8, e54985. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, S.; Li, L.; Zhang, X.; Xu, H.; An, D. Mapping QTLs for salt tolerance with additive, epistatic and QTL× treatment interaction effects at seedling stage in wheat. Plant Breed. 2013, 132, 276–283. [Google Scholar] [CrossRef]
- Goff, S.A.; Ricke, D.; Lan, T.H.; Presting, G.; Wang, R.; Dunn, M.; Glazebrook, J.; Sessions, A.; Oeller, P.; Varma, H.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica). Science 2002, 296, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.S.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 2002, 296, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178. [Google Scholar] [CrossRef] [PubMed]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Weckwerth, W. Green systems biology—From single genomes, proteomes and metabolomes to ecosystems research and biotechnology. J. Prot. 2011, 75, 284–305. [Google Scholar] [CrossRef] [PubMed]
- Castillo, A.; Budak, H.; Varshney, R.K.; Dorado, G.; Graner, A.; Hernandez, P. Transferability and polymorphism of barley EST-SSR markers used for phylogenetic analysis in Hordeum chilense. BMC Plant Biol. 2008, 8, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.P.; Tuna, M.; Budak, H.; Huo, N.; Gu, Y.Q.; Steinwand, M.A. Development of SSR markers and analysis of diversity in Turkish populations of Brachypodium distachyon. BMC Plant Biol. 2009, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Hussain, B. Modernization in plant breeding approaches for improving biotic stress resistance in crop plants. Turkish J. Agric. For. 2015, 39, 515–530. [Google Scholar] [CrossRef]
- Saade, S.; Maurer, A.; Shahid, M.; Oakey, H.; Schmöckel, S.M.; Negrão, S.; Pillen, K.; Tester, M. Yield-related salinity tolerance traits identified in a nested association mapping (NAM) population of wild barley. Sci. Rep. 2016, 6, 32586. [Google Scholar] [CrossRef] [PubMed]
- Filiz, E.; Ozdemir, B.S.; Budak, F.; Vogel, J.P.; Tuna, M.; Budak, H. Molecular, morphological and cytological analysis of diverse Brachypodium distachyon inbred lines. Genome 2009, 52, 876–890. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.J.; Šimková, H.; Šafář, J.; Jurman, I.; Cattonaro, F.; Vautrin, S.; Bellec, A.; Berges, H.; Dolezel, J.; Budak, H. Functional features of a single chromosome arm in wheat (1AL) determined from its structure. Funct. Integr. Genom. 2012, 12, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Akpinar, B.A.; Lucas, S.; Budak, H. A large-scale chromosome-specific SNP discovery guideline. Funct. Integr. Genom. 2017, 17, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Winfield, M.O.; Allen, A.M.; Burridge, A.J.; Barker, G.L.; Benbow, H.R.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; Scopes, G.; et al. High-density SNP genotyping array for hexaploid wheat and its secondary and tertiary gene pool. Plant Biotechnol. J. 2016, 14, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.J.; Salantur, A.; Yazar, S.; Budak, H. High-throughput SNP genotyping of modern and wild emmer wheat for yield and root morphology using a combined association and linkage analysis. Funct. Integ. Genom. 2017, 17, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Peleg, Z.; Cakmak, I.; Ozturk, L.; Yazici, A.; Jun, Y.; Budak, H.; Korol, A.B.; Fahima, T.; Saranga, Y. Quantitative trait loci conferring grain mineral nutrient concentrations in durum wheat× wild emmer wheat RIL population. Theor. App. Genet. 2009, 119, 353–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonforte, A.; Sudheesh, S.; Cogan, N.O.; Salisbury, P.A.; Nicolas, M.E.; Materne, M.; Forster, J.W.; Kaur, S. SNP marker discovery, linkage map construction and identification of QTLs for enhanced salinity tolerance in field pea (Pisum sativum L.). BMC Plant. Biol. 2013, 13, 161. [Google Scholar] [CrossRef] [PubMed]
- Lorieux, M. MapDisto: Fast and efficient computation of genetic linkage maps. Mol. Breed. 2012, 30, 1231–1235. [Google Scholar] [CrossRef]
- Masoudi, B.; Mardi, M.; Hervan, E.M.; Bihamta, M.R.; Naghavi, M.R.; Nakhoda, B.; Amini, A. QTL mapping of salt tolerance traits with different effects at the seedling stage of bread wheat. Plant Mol. Biol. Report. 2015, 33, 1790–1803. [Google Scholar] [CrossRef]
- Genc, Y.; Oldach, K.; Verbyla, A.P.; Lott, G.; Hassan, M.; Tester, M.; Wallwork, H.; McDonald, G.K. Sodium exclusion QTL associated with improved seedling growth in bread wheat under salinity stress. Theor. App. Genet. 2010, 121, 877–894. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Balyan, H.S.; Gahlaut, V.; Kulwal, P.L. Phenotyping, genetic dissection and breeding for drought and heat tolerance in common wheat: Status and prospects. Plant Breed. Rev. 2012, 36, 85–168. [Google Scholar]
- Budak, H.; Hussain, B.; Khan, Z.; Ozturk, N.Z.; Ullah, N. From genetics to functional genomics: Improvement in drought signaling and tolerance in wheat. Front. Plant Sci. 2015, 6, 1012. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, S.; Malik, R.; Narwal, S.; Tyagi, B.S.; Mittal, V.; Kharub, A.S.; Tiwari, V.; Sharma, I. Genetic and molecular dissection of drought tolerance in wheat and barley. J. Wheat Res. 2016, 7, 1–13. [Google Scholar]
- Salvi, S.; Tuberosa, R. The crop QTLome comes of age. Curr. Opi. Biotec. 2015, 32, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Singh, K.; Patil, R.V.; Kadam, S.; Bharti, S.; Prasad, P.; Singh, N.K.; Khanna-Chopra, R. Genomic regions associated with grain yield under drought stress in wheat (Triticum aestivum L.). Euphytica 2015, 203, 449–467. [Google Scholar] [CrossRef]
- Kirigwi, F.M.; Van Ginkel, M.; Brown-Guedira, G.; Gill, B.S.; Paulsen, G.M.; Fritz, A.K. Markers associated with a QTL for grain yield in wheat under drought. Mol. Breed. 2007, 20, 401–413. [Google Scholar] [CrossRef]
- Pinto, R.S.; Reynolds, M.P.; Mathews, K.L.; McIntyre, C.L.; Olivares-Villegas, J.J.; Chapman, S.C. Heat and drought adaptive QTL in a wheat population designed to minimize confounding agronomic effects. Theor. Appl. Genet. 2010, 121, 1001–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quarrie, S.A.; Pekic Quarrie, S.; Radosevic, R.; Rancic, D.; Kaminska, A.; Barnes, J.D.; Leverington, M.; Ceoloni, C.; Dodig, D. Dissecting a wheat QTL for yield present in a range of environments: From the QTL to candidate genes. J. Exp. Bot. 2006, 57, 2627–2637. [Google Scholar] [CrossRef] [PubMed]
- Golabadi, M.; Arzani, A.; Mirmohammadi Maibody, S.A.M.; Tabatabaei, B.E.S.; Mohammadi, S.A. Identification of microsatellite markers linked with yield components under drought stress at terminal growth stages in durum wheat. Euphytica 2011, 177, 207–221. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P.; McIntyre, C.L.; Mathews, K.L.; Jalal Kamali, M.R.; Mossad, M.; Feltaous, Y.; Tahir, I.S.A.; Chatrath, R.; Ogbonnaya, F.; et al. QTL for yield and associated traits in the Seri/Babax population grown across several environments in Mexico, in the West Asia, North Africa and South Asia regions. Theor. Appl. Genet. 2013, 126, 971–984. [Google Scholar] [CrossRef] [PubMed]
- Maccaferri, M.; Sanguineti, M.C.; Corneti, S.; Ortega, J.L.A.; Ben Salem, M.; Bort, J.; DeAmbrogio, E.; Del Moral, L.F.G.; Demontis, A.; El-Ahmed, A.; et al. Quantitative trait loci for grain yield and adaptation of durum wheat (Triticum durum Desf.) across a wide range of water availability. Genetics 2008, 178, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Salem, K.F.M.; Roder, M.S.; Borner, A. Identification and mapping quantitative trait loci for stem reserve mobilisation in wheat (Triticum aestivum L.). Cereal Res. Commun. 2007, 35, 1367–1374. [Google Scholar] [CrossRef]
- Bennett, D.; Izanloo, A.; Reynolds, M.; Kuchel, H.; Langridge, P.; Schnurbusch, T. Genetic dissection of grain yield and physical grain quality in bread wheat (Triticum aestivum L.) under water-limited environments. Theor. Appl. Genet. 2012, 125, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sehgal, S.K.; Kumar, U.; Prasad, P.V.V.; Joshi, A.K.; Gill, B.S. Genomic characterization of drought tolerance-related traits in spring wheat. Euphytica 2012, 186, 265–276. [Google Scholar] [CrossRef]
- Myles, S.; Peiffer, J.; Brown, P.J.; Ersoz, E.S.; Zhang, Z.; Costich, D.E.; Buckler, E.S. Association mapping: Critical considerations shift from genotyping to experimental design. Plant Cell 2009, 21, 2194–2202. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J.; Burgueno, J.; Dreisigacker, S.; Vargas, M.; Herrera-Foessel, S.A.; Lillemo, M.; Singh, R.P.; Trethowen, R.; Warburton, M.; Franco, J.; et al. Association analysis of historical bread wheat germplasm using additive genetic covariance of relatives and population structure. Genetics 2007, 177, 1889–1913. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, N.; Latos, P.; Hynes-Allen, A.; Loos, R.; Leaford, D.; O’Shaughnessy, A.; Mosaku, O.; Signolet, J.; Brennecke, P.; Kalkan, T.; et al. NuRD suppresses pluripotency gene expression to promote transcriptional heterogeneity and lineage commitment. Cell Stem Cell 2012, 10, 583–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heffner, E.L.; Sorrells, M.E.; Jannink, J.L. Genomic selection for crop improvement. Crop Sci. 2009, 49, 1–12. [Google Scholar] [CrossRef]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: A review. Plant Meth. 2013, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Yu, D.; Wang, D.; Bai, X.; Zhu, Y.; Guo, C. Genomic selection of seed weight based on low-density SCAR markers in soybean. Genet. Mol. Res. 2012, 12, 2178–2188. [Google Scholar] [CrossRef] [PubMed]
- Jannink, J.L.; Lorenz, A.J.; Iwata, H. Genomic selection in plant breeding: From theory to practice. Brief. Func. Genom. 2010, 9, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Nadaf, J.; Riggio, V.; Yu, T.P.; Pong-Wong, R. Effect of the prior distribution of SNP effects on the estimation of total breeding value. BMC Proc. 2012, 6, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogutu, J.O.; Schulz-Streeck, T.; Piepho, H.P. Genomic selection using regularized linear regression models: Ridge regression, lasso, elastic net and their extensions. BMC Proc. 2012, 6, S10. [Google Scholar] [CrossRef] [PubMed]
- Mochida, K.; Yamazaki, Y.; Ogihara, Y. Discrimination of homoeologous gene expression in hexaploid wheat by SNP analysis of contigs grouped from a large number of expressed sequence tags. Mol. Genet. Genom. 2004, 270, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Leader, D.J. Transcriptional analysis and functional genomics in wheat. J. Cereal Sci. 2005, 41, 149–163. [Google Scholar] [CrossRef]
- Close, T.J.; Wanamaker, S.I.; Caldo, R.A.; Turner, S.M.; Ashlock, D.A.; Dickerson, J.A.; Wing, R.A.; Muehlbauer, G.J.; Kleinhofs, A.; Wise, R.P. A new resource for cereal genomics: 22K barley GeneChip comes of age. Plant Physio. 2004, 134, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Ueda, A.; Kathiresan, A.; Bennett, J.; Takabe, T. Comparative transcriptome analyses of barley and rice under salt stress. Theor. App. Genet. 2006, 112, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Walia, H.; Wilson, C.; Condamine, P.; Ismail, A.M.; Xu, J.; Cui, X.; Close, T.J. Array-based genotyping and expression analysis of barley cv. Maythorpe and Golden Promise. BMC Genom. 2007, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Hong, M.J.; Jang, J.H.; Seo, Y.W. cDNA-AFLP analysis reveals differential gene expression in response to salt stress in Brachypodium distachyon. Genes Genom. 2012, 34, 475–484. [Google Scholar] [CrossRef]
- Garg, B.; Puranik, S.; Misra, S.; Tripathi, B.N.; Prasad, M. Transcript profiling identifies novel transcripts with unknown functions as primary response components to osmotic stress in wheat (Triticum aestivum L.). Plant Cell Tissue Organ Cult. 2013, 113, 91–101. [Google Scholar] [CrossRef]
- Meng, C.; Quan, T.Y.; Li, Z.Y.; Cui, K.L.; Yan, L.; Liang, Y.; Dai, J.L.; Xia, J.M.; Liu, S.W. Transcriptome profiling reveals the genetic basis of alkalinity tolerance in wheat. BMC Genom. 2017, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Poersch-Bortolon, L.B.; Pereira, J.F.; Nhani Junior, A.; Gonzáles, H.H.S.; Torres, G.A.M.; Consoli, L.; Arenhart, R.A.; Bodanese-zanettini, M.H.; Margis-Pinheiro, M. Gene expression analysis reveals important pathways for drought response in leaves and roots of a wheat cultivar adapted to rainfed cropping in the Cerrado biome. Genet. Mol. Biol. 2016, 39, 629–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Li, R.; Wang, H.; Li, D.; Wang, X.; Zhang, Y.; Zhen, W.; Duan, H.; Yan, G.; Li, Y. Transcriptomics analyses reveal wheat responses to drought stress during reproductive stages under field conditions. Front. Plant Sci. 2017, 8, 592. [Google Scholar] [CrossRef] [PubMed]
- Goyal, E.; Amit, S.K.; Singh, R.S.; Mahato, A.K.; Chand, S.; Kanika, K. Transcriptome profiling of the salt-stress response in Triticum aestivum cv. Kharchia Local. Sci. Rep. 2016, 6, 27752. [Google Scholar] [CrossRef] [PubMed]
- Haque, E.; Kawaguchi, K.; Komatsu, S. Analysis of proteins in aerenchymatous seminal roots of wheat grown in hypoxic soils under waterlogged conditions (supplementary material). Protein Pept. Lett. 2011, 18, 912–924. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.J.; Oyanagi, A.; Komatsu, S. Cell wall proteome of wheat roots under flooding stress using gel-based and LC MS/MS-based proteomics approaches. Biochim. Biophys. Acta 2010, 1804, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, S.; Roy Choudhury, S.; Pandey, S. Comparative quantitative proteomics analysis of the ABA response of roots of drought-sensitive and drought-tolerant wheat varieties identifies proteomic signatures of drought adaptability. J. Prot. Res. 2014, 13, 1688–1701. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Cavaliere, C.; Foglia, P.; Gubbiotti, R.; Samperi, R.; Laganà, A. Analysis of drought responsive proteins in wheat (Triticum durum) by 2D-PAGE and MALDI-TOF mass spectrometry. Plant Sci. 2009, 177, 570–576. [Google Scholar] [CrossRef]
- Demirevska, K.; Zasheva, D.; Dimitrov, R.; Simova-Stoilova, L.; Stamenova, M.; Feller, U. Drought stress effects on Rubisco in wheat: Changes in the Rubisco large subunit. Acta Physiol. Plant. 2009, 31, 1129. [Google Scholar] [CrossRef]
- Kamal, A.H.M.; Cho, K.; Choi, J.S.; Jin, Y.; Park, C.S.; Lee, J.S.; Woo, S.H. Patterns of protein expression in water-stressed wheat chloroplasts. Biol. Plant. 2013, 57, 305–312. [Google Scholar] [CrossRef]
- Jiang, S.S.; Liang, X.N.; Li, X.; Wang, S.L.; Lv, D.W.; Ma, C.Y.; Li, X.H.; Ma, W.J.; Yan, Y.M. Wheat drought-responsive grain proteome analysis by linear and nonlinear 2-DE and MALDI-TOF mass spectrometry. Int. J. Mol. Sci. 2012, 13, 16065–16083. [Google Scholar] [CrossRef] [PubMed]
- Ford, K.L.; Cassin, A.; Bacic, A.F. Quantitative proteomic analysis of wheat cultivars with differing drought stress tolerance. Front. Plant Sci. 2011, 2, 44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Lv, D.; Ge, P.; Bian, Y.; Chen, G.; Zhu, G.; Li, X.; Yan, Y. Phosphoproteome analysis reveals new drought response and defense mechanisms of seedling leaves in bread wheat (Triticum aestivum L.). J. Prot. 2014, 109, 290–308. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Jørgensen, A.D.; Li, H.; Søndergaard, I.; Finnie, C.; Svensson, B.; Jiang, D.; Wollenweber, B.; Jacobsen, S. Implications of high-temperature events and water deficits on protein profiles in wheat (Triticum aestivum L. cv. Vinjett) grain. Proteomics 2011, 11, 1684–1695. [Google Scholar] [CrossRef] [PubMed]
- Hurkman, W.J.; Vensel, W.H.; Tanaka, C.K.; Whitehand, L.; Altenbach, S.B. Effect of high temperature on albumin and globulin accumulation in the endosperm proteome of the developing wheat grain. J. Cereal Sci. 2009, 49, 12–23. [Google Scholar] [CrossRef]
- Kamal, A.H.M.; Cho, K.; Kim, D.E.; Uozumi, N.; Chung, K.Y.; Lee, S.Y.; Choi, J.S.; Cho, S.W.; Shen, C.S.; Woo, S.H. Changes in physiology and protein abundance in salt-stressed wheat chloroplasts. Mol. Biol. Rep. 2012, 39, 9059–9074. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Yan, X.; Li, X.; Guo, G.; Hu, Y.; Ma, W.; Yan, Y. Proteome analysis of wheat leaf under salt stress by two-dimensional difference gel electrophoresis (2D-DIGE). Phytochemistry 2011, 72, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, R.P.; Millar, A.H.; Taylor, N.L. Wheat mitochondrial proteomes provide new links between antioxidant defense and plant salinity tolerance. J. Prot. Res. 2010, 9, 6595–6604. [Google Scholar] [CrossRef] [PubMed]
- Delisle, G.; Champoux, M.; Houde, M. Characterization of oxalate oxidase and cell death in Al-sensitive and tolerant wheat roots. Plant Cell Physiol. 2001, 42, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.W.; Roy, S.K.; Kamal, A.H.M.; Cho, K.; Cho, S.W.; Park, C.S.; Choi, J.S.; Komatsu, S.; Woo, S.H. Proteome analysis of roots of wheat seedlings under aluminum stress. Mol. Biol. Rep. 2014, 41, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Peng, X.; Xuan, H.; Wei, L.; Yang, Y.; Guo, T.; Kang, G. Proteomic analysis of leaves and roots of common wheat (Triticum aestivum L.) under copper-stress conditions. J. Proteome Res. 2013, 12, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.H.M.; Cho, K.; Komatsu, S.; Uozumi, N.; Choi, J.S.; Woo, S.H. Towards an understanding of wheat chloroplasts: A methodical investigation of thylakoid proteome. Mol. Biol. Rep. 2012, 39, 5069–5083. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Kamal, A.H.M.; Shin, K.H.; Choi, J.S.; Heo, H.Y.; Woo, S.H. Large-scale proteome investigation in wild relatives (A., B and D genomes) of wheat. Acta Biochim. Biophys. Sin. 2010, 42, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, H.; Xu, Y.; Li, X.X.; Zhang, H.J. Differential proteomic analysis of cadmium-responsive proteins in wheat leaves. Biol. Plant. 2011, 55, 586–590. [Google Scholar] [CrossRef]
- Cailin, G.E.; Yan, D.I.N.G.; Zegang, W.A.N.G.; Dingzhen, W.A.N.; Yulong, W.A.N.G.; Shang, Q.; Shishi, L.U.O. Responses of wheat seedlings to cadmium, mercury and trichlorobenzene stresses. J. Environ. Sci. 2009, 21, 806–813. [Google Scholar]
- Bino, R.J.; Hall, R.D.; Fiehn, O.; Kopka, J.; Saito, K.; Draper, J.; Nikolau, B.J.; Mendes, P.; Roessner-Tunali, U.; Beale, M.H.; et al. Potential of metabolomics as a functional genomics tool. Trends Plant Sci. 2004, 9, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Matsuda, F. Metabolomics for functional genomics, systems biology and biotechnology. Ann. Review Plant Biol. 2010, 61, 463–489. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Hirai, M.Y.; Yonekura-Sakakibara, K. Decoding genes with coexpression networks and metabolomics–‘majority report by precogs’. Trends Plant Sci. 2008, 13, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.S.; Galbraith, D.W.; Dai, S.Y.; Griffin, P.; Stewart Jr, C.N. Plant systems biology comes of age. Trends Plant Sci. 2008, 13, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R. Metabolome characterization in plant system analysis. Funct. Plant Biol. 2003, 30, 111–120. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Saito, K. Review: Genetically modified plants for the promotion of human health. Biotech. Lett. 2006, 28, 1983–1991. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Yang, L.; Zhang, D.; Shi, J. Plant metabolomics: An indispensable system biology tool for plant science. Inter. J. Mol. Sci. 2016, 17, 767. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Li, K.; Alseekh, S.; Omranian, N.; Zhao, L.; Zhou, Y.; Xiao, Y.; Jin, M.; Ying, N.; Liu, H.; et al. Genetic determinants of the network of primary metabolism and their relationships to plant performance in a maize recombinant inbred line population. Plant Cell 2015, 27, 1839–1856. [Google Scholar] [CrossRef] [PubMed]
- Kusano, M.; Saito, K. Role of metabolomics in crop improvement. J. Plant Biochem. Biotech. 2012, 21, 24–31. [Google Scholar] [CrossRef]
- Hall, R.; Beale, M.; Fiehn, O.; Hardy, N.; Sumner, L.; Bino, R. Plant metabolomics: The missing link in functional genomics strategies. Plant Cell 2002, 14, 1437–1440. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Schauer, N. Metabolomics-assisted breeding: A viable option for crop improvement. Trends Genet. 2009, 25, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, F.; Hirai, M.Y.; Sasaki, E.; Akiyama, K.; Yonekura-Sakakibara, K.; Provart, N.J.; Sakurai, T.; Shimada, Y.; Saito, K. AtMetExpress development: A phytochemical atlas of Arabidopsis development. Plant Physiol. 2010, 152, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Huhman, D.V.; Sumner, L.W. Mass spectrometry strategies in metabolomics. J. Biol. Chem. 2011, 286, 25435–25442. [Google Scholar] [CrossRef] [PubMed]
- Tohge, T.; Fernie, A.R. Web-based resources for mass-spectrometry-based metabolomics: A user’s guide. Phytochemistry 2009, 70, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Afendi, F.M.; Okada, T.; Yamazaki, M.; Hirai-Morita, A.; Nakamura, Y.; Nakamura, K.; Ikeda, S.; Takahashi, H.; Altaf-ul Amin, M.; Darusman, M.K.; et al. KNApSAcK family databases: Integrated metabolite–plant species databases for multifaceted plant research. Plant Cell Physiol. 2011, 53, e1. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.K.; Roorkiwal, M.; Singh, V.K.; Ramalingam, A.; Kudapa, H.; Thudi, M.; Chitikineni, A.; Rathore, A.; Varshney, R.K. Emerging genomic tools for legume breeding: Current status and future prospects. Front. Plant Sci. 2016, 7, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urano, K.; Kurihara, Y.; Seki, M.; Shinozaki, K. ‘Omics’ analyses of regulatory networks in plant abiotic stress responses. Curr. Opin. Plant Biol. 2010, 13, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Obata, T.; Fernie, A.R. The use of metabolomics to dissect plant responses to abiotic stresses. Cell. Mol. Life Sci. 2012, 69, 3225–3243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zivy, M.; Wienkoop, S.; Renaut, J.; Pinheiro, C.; Goulas, E.; Carpentier, S. The quest for tolerant varieties: The importance of integrating “omics” techniques to phenotyping. Front. Plant Sci. 2015, 6, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, T.R.; Gill, B.S. The deletion stocks of common wheat. J. Heredity 1996, 87, 295–307. [Google Scholar] [CrossRef]
- Erayman, M.; Sandhu, D.; Sidhu, D.; Dilbirligi, M.; Baenziger, P.S.; Gill, K.S. Demarcating the gene-rich regions of the wheat genome. Nucleic Acids Res. 2004, 32, 3546–3565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francki, M.G.; Hayton, S.; Gummer, J.; Rawlinson, C.; Trengove, R.D. Metabolomic profiling and genomic analysis of wheat aneuploid lines to identify genes controlling biochemical pathways in mature grain. Plant Biol. J. 2016, 14, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Michaletti, A.; Naghavi, M.R.; Toorchi, M.; Zolla, L.; Rinalducci, S. Metabolomics and proteomics reveal drought-stress responses of leaf tissues from spring-wheat. Sci. Rep. 2018, 8, 5710. [Google Scholar] [CrossRef] [PubMed]
- Salt, D.E.; Baxter, I.; Lahner, B. Ionomics and the study of the plant ionome. Annu. Rev. Plant Biol. 2008, 59, 709–733. [Google Scholar] [CrossRef] [PubMed]
- Shelden, M.C.; Roessner, U. Advances in functional genomics for investigating salinity stress tolerance mechanisms in cereals. Front. Plant Sci. 2013, 4, 123. [Google Scholar] [CrossRef] [PubMed]
- Satismruti, K.; Senthil, N.; Vellaikumar, S.; Ranjani, R.V.; Raveendran, M. Plant ionomics: A platform for identifying novel gene regulating plant mineral nutrition. Am. J. Plant Sci. 2013, 4, 1309–1315. [Google Scholar] [CrossRef]
- Tuberosa, R. Phenotyping for drought tolerance of crops in the genomics era. Front. Physiol. 2012, 3, 347. [Google Scholar] [CrossRef] [PubMed]
- Margulies, M.; Egholm, M.; Altman, W.E.; Attiya, S.; Bader, J.S.; Bemben, L.A.; Berca, J.; Braverman, M.S.; Chen, Y.J.; Chen, Z.; et al. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005, 437, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, K.F.; Rogers, J.; Pozniak, C.; Eversole, K.; Feuillet, C.; Gill, B.; Friebe, B.; Lukaszewski, A.J.; Sourdille, P.; Endo, T.R.; et al. International Wheat Genome Sequencing Consortium. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar]
- Shah, T.; Tayyaba, A.; Sadia, L.; Mehmood, A.N. Genome editing tools: Advancing crop transformation and overview of tools. Plant Physiol. Biochem. 2018. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| S.No. | QTL | Linked Markers | Position | Env. a | PVE (R2) b | References |
|---|---|---|---|---|---|---|
| Agronomic traits | ||||||
| A. Grain Yield | ||||||
| 1 | qGYWD.3B.2 | Xgpw7774 | 97.6 | 7/4 | 19.6 | [41] |
| 2 | 4A | Xwmc420 | 90.4 | Mean/2 | 20 | [42] |
| 3 | 4A-a | Xgwm397 | 6 | 7/5 | 23.9 | [43] |
| 4 | Qyld.csdh.7AL | Xgwm322 | 155.9 | 21/11 | 20.0 * | [44] |
| B. 1000-Grain weight | ||||||
| 1 | 3B | Xbarc101 | 86.1 | Mean/2 | 45.2 | [45] |
| 2 | QTgw-7D-b | XC29-P13 | 12.5 | 11/10 | 21.9 | [46] |
| C. Days to Heading | ||||||
| 1 | QDh-7D.b | XC29-P13 | 12.5 | 11/11 | 22.7 | [47] |
| 2 | QHd.idw-2A.2 | Xwmc177 | 46.1 | 13/16 | 32.2 | [46] |
| D. Days to Maturity | ||||||
| 1 | QDm-7D.b | X7D-acc/cat-10 | 2.7 | 11/10 | 22.7 | [48] |
| Physiological Traits | ||||||
| A. Stem Reserve Mobilization | ||||||
| 1 | QSrm.ipk-2D | Xgwm249a | 142 | 2/2 | 42.2 | [48] |
| 2 | QSrm.ipk-5D | Xfbb238b | 19 | 2/2 | 37.5 | [48] |
| 3 | QSrm.ipk-7D | Xfbb189b | 338 | 2/2 | 21 | [48] |
| B. Water Soluble Carbohydrate | ||||||
| 1 | QWsc-c.aww-3A | Xwmc0388A | 64.9 | 2/2 | 19 | [49] |
| C. SPAD/Chlorophyll Content | ||||||
| 1 | Qchl.ksu-3B | Xbarc68 | 67.2 | 3/2 | 59.1 | [50] |
| Stress/Conditions | Treatment Time and Dose | Cultivar | Organ/Organelle | Proteomic Technologies | Stress Induced Modulation of Metabolic Pathways | Differentially Expressed Protein Classification | References | |
|---|---|---|---|---|---|---|---|---|
| Functions | Localizations | |||||||
| Flooding | 7 d | Bobwhite line SH 9826 | Seminal root | 2-DE, nano LC-MS/MS | Antioxidant defense | StrRes | - | [71] |
| Flooding | 2 d | Shiroganekomugi | Root | 2-DE, nano LC-MS/MS | Carbohydrate (Glycolysis) | EnMet, ProtMet, SigTran, Tranp | Cell wall | [72] |
| Drought | 100 d | Opata, Nesser | Root | iTRAQ | Energy metabolism, Replication, Repair | EnStr, Oxired, Trans | Mem, Cyto, Cell wall, Mito, Nucl, Plast, Vacu | [73] |
| Drought | 7 d | Ofanto | Leaf | 2-DE, MALDI-TOF | Carbohydrate (Glycolysis, gluconeogenesis) | PTR, StrRes, TCA, ROSsca, AAB, GG | - | [74] |
| Drought | 7 d | Katya, Sadovo, Zlatitza, Miziya | Leaf | SDS-PAGE, 2-DE | Energy (photosynthesis) | EnMet, EnvDevS | Chlo | [75] |
| Drought | 9 d | Keumkang | Leaf | 2-DE, MALDI-TOF/TOF | Energy (photosynthesis) | Photo | Chlo | [76] |
| Drought | 10, 15, 20 and 25 d | Janz, Kauz | Seed | 2-DE, MALDI-TOF | Carbohydrate metabolism | ROSsca, CarMet, SigTran | - | [77] |
| Drought | 14, 24 d | Kukri, Excalibur | Leaf | iTRAQ | Energy (photosynthesis) | Photo, GG, ProtF, Tranp, EnStr | - | [78] |
| Drought | 20% PEG | Hanxuan 10 and Ningchun 47 | Leaf | nano LC-MS/MS | Antioxidant defense | DRM, SigTran, StrRes, ROSsca | - | [79] |
| Heat and Drought | 10 d | Vinjett | Kernel | 2-DE, MALDI-TOF | Carbohydrate (Glycolysis) | CarboMet, STP | - | [80] |
| High Temperature | 37 °C d, 28 °C N/10 d, 20 d | Butte 86 | Endosperm | 2-DE, QSTAR PULSAR-TOF | Carbohydrate metabolism | CarboMet, NitMet, ProtMet, StrRes, STP, SigTran, Tranp, Trans | - | [81] |
| Salt | 150 mM NaCl/1 d, 2 d, 3 d | Keumkang | Leaf | 2-DE, LTQ-FTICR-MS | Energy (photosynthesis) | Photo, StrRes | Chlo | [82] |
| Salt | 1.0, 1.5, 2.0 and 2.5% NaCl in HS/2 d | Zhenhmai 9023 | Leaf | 2D-DIGE/Q-TOF-MS | Carbohydrate metabolism | CarMet, ProtF, Tranp, ROS, ATP | - | [83] |
| Salt | 200 mM | Wyalkatchem, Janz | Shoot | 2-DE, LC-MS/MS | - | - | Mito | [84] |
| Aluminum | 250 µM/2 d, 3 d | Atlas-66, Fredrick | Root | SDS-PGE, Immunoblot | Signaling pathway | Oxi | - | [85] |
| Aluminum | 100, 150 µM/5 d | Keumkang | Root | 2-DE, LTQ-FTICR-MS | Energy (Glycolysis) | Gly, Tranp, SigTran, StrRes, EnMet | - | [86] |
| Copper | 100 µM/3 d | Yumai 34 | Root, Leaf | 2-DE, HPLC-Chip/ESI-Q-TOF/MS/MS | Energy (photosynthesis), antioxidant defense | StrRes, SigTran, ProtMet, CarMet, Photo, EnMet | - | [87] |
| Protein Profiling | 20 d | Keumkang | Leaf | SDS-PAGE, LTQ-FTICR | Energy (photosynthesis) | COB, DevPro, DRM, ProtF, ProtMet, StrRes, Tranp, Trans | Chlo | [88] |
| Protein Profiling | Mature seed | Wild type (AA, BB, DD genome) | Seed | SDS-PAGE, nano LC-MS/MS | Carbohydrate metabolism | StrRes, EnMet, ProtS, CGD, COD, ProtF, SigTran, STP, Tranp | - | [89] |
| Cadmium | 10, 100 and 200 µM | Yangmai 15 | Leaf | IPG, MALDI-TOF | Energy (photosynthesis) | Oxi, ProtMet, Photo | - | [90] |
| Cadmium | 0.5 mM/L | Yangmai 13 | Leaf | IPG, MALDI-TOF | Antioxidant defense | ROSsca | - | [91] |
| Resources | Description/URL |
|---|---|
| Genome sequence | Coordinated effort underway by the IWGSC (http://www.wheatgenome.org) Recognized as a priority by the research community (http://www.csrees.usda.gov/nea/plants/pdfs/wheat_conference_summary.pdf) |
| ESTs 1 | 1,050,791 entries |
| Oligonucleotide microarray | 1,050,791 entries |
| cDNA microarray | Multiple including ~9 K array |
| Tiling microarray | Not currently available |
| Serial Analysis of Gene Expression (SAGE) | Applied for studying, developing wheat caryopsis |
| Massively Parallel Signature Sequencing (MPSS) | Not reported |
| Sequencing-by-synthesis | Roche 454 cDNA sequencing [119] |
| Deletion and aneuploid genetic stocks | Roche 454 cDNA sequencing [119] |
| Transformation | Biolistic- and Agrobacterium-mediated DNA delivery systems |
| Gene knockdown | RNA interference Viral-induced gene silencing |
| Databases/tools | Graingenes (http://wheat.pw.usda.gov) Gramene (http://www.gramene.org) TIGR Genome Database (http://www.tigr.org/tdb/e2k1/tae1) Wheat Genome Project (http://wheat.pw.usda.gov/NSF/htmlversion.html) Wheat Genome Project (http://wheat.pw.usda.gov/NSF/htmlversion.html) wEST (http://wheat.pw.usda.gov/wEST) CerealsDB (http://www.cerealsdb.uk.net) HarvEST Wheat (http://harvest.ucr.edu) PLEXdb (http://plexdb.org) Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo) ArrayExpress (http://www.ebi.ac.uk/microarray-as/ae) GrainSAGE (http://www.scu.edu.au/research/cpcg/igfp/index.php) Wheat SNP Project (http://wheat.pw.usda.gov/SNP/new/index.shtml) [120] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, T.; Xu, J.; Zou, X.; Cheng, Y.; Nasir, M.; Zhang, X. Omics Approaches for Engineering Wheat Production under Abiotic Stresses. Int. J. Mol. Sci. 2018, 19, 2390. https://doi.org/10.3390/ijms19082390
Shah T, Xu J, Zou X, Cheng Y, Nasir M, Zhang X. Omics Approaches for Engineering Wheat Production under Abiotic Stresses. International Journal of Molecular Sciences. 2018; 19(8):2390. https://doi.org/10.3390/ijms19082390
Chicago/Turabian StyleShah, Tariq, Jinsong Xu, Xiling Zou, Yong Cheng, Mubasher Nasir, and Xuekun Zhang. 2018. "Omics Approaches for Engineering Wheat Production under Abiotic Stresses" International Journal of Molecular Sciences 19, no. 8: 2390. https://doi.org/10.3390/ijms19082390
APA StyleShah, T., Xu, J., Zou, X., Cheng, Y., Nasir, M., & Zhang, X. (2018). Omics Approaches for Engineering Wheat Production under Abiotic Stresses. International Journal of Molecular Sciences, 19(8), 2390. https://doi.org/10.3390/ijms19082390