NOX Inhibition Improves β-Adrenergic Stimulated Contractility and Intracellular Calcium Handling in the Aged Rat Heart

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. General Characteristic of Adult and Senescent Rats
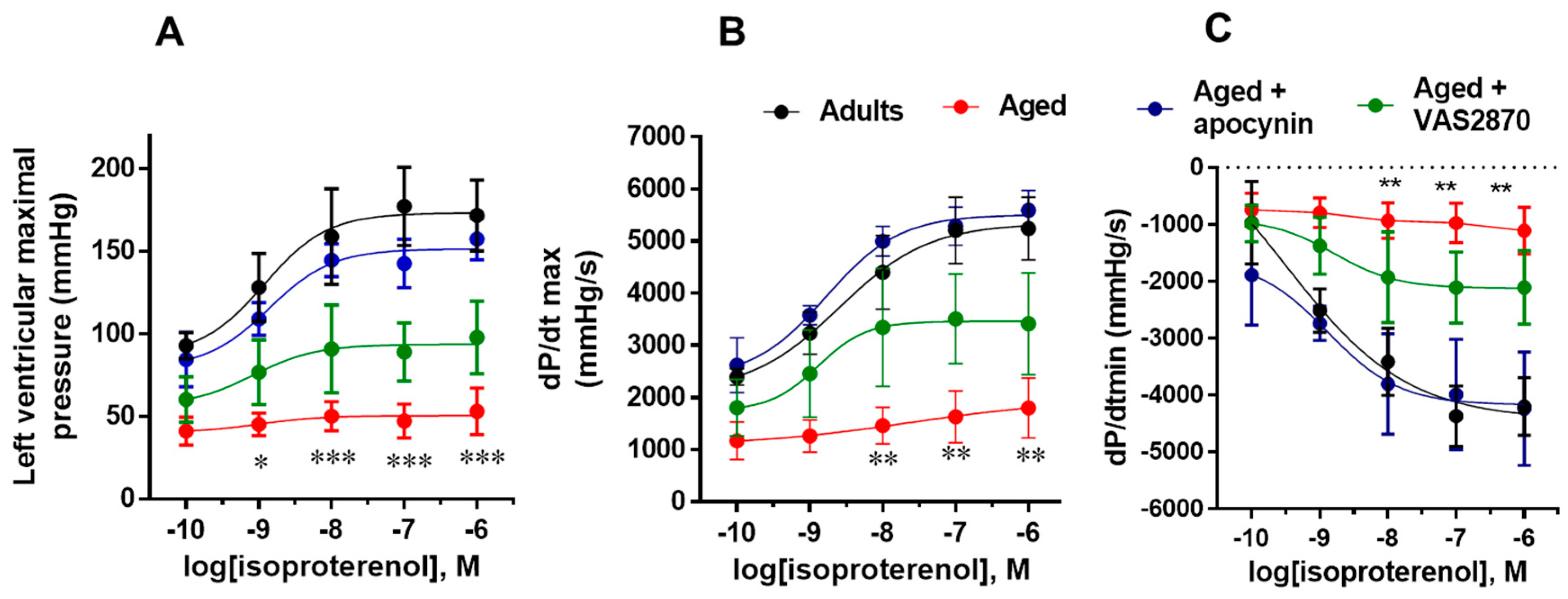
2.2. Isoproterenol Response in the Isolated Heart
2.3. Isolated Cardiomyocytes
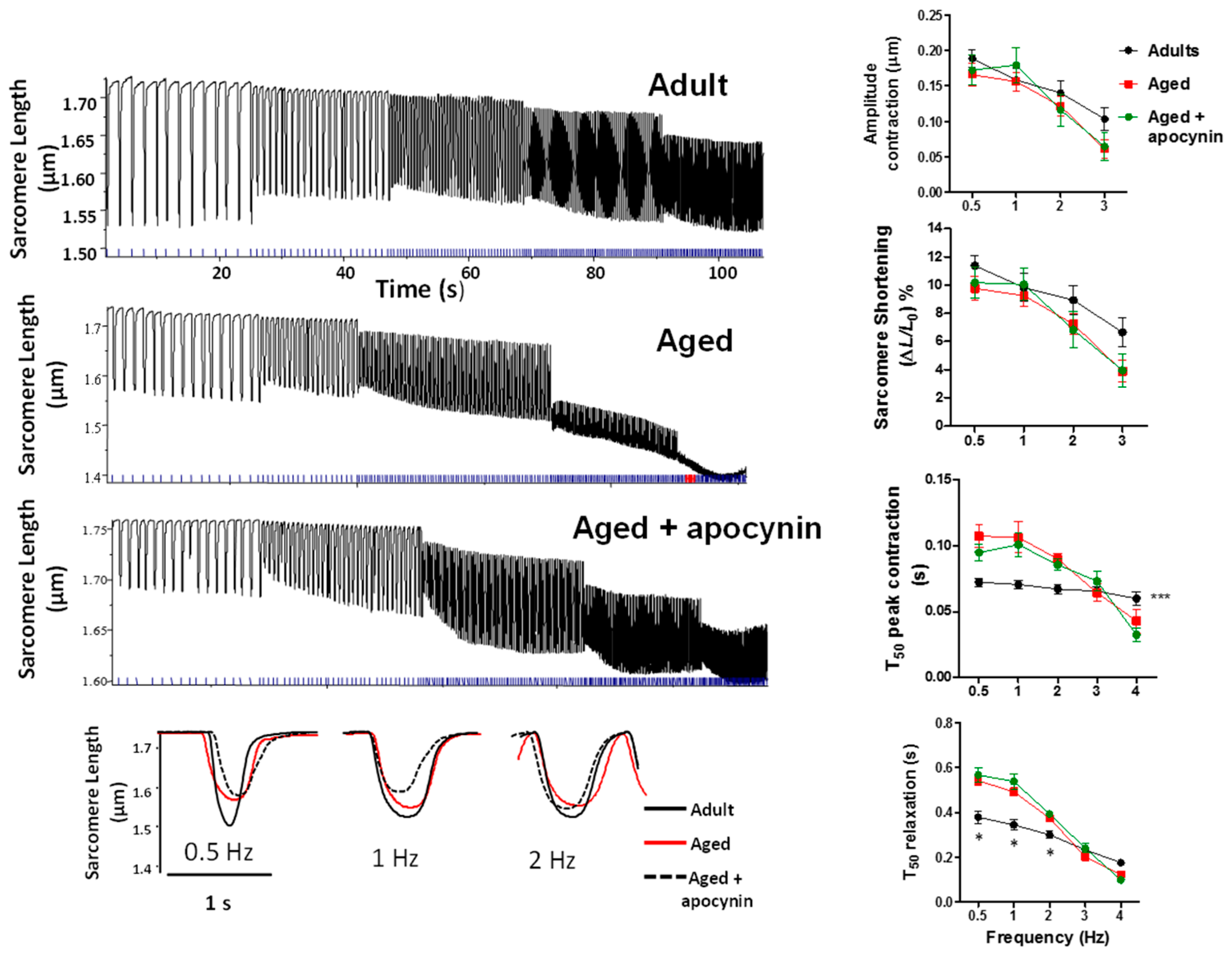
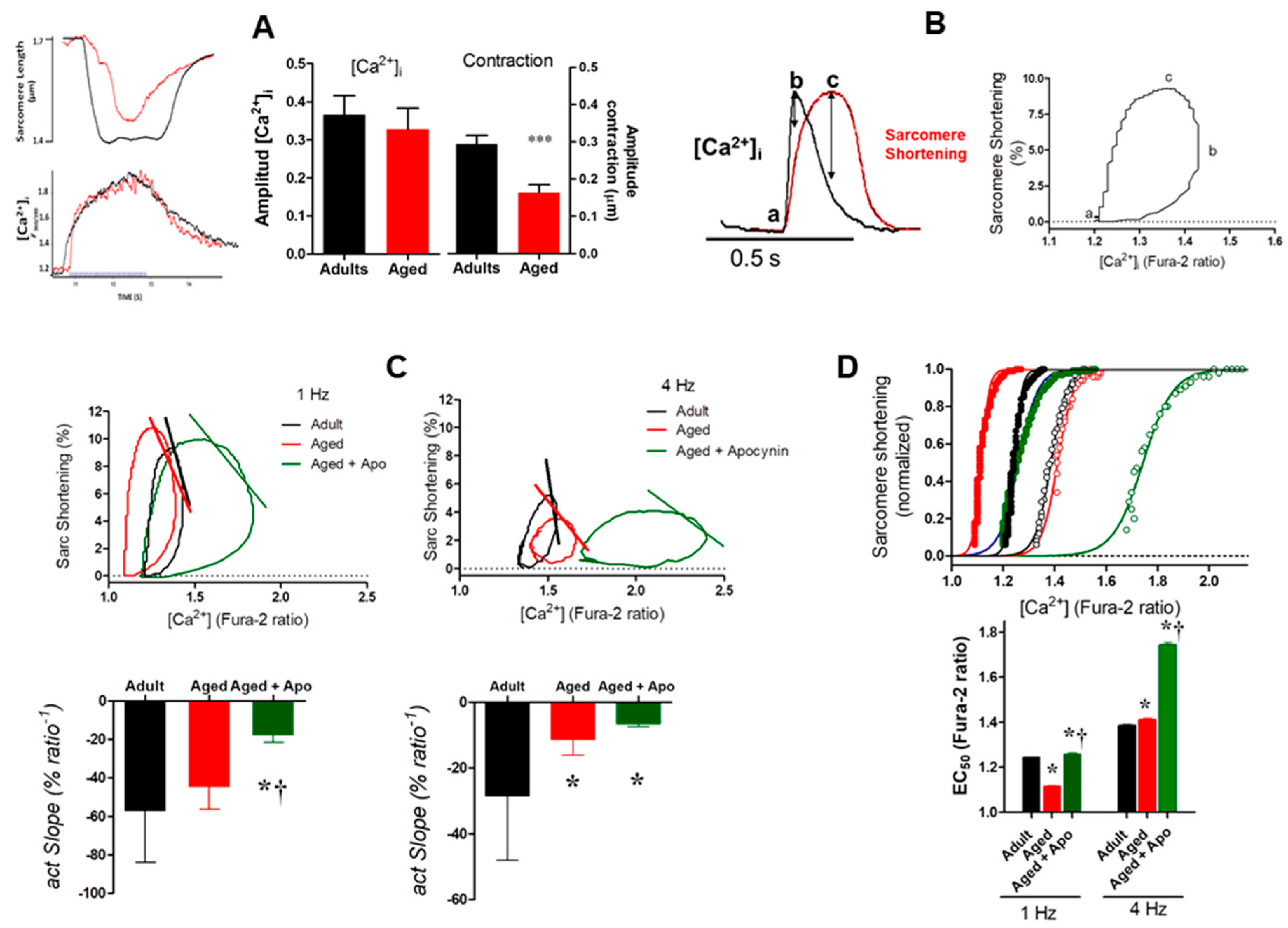
2.3.1. Sarcomere Shortening
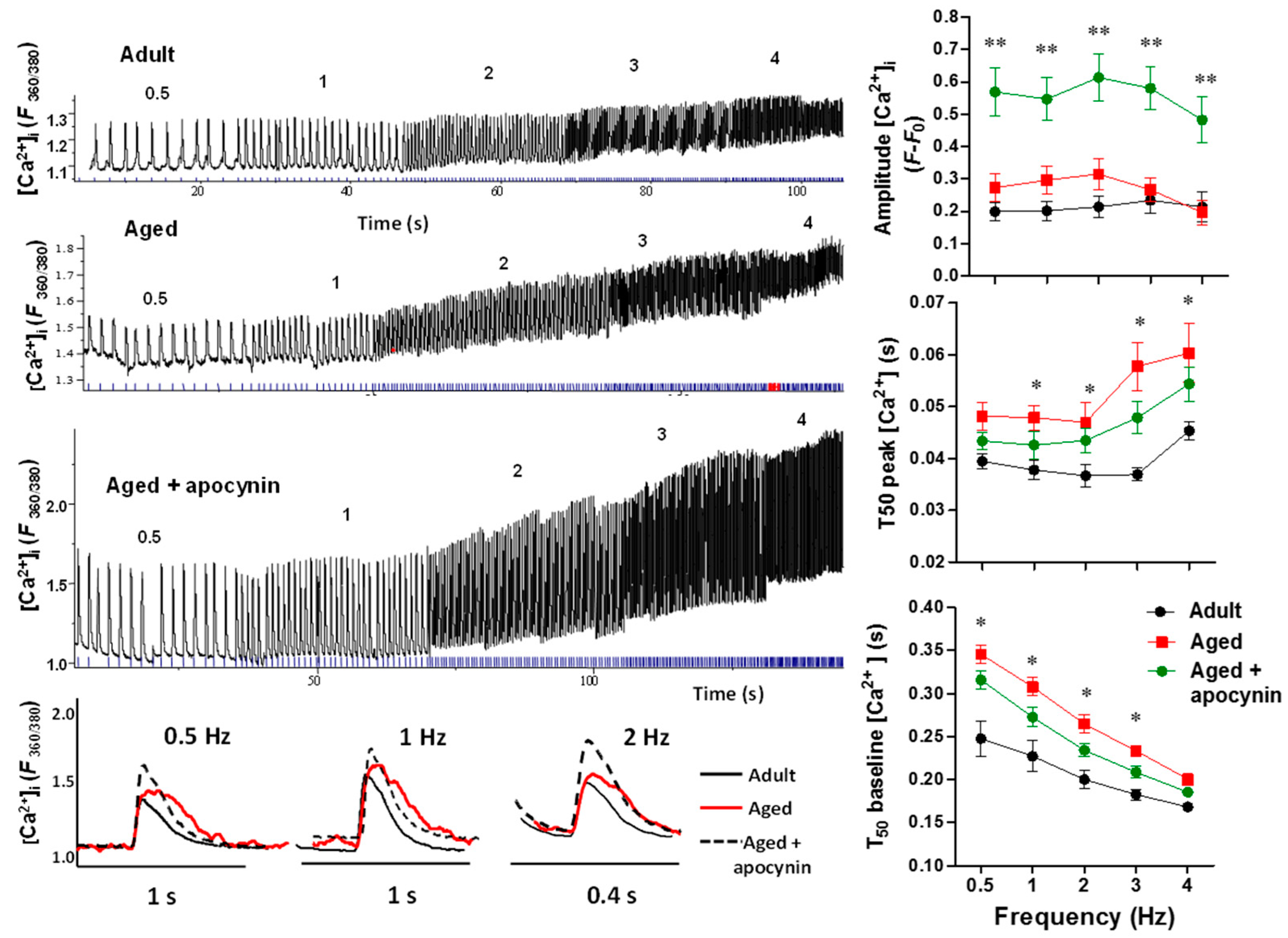
2.3.2. Intracellular Ca2+ Transients
2.3.3. Myofilaments Ca2+ Sensitivity
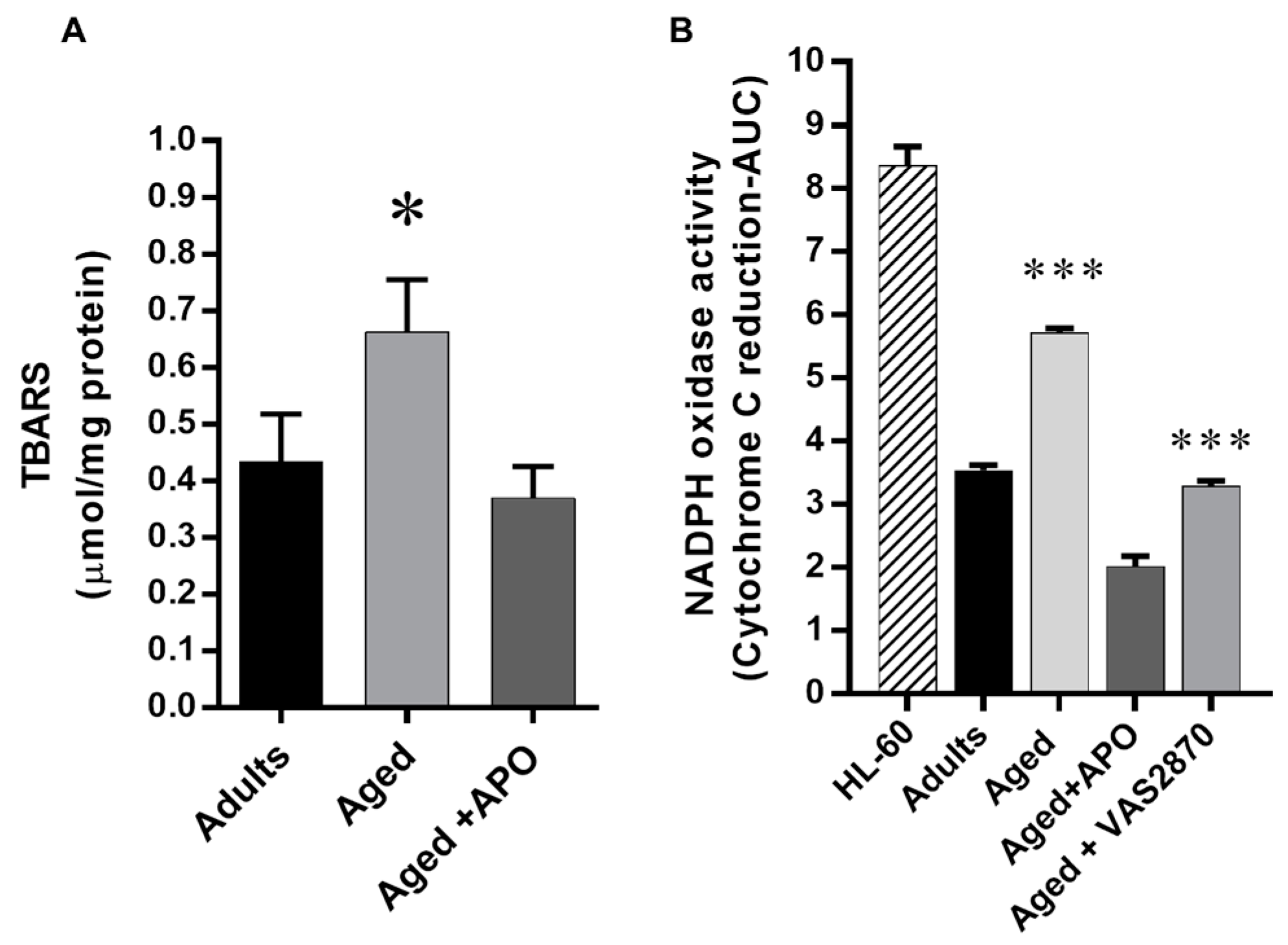
2.4. Cardiac Oxidative Stress
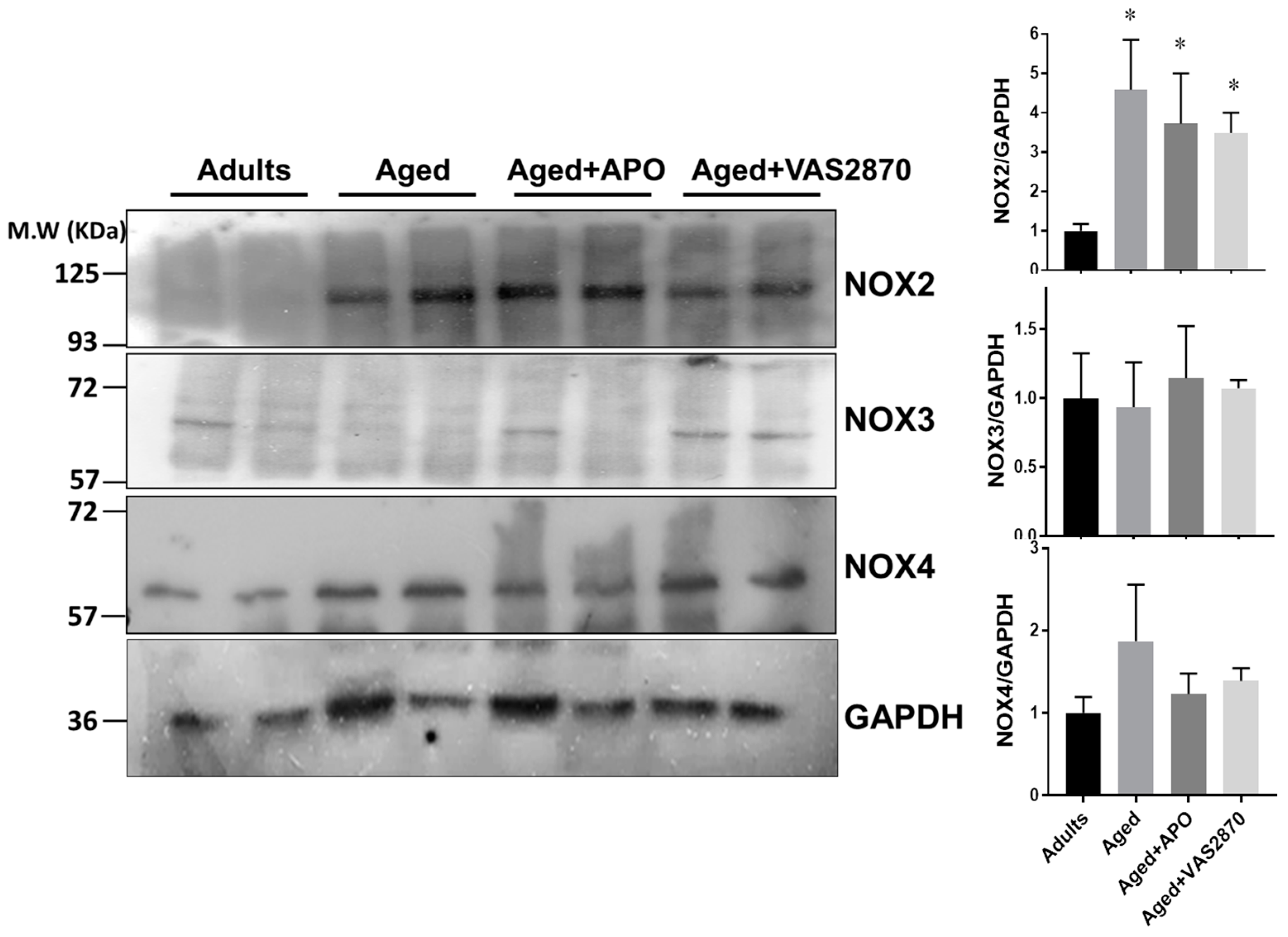
Expression of NOX Isoforms
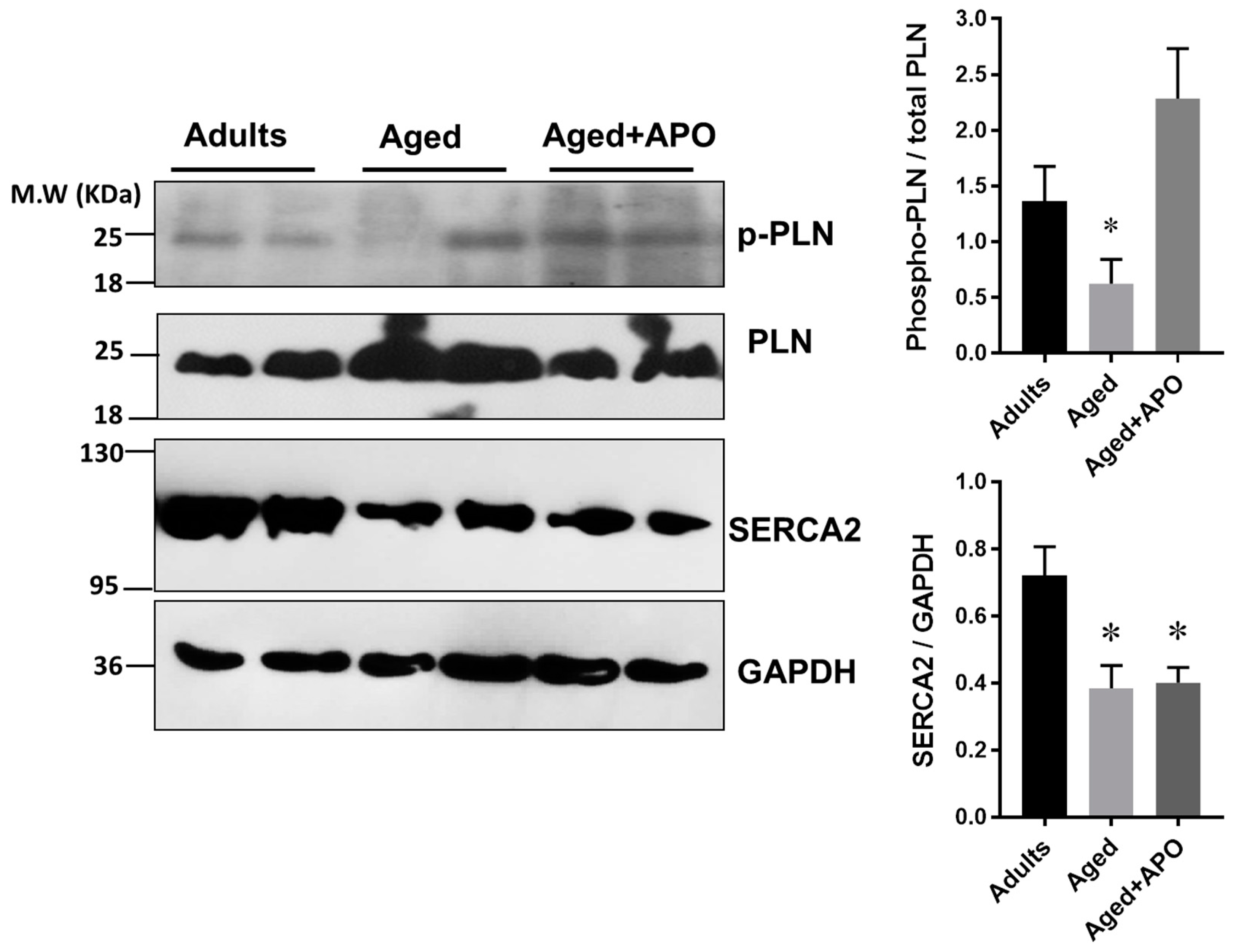
2.5. Ca2+-Handling Proteins Levels
3. Discussion
Study Limitations
4. Materials and Methods
4.1. Animals
4.2. Isolated Heart Experimental Protocols
4.3. Isolation of Rat Ventricular Myocytes
4.4. Measurement of Cardiomyocytes Shortening/Relengthening
4.5. Intracellular Calcium
4.5.1. Analysis of Ca2+ Transient Data
4.5.2. Assessment of Calcium Myofilament Responsiveness
4.6. Oxidative Stress Analysis
NADPH Oxidase Activity
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Boyle, A.J.; Shih, H.; Hwang, J.; Ye, J.; Lee, B.; Zhang, Y.; Kwon, D.; Jun, K.; Zheng, D.; Sievers, R.; et al. Cardiomyopathy of aging in the mammalian heart is characterized by myocardial hypertrophy, fibrosis and a predisposition towards cardiomyocyte apoptosis and autophagy. Exp. Gerontol. 2011, 46, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakatta, E.G.; Levy, D. Arterial and cardiac aging: Major shareholders in cardiovascular disease enterprises: Part II: The aging heart in health: Links to heart disease. Circulation 2003, 107, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Raju, S.V.; Barouch, L.A.; Hare, J.M. Nitric oxide and oxidative stress in cardiovascular aging. Sci. Aging Knowl. Environ. 2005, 21, Re4. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.F.; Chen, T.; Johnson, S.C.; Szeto, H.; Rabinovitch, P.S. Cardiac aging: From molecular mechanisms to significance in human health and disease. Antioxid. Redox Signal. 2012, 16, 1492–1526. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kwak, H.B.; Hord, J.; Kim, J.H.; Lawler, J.M. Exercise training attenuates age-dependent elevation of angiotensin II type 1 receptor and NOX2 signaling in the rat heart. Exp. Gerontol. 2015, 70, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Oudot, A.; Martin, C.; Busseuil, D.; Vergely, C.; Demaison, L.; Rochette, L. NADPH oxidases are in part responsible for increased cardiovascular superoxide production during aging. Free Radic. Biol. Med. 2006, 40, 2214–2222. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.H. Aging: A revisited theory based on free radicals generated by NOX family NADPH oxidases. Exp. Gerontol. 2007, 42, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, J.; Walker, S.J.; Dworakowski, R.; Lakatta, E.G.; Shah, A.M. Involvement of NADPH oxidase in age-associated cardiac remodeling. J. Mol. Cell. Cardiol. 2010, 48, 765–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosser, B.L.; Ward, C.W.; Lederer, W.J. X-ROS signaling: Rapid mechano-chemo transduction in heart. Science 2011, 333, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; Du, M.; Dolence, E.K.; Fang, C.X.; Mayer, G.E.; Ceylan-Isik, A.F.; LaCour, K.H.; Yang, X.; Wilbert, C.J.; Sreejayan, N.; et al. Aging induces cardiac diastolic dysfunction, oxidative stress, accumulation of advanced glycation endproducts and protein modification. Aging Cell 2005, 4, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandy, S.A.; Howlett, S.E. Cardiac excitation-contraction coupling is altered in myocytes from aged male mice but not in cells from aged female mice. Am. J. Physiol.-Heart Circ. Physiol. 2006, 291, H2362–H2370. [Google Scholar] [CrossRef] [PubMed]
- Fares, E.; Howlett, S.E. Effect of age on cardiac excitation-contraction coupling. Clin. Exp. Pharmacol. Physiol. 2010, 37, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Janczewski, A.M.; Lakatta, E.G. Modulation of sarcoplasmic reticulum Ca2+ cycling in systolic and diastolic heart failure associated with aging. Heart Fail. Rev. 2010, 15, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Hummel, S.L.; Salazar, J.B.; Taffet, G.E.; Zieman, S.; Schwartz, J.B. Cardiovascular physiology in the older adults. J. Geriatr. Cardiol. 2015, 12, 196–201. [Google Scholar] [PubMed]
- Kranias, E.G.; Hajjar, R.J. Modulation of cardiac contractility by the phospholamban/SERCA2a regulatome. Circ. Res. 2012, 110, 1646. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, D.H.; Kranias, E.G. Calcium: Phospholamban: A crucial regulator of cardiac contractility. Nat. Rev. Mol. Cell Biol. 2003, 4, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Feridooni, H.A.; Dibb, K.M.; Howlett, S.E. How cardiomyocyte excitation, calcium release and contraction become altered with age. J. Mol. Cell. Cardiol. 2015, 83, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.R.; Treuer, A.V.; Lamirault, G.; Mayo, V.; Cao, Y.; Dulce, R.A.; Hare, J.M. NADPH oxidase-2 inhibition restores contractility and intracellular calcium handling and reduces arrhythmogenicity in dystrophic cardiomyopathy. Am. J. Physiol.-Heart Circ. Physiol. 2014, 307, H710–H721. [Google Scholar] [CrossRef] [PubMed]
- Ago, T.; Matsushima, S.; Kuroda, J.; Zablocki, D.; Kitazono, T.; Sadoshima, J. The NADPH oxidase NOX4 and aging in the heart. Aging 2010, 2, 1012. [Google Scholar] [CrossRef] [PubMed]
- Howlett, S.E. Age-associated changes in excitation-contraction coupling are more prominent in ventricular myocytes from male rats than in myocytes from female rats. Am. J. Physiol.-Heart Circ. Physiol. 2009, 298, H659–H670. [Google Scholar] [CrossRef] [PubMed]
- Howlett, S.E.; Grandy, S.A.; Ferrier, G.R. Calcium spark properties in ventricular myocytes are altered in aged mice. Am. J. Physiol.-Heart Circ. Physiol. 2006, 290, H1566–H1574. [Google Scholar] [CrossRef] [PubMed]
- Weisleder, N.; Ma, J. Altered Ca2+ sparks in aging skeletal and cardiac muscle. Ageing Res. Rev. 2008, 7, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, G.; Borschke, B.; Rueckschloss, U. Ca2+ transients of cardiomyocytes from senescent mice peak late and decay slowly. Cell Calcium 2003, 34, 271–280. [Google Scholar] [CrossRef]
- Donoso, P.; Finkelstein, J.P.; Montecinos, L.; Said, M.; Sanchez, G.; Vittone, L.; Bull, R. Stimulation of NOX2 in isolated hearts reversibly sensitizes RyR2 channels to activation by cytoplasmic calcium. J. Mol. Cell. Cardiol. 2014, 68, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.R.; Treuer, A.; Sun, Q.A.; Stamler, J.S.; Hare, J.M. S-Nitrosylation of cardiac ion channels. J. Cardiovasc. Pharmacol. 2009, 54, 188. [Google Scholar] [CrossRef] [PubMed]
- Treuer, A.V.; Gonzalez, D.R. Nitric oxide synthases, S-nitrosylation and cardiovascular health: From molecular mechanisms to therapeutic opportunities (review). Mol. Med. Rep. 2015, 11, 1555–1565. [Google Scholar] [CrossRef] [PubMed]
- Rueckschloss, U.; Villmow, M.; Klockner, U. NADPH oxidase-derived superoxide impairs calcium transients and contraction in aged murine ventricular myocytes. Exp. Gerontol. 2010, 45, 788–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, L.L.; Li, W.; Lu, Y.; Centracchio, J.; Terentyeva, R.; Koren, G.; Terentyev, D. Redox modification of ryanodine receptors by mitochondria-derived reactive oxygen species contributes to aberrant Ca2+ handling in ageing rabbit hearts. J. Physiol. 2013, 591, 5895–5911. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.C.; Betzenhauser, M.J.; Reiken, S.; Meli, A.C.; Umanskaya, A.; Xie, W.; Shiomi, T.; Zalk, R.; Lacampagne, A.; Marks, A.R. Ryanodine receptor oxidation causes intracellular calcium leak and muscle weakness in aging. Cell. Metab. 2011, 14, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Siwik, D.A.; Lancel, S.; Zhang, J.; Kuster, G.M.; Luptak, I.; Wang, L.; Tong, X.; Kang, Y.J.; Cohen, R.A.; et al. Hydrogen peroxide-mediated SERCA cysteine 674 oxidation contributes to impaired cardiac myocyte relaxation in senescent mouse heart. J. Am. Heart Assoc. Cardiovasc. Cerebrovasc. Dis. 2013, 2, e000184. [Google Scholar] [CrossRef] [PubMed]
- Villmow, M.; Klockner, U.; Heymes, C.; Gekle, M.; Rueckschloss, U. NOS1 induces NADPH oxidases and impairs contraction kinetics in aged murine ventricular myocytes. Basic Res. Cardiol. 2015, 110, 48. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo-Freitas, C.; Dulce, R.A.; Foster, M.W.; Liang, J.; Yamashita, A.M.; Lima-Rosa, F.L.; Thompson, J.W.; Moseley, M.A.; Hare, J.M.; Nogueira, L.; et al. S-Nitrosylation of sarcomeric proteins depresses myofilament Ca2+ sensitivity in intact cardiomyocytes. Antioxid. Redox Signal. 2015, 23, 1017–1034. [Google Scholar] [CrossRef] [PubMed]
- Chakouri, N.; Reboul, C.; Boulghobra, D.; Kleindienst, A.; Nottin, S.; Gayrard, S.; Roubille, F.; Matecki, S.; Lacampagne, A.; Cazorla, O. Stress-induced protein S-glutathionylation and phosphorylation crosstalk in cardiac sarcomeric proteins-Impact on heart function. Int. J. Cardiol. 2018, 258, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Wilder, T.; Ryba, D.M.; Wieczorek, D.F.; Wolska, B.M.; Solaro, R.J. N-acetylcysteine reverses diastolic dysfunction and hypertrophy in familial hypertrophic cardiomyopathy. Am. J. Physiol.-Heart Circ. Physiol. 2015, 309, H1720–H1730. [Google Scholar] [CrossRef] [PubMed]
- Andreollo, N.A.; Santos, E.F.D.; Araujo, M.R.; Lopes, L.R. Rat’s age versus human’s age: What is the relationship? Arq. Bras. Cir. Dig. 2012, 25, 49–51. [Google Scholar] [CrossRef] [PubMed]
- Ten Freyhaus, H.; Huntgeburth, M.; Wingler, K.; Schnitker, J.; Bäumer, A.T.; Vantler, M.; Bekhite, M.M.; Wartenberg, M.; Sauer, H.; Rosenkranz, S. Novel NOX inhibitor VAS2870 attenuates PDGF-dependent smooth muscle cell chemotaxis, but not proliferation. Cardiovasc. Res. 2006, 71, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, J. Impaired S-nitrosylation of the ryanodine receptor caused by xanthine oxidase activity contributes to calcium leak in heart failure. J. Biol. Chem. 2010, 285, 28938–28945. [Google Scholar]
- Dulce, R.A.; Yiginer, O.; Gonzalez, D.R.; Goss, G.; Feng, N.; Zheng, M.; Hare, J.M. Hydralazine and organic nitrates restore impaired excitation-contraction coupling by reducing calcium leak associated with nitroso-redox imbalance. J. Biol. Chem. 2013, 288, 6522–6533. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Correa, G.A.; Cortassa, S.; Stanley, B.; Gao, W.D.; Murphy, A.M. Calcium sensitivity, force frequency relationship and cardiac troponin I: Critical role of PKA and PKC phosphorylation sites. J. Mol. Cell. Cardiol. 2010, 48, 943–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, C.W.; Gaffin, R.D.; Zawieja, D.C.; Muthuchamy, M. Roles of phosphorylation of myosin binding protein-C and troponin I in mouse cardiac muscle twitch dynamics. J. Physiol. 2004, 558, 927–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adults | Aged | p-Value | |
|---|---|---|---|
| N (hearts) | 12 | 19 | - |
| Age (months, range) | 5–6 | 20–24 | - |
| Heart weight (g) | 1.45 ± 0.08 | 2.19 ± 0.13 | 0.00069551 |
| Body weight (g) | 344.4 ± 50 | 702.5 ± 82 | <0.0001 |
| Developed pressure (mmHg) | 108.3 ± 53.5 | 41.4 ± 19.1 | 0.02956983 |
| dP/dtmax (mmHg/s) | 2808 ± 1269 | 1170 ± 815 | 0.0281473 |
| dP/dtmin (−mmHg/s) | −2054 ± 988 | −743 ± 650 | 0.02598013 |
| Coronary perfusion pressure (mmHg) | 60.5 ± 34 | 37.9 ± 9.9 | 0.17102549 |
| Sarcomere Shortening | Adults | Aged | p-Value |
|---|---|---|---|
| N (hearts) | 5 | 4 | - |
| N (cells) | 17 | 22 | - |
| Sarcomere Length (µm) | 1.74 ± 0.066 | 1.73 ± 0.077 | 0.289 |
| Contraction Amplitude (µm) | 0.189 ± 0.056 | 0.17 ± 0.07 | 0.185 |
| % Peak to baseline | 10.8 ± 3.1 | 9.76 ± 4.1 | 0.185 |
| Time to peak 50% (s) | 0.072 ± 0.013 | 0.107 ± 0.04 | <0.005 |
| Time to 50% baseline (s) | 0.379 ± 0.12 | 0.542 ± 0.09 | <0.005 |
| Tau relaxation (s) | 0.193 ± 0.407 | 0.186 ± 0.08 | 0.407 |
| Intracellular Ca2+ | |||
| Diastolic [Ca2+]i (fura ratio) | 1.2 ± 0.18 | 1.06 ± 0.05 | 0.0051 |
| Systolic [Ca2+]i (fura ratio) | 0.20 ± 0.12 | 0.27 ± 0.2 | 0.0916 |
| % Peak to baseline | 17.1 ± 11.1 | 26.7 ± 20.7 | 0.044 |
| Time to peak 50% [Ca2+]i (s) | 0.04 ± 0.005 | 0.05 ± 0.002 | 0.0067 |
| Time to 50% baseline [Ca2+]i (s) | 0.25 ± 0.08 | 0.35 ± 0.01 | <0.005 |
| Tau [Ca2+]i decay (s) | 0.26 ± 0.148 | 0.25 ± 0.06 | 0.420 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valdés, Á.; Treuer, A.V.; Barrios, G.; Ponce, N.; Fuentealba, R.; Dulce, R.A.; González, D.R. NOX Inhibition Improves β-Adrenergic Stimulated Contractility and Intracellular Calcium Handling in the Aged Rat Heart. Int. J. Mol. Sci. 2018, 19, 2404. https://doi.org/10.3390/ijms19082404
Valdés Á, Treuer AV, Barrios G, Ponce N, Fuentealba R, Dulce RA, González DR. NOX Inhibition Improves β-Adrenergic Stimulated Contractility and Intracellular Calcium Handling in the Aged Rat Heart. International Journal of Molecular Sciences. 2018; 19(8):2404. https://doi.org/10.3390/ijms19082404
Chicago/Turabian StyleValdés, Álvaro, Adriana V. Treuer, Guillermo Barrios, Nikol Ponce, Roberto Fuentealba, Raul A. Dulce, and Daniel R. González. 2018. "NOX Inhibition Improves β-Adrenergic Stimulated Contractility and Intracellular Calcium Handling in the Aged Rat Heart" International Journal of Molecular Sciences 19, no. 8: 2404. https://doi.org/10.3390/ijms19082404
APA StyleValdés, Á., Treuer, A. V., Barrios, G., Ponce, N., Fuentealba, R., Dulce, R. A., & González, D. R. (2018). NOX Inhibition Improves β-Adrenergic Stimulated Contractility and Intracellular Calcium Handling in the Aged Rat Heart. International Journal of Molecular Sciences, 19(8), 2404. https://doi.org/10.3390/ijms19082404