Proteomic Signatures Reveal Differences in Stress Response, Antioxidant Defense and Proteasomal Activity in Fertile Men with High Seminal ROS Levels

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Semen Analysis and ROS Levels
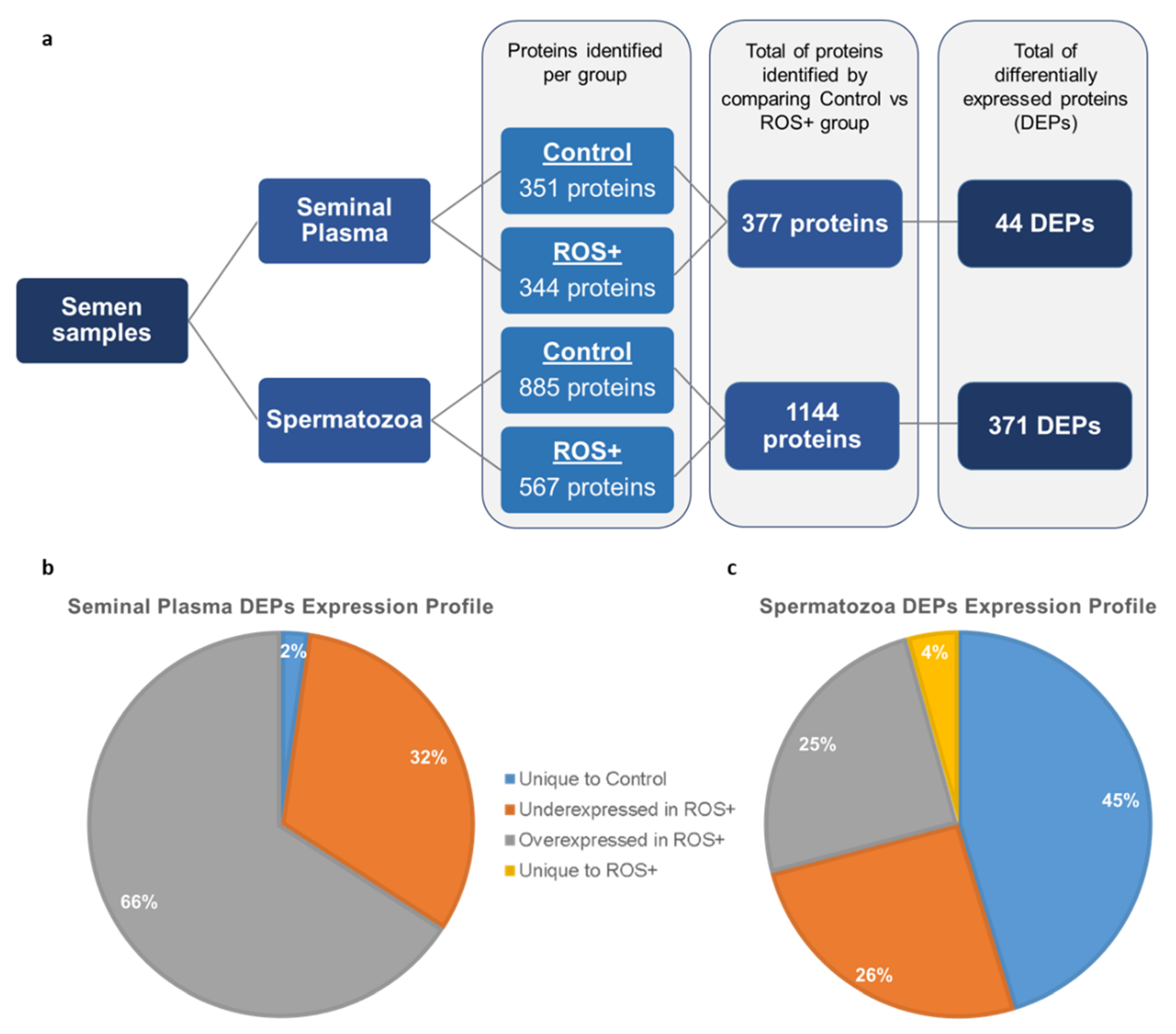
2.2. Global Proteomic Profile of Seminal Plasma and Spermatozoa
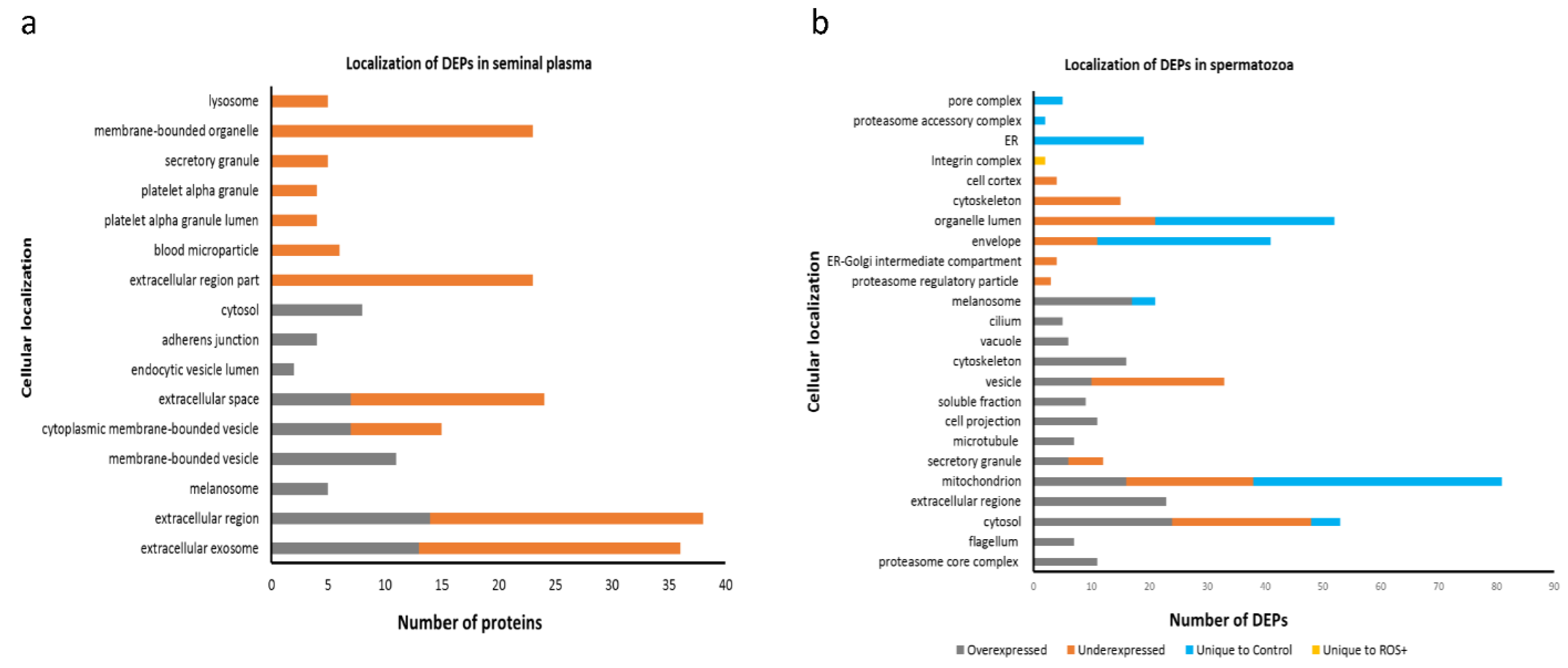
2.3. Functional Annotations and Pathway Analysis
2.4. Upstream Regulators
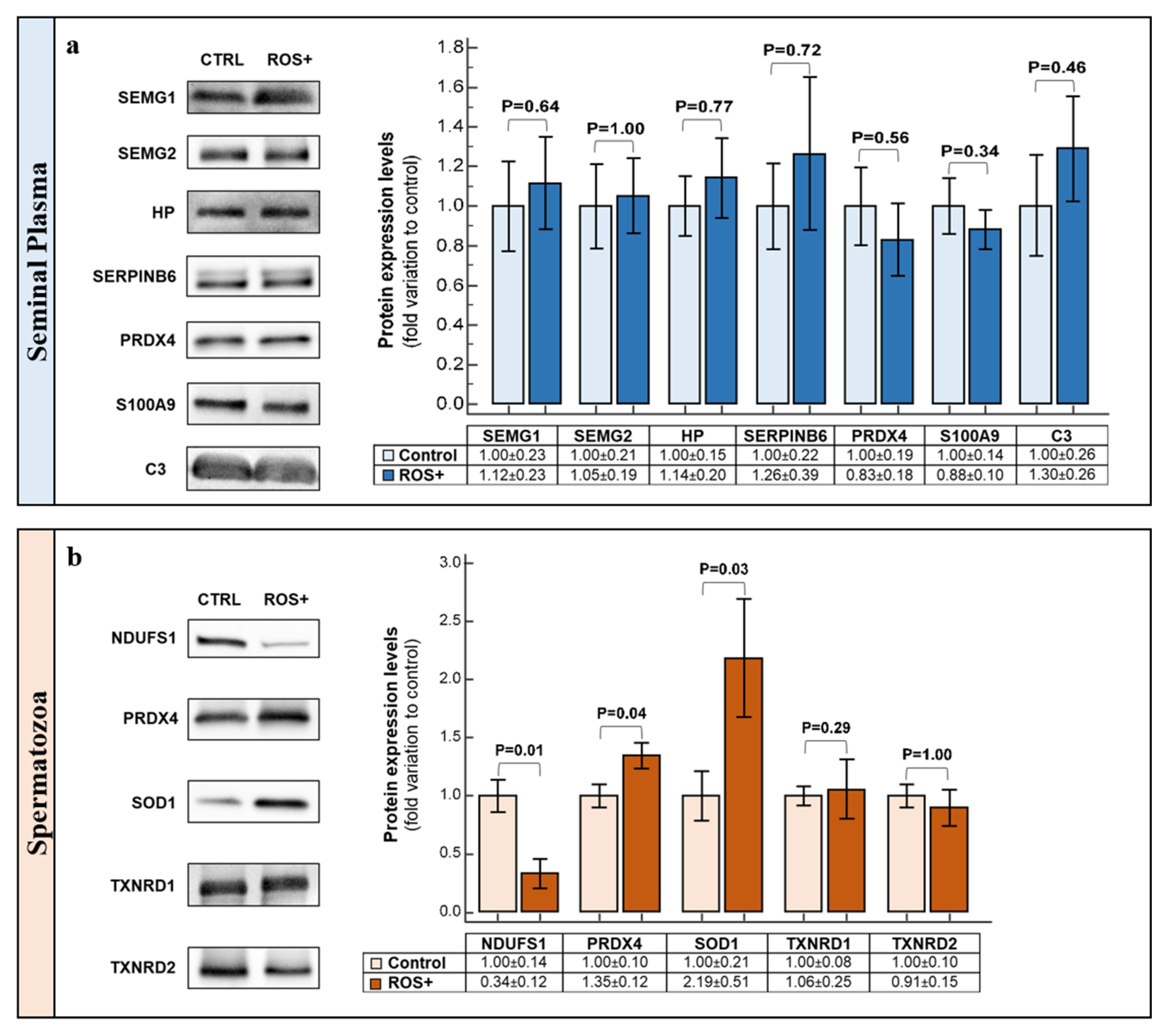
2.5. Western Blot
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Semen Analysis
4.3. Measurement of Reactive Oxygen Species
4.4. Protein Extraction and Quantification
4.5. Quantitative Proteomic Analysis
4.6. Bioinformatic Analysis
4.7. Western Blot
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agarwal, A.; Saleh, R.A.; Bedaiwy, M.A. Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil. Steril. 2003, 79, 829–843. [Google Scholar] [CrossRef] [Green Version]
- Lavranos, G.; Balla, M.; Tzortzopoulou, A.; Syriou, V.; Angelopoulou, R. Investigating ROS sources in male infertility: A common end for numerous pathways. Reprod. Toxicol. 2012, 34, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Agarwal, A. Role of reactive oxygen species in male infertility. Urology 1996, 48, 835–850. [Google Scholar] [CrossRef]
- Agarwal, A.; Sharma, R.K.; Nallella, K.P.; Thomas, A.J.; Alvarez, J.G.; Sikka, S.C. Reactive oxygen species as an independent marker of male factor infertility. Fertil. Steril. 2006, 86, 878–885. [Google Scholar] [CrossRef]
- Panner Selvam, M.K.; Agarwal, A.; Sharma, R.; Samanta, L. Treatment of semen samples with α-chymotrypsin alters the expression pattern of sperm functional proteins—A pilot study. Andrology 2018, 6, 345–350. [Google Scholar] [CrossRef]
- Ko, E.Y.; Sabanegh, E.S., Jr.; Agarwal, A. Male infertility testing: Reactive oxygen species and antioxidant capacity. Fertil. Steril. 2014, 102, 1518–1527. [Google Scholar] [CrossRef]
- Wang, X.; Grammatikakis, N.; Siganou, A.; Calderwood, S.K. Regulation of molecular chaperone gene transcription involves the serine phosphorylation, 14-3-3ε binding, and cytoplasmic sequestration of heat shock factor 1. Mol. Cell. Biol. 2003, 23, 6013–6026. [Google Scholar] [CrossRef] [PubMed]
- Kovac, J.R.; Pastuszak, A.W.; Lamb, D.J. The use of genomics, proteomics, and metabolomics in identifying biomarkers of male infertility. Fertil. Steril. 2013, 99, 998–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremellen, K. Oxidative stress and male infertility—A Clinical Perspective. Hum. Reprod. Update 2008, 14, 243–258. [Google Scholar] [CrossRef]
- Lewis, S.E. Sperm DNA fragmentation and base oxidation. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2014; Volume 791, pp. 103–116. [Google Scholar]
- Simon, L.; Proutski, I.; Stevenson, M.; Jennings, D.; McManus, J.; Lutton, D.; Lewis, S. Sperm DNA damage has a negative association with live-birth rates after IVF. Reprod. Biomed. Online 2013, 26, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.; Bronson, R.; Smith, T.; De Iuliis, G. The source and significance of DNA damage in human spermatozoa; a commentary on diagnostic strategies and straw man fallacies. Mol. Hum. Reprod. 2013, 19, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Bertolla, R.P.; Samanta, L. Sperm proteomics: Potential impact on male infertility treatment. Expert Rev. Proteom. 2016, 13, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Hamada, A.; Sharma, R.; Du Plessis, S.S.; Willard, B.; Yadav, S.P.; Sabanegh, E.; Agarwal, A. Two-dimensional differential in-gel electrophoresis–based proteomics of male gametes in relation to oxidative stress. Fertil. Steril. 2013, 99, 1216–1226.e2. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Agarwal, A.; Mohanty, G.; Hamada, A.J.; Gopalan, B.; Willard, B.; Yadav, S.; Du Plessis, S. Proteomic analysis of human spermatozoa proteins with oxidative stress. Reprod. Biol. Endocrinol. 2013, 11, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayaz, A.; Agarwal, A.; Sharma, R.; Arafa, M.; Elbardisi, H.; Cui, Z. Impact of precise modulation of reactive oxygen species levels on spermatozoa proteins in infertile men. Clin. Proteom. 2015, 12. [Google Scholar] [CrossRef]
- Agarwal, A.; Ahmad, G.; Sharma, R. Reference values of reactive oxygen species in seminal ejaculates using chemiluminescence assay. J. Assist. Reprod. Genet. 2015, 32, 1721–1729. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Aitken, R. Oxidative stress and the etiology of male infertility. J. Assist. Reprod. Genet. 2016, 33, 1691–1692. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Mens Health 2014, 32, 1–17. [Google Scholar] [CrossRef]
- Agarwal, A.; Sharma, R.K.; Sharma, R.; Assidi, M.; Abuzenadah, A.M.; Alshahrani, S.; Durairajanayagam, D.; Sabanegh, E. Characterizing semen parameters and their association with reactive oxygen species in infertile men. Reprod. Biol. Endocrinol. 2014, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, M.H.; Sharma, R.K.; Thornton, J.; Mascha, E.; Abdel-Hafez, M.A.; Thomas, A.J.; Agarwal, A. Relationship between ROS production, apoptosis and DNA denaturation in spermatozoa from patients examined for infertility. Hum. Reprod. 2004, 19, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.; Sharma, R.; Agarwal, A. Proteomic analysis of mature and immature ejaculated spermatozoa from fertile men. Asian J. Androl. 2016, 18, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Chiriva-Internati, M.; Gagliano, N.; Donetti, E.; Costa, F.; Grizzi, F.; Franceschini, B.; Albani, E.; Levi-Setti, P.E.; Gioia, M.; Jenkins, M. Sperm protein 17 is expressed in the sperm fibrous sheath. J. Transl. Med. 2009, 7, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grizzi, F.; Chiriva–Internati, M.; Franceschini, B.; Hermonat, P.L.; Soda, G.; Lim, S.H.; Dioguardi, N. Immunolocalization of sperm protein 17 in human testis and ejaculated spermatozoa. J. Histochem. Cytochem. 2003, 51, 1245–1248. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.; Lundwall, Å.; Malm, J. The semenogelins: Proteins with functions beyond reproduction? Cell. Mol. Life Sci. 2006, 63, 2886–2888. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.; Linse, S.; Frohm, B.; Lundwall, Å.; Johan, M. Semenogelins I and II bind zinc and regulate the activity of prostate-specific antigen. Biochem. J. 2005, 387, 447–453. [Google Scholar] [CrossRef]
- Prassas, I.; Eissa, A.; Poda, G.; Diamandis, E.P. Unleashing the therapeutic potential of human kallikrein-related serine proteases. Nat. Rev. Drug Discov. 2015, 14, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Silverman, G.A.; Whisstock, J.; Askew, D.; Pak, S.; Luke, C.; Cataltepe, S.; Irving, J.; Bird, P. Human clade B serpins (ov-serpins) belong to a cohort of evolutionarily dispersed intracellular proteinase inhibitor clades that protect cells from promiscuous proteolysis. Cell. Mol. Life Sci. 2004, 61, 301–325. [Google Scholar] [CrossRef]
- Melamed-Frank, M.; Lache, O.; Enav, B.I.; Szafranek, T.; Levy, N.S.; Ricklis, R.M.; Levy, A.P. Structure-function analysis of the antioxidant properties of haptoglobin. Blood 2001, 98, 3693–3698. [Google Scholar] [CrossRef] [Green Version]
- Kapralov, A.A.; Vlasova, I.I.; Feng, W.; Maeda, A.; Walson, K.; Tyurin, V.A.; Huang, Z.; Aneja, R.K.; Carcillo, J.; Bayir, H. Peroxidase activity of hemoglobin/haptoglobin complexes: Covalent aggregation and oxidative stress in plasma and macrophages. J. Biol. Chem. 2009, 284, 30395–30407. [Google Scholar] [CrossRef]
- O’Flaherty, C.; Rico de Souza, A. Hydrogen peroxide modifies human sperm peroxiredoxins in a dose-dependent manner. Biol. Reprod. 2011, 84, 238–247. [Google Scholar] [CrossRef]
- O’Flaherty, C. The enzymatic antioxidant system of human spermatozoa. Adv. Androl. 2014, 2014, 1–15. [Google Scholar]
- Srikrishna, G. S100A8 and S100A9: New insights into their roles in malignancy. J. Innate Immun. 2012, 4, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Foell, D.; Wittkowski, H.; Vogl, T.; Roth, J. S100 proteins expressed in phagocytes: A novel group of damage-associated molecular pattern molecules. J. Leukoc. Biol. 2007, 81, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Ingram, S.; Diotallevi, M. Reactive oxygen species: Rapid fire in inflammation. Biochemist 2017, 39, 30–33. [Google Scholar]
- Antoniassi, M.P.; Intasqui, P.; Camargo, M.; Zylbersztejn, D.S.; Carvalho, V.M.; Cardozo, K.H.; Bertolla, R.P. Analysis of the functional aspects and seminal plasma proteomic profile of sperm from smokers. BJU Int. 2016, 118, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Benedyk, M.; Sopalla, C.; Nacken, W.; Bode, G.; Melkonyan, H.; Banfi, B.; Kerkhoff, C. HaCaT keratinocytes overexpressing the S100 proteins S100A8 and S100A9 show increased NADPH oxidase and NF-κB activities. J. Investig. Dermatol. 2007, 127, 2001–2011. [Google Scholar] [CrossRef] [PubMed]
- Samanta, L.; Parida, R.; Dias, T.R.; Agarwal, A. The enigmatic seminal plasma: A proteomics insight from ejaculation to fertilization. Reprod. Biol. Endocrinol. 2018, 16, 41. [Google Scholar] [CrossRef]
- Mastellos, D.; Lambris, J.D. Complement: More than a ‘guard’against invading pathogens? Trends Immunol. 2002, 23, 485–491. [Google Scholar] [CrossRef]
- Andrews, E.; Feldhoff, P.; Feldhoff, R.; Lassiter, H. Comparative effects of cytokines and cytokine combinations on complement component C3 secretion by HepG2 cells. Cytokine 2003, 23, 164–169. [Google Scholar] [CrossRef]
- Chowdhury, N.; Kamada, M.; Takikawa, M.; Mori, H.; Gima, H.; Aono, T. Complement-inhibiting activity of human seminal plasma and semen quality. Arch. Androl. 1996, 36, 109–118. [Google Scholar] [CrossRef]
- Janssen, R.J.; Nijtmans, L.G.; Van Den Heuvel, L.P.; Smeitink, J.A. Mitochondrial complex I: Structure, function and pathology. J. Inherit. Metab. Dis. 2006, 29, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Mimaki, M.; Wang, X.; McKenzie, M.; Thorburn, D.R.; Ryan, M.T. Understanding mitochondrial complex I assembly in health and disease. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 851–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Sampath, V.; Garland, J.S.; Helbling, D.; Dimmock, D.; Mulrooney, N.P.; Simpson, P.M.; Murray, J.C.; Dagle, J.M. Antioxidant response genes sequence variants and BPD susceptibility in VLBW infants. Pediatr. Res. 2015, 77, 477. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Kil, I.S. Multiple functions and regulation of mammalian peroxiredoxins. Annu. Rev. Biochem. 2017, 86, 749–775. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A.; Lu, J. Thioredoxin and thioredoxin reductase: Current research with special reference to human disease. Biochem. Biophys. Res. Commun. 2010, 396, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Gong, S.; Gabriel, M.C.S.; Zini, A.; Chan, P.; O’Flaherty, C. Low amounts and high thiol oxidation of peroxiredoxins in spermatozoa from infertile men. J. Androl. 2012, 33, 1342–1351. [Google Scholar] [CrossRef]
- Grune, T.; Merker, K.; Sandig, G.; Davies, K.J. Selective degradation of oxidatively modified protein substrates by the proteasome. Biochem. Biophys. Res. Commun. 2003, 305, 709–718. [Google Scholar] [CrossRef]
- Hirano, Y.; Hendil, K.B.; Yashiroda, H.; Iemura, S.-I.; Nagane, R.; Hioki, Y.; Natsume, T.; Tanaka, K.; Murata, S. A heterodimeric complex that promotes the assembly of mammalian 20S proteasomes. Nature 2005, 437, 1381–1385. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.; Sharma, R. Leukocytospermia Quantitation (ENDTZ) Test. In Andrological Evaluation of Male Infertility; Springer: Cham, Switzerland, 2016; pp. 69–72. [Google Scholar]
- Agarwal, A.; Gupta, S.; Sharma, R. Reactive oxygen species (ROS) measurement. In Andrological Evaluation of Male Infertility; Springer: Cham, Switzerland, 2016; pp. 155–163. [Google Scholar]
- Agarwal, A.; Ayaz, A.; Samanta, L.; Sharma, R.; Assidi, M.; Abuzenadah, A.M.; Sabanegh, E. Comparative proteomic network signatures in seminal plasma of infertile men as a function of reactive oxygen species. Clin. Proteom. 2015, 12, 23. [Google Scholar] [CrossRef]
- Boyle, E.I.; Weng, S.; Gollub, J.; Jin, H.; Botstein, D.; Cherry, J.M.; Sherlock, G. GO::TermFinder—Open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 2004, 20, 3710–3715. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, V.N.; Perlman, D.H.; Costello, C.E.; McComb, M.E. Software tool for researching annotations of proteins: Open-source protein annotation software with data visualization. Anal. Chem. 2009, 81, 9819–9823. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameter | WHO 1 | Control | ROS+ | p-Value |
|---|---|---|---|---|
| Volume (mL) | >1.5 | 4.24 ± 0.67 | 3.76 ± 0.72 | 0.4384 |
| pH | 7.6–8 | 7.66 ± 0.07 | 7.60 ± 0.05 | 0.3333 |
| Sperm motility (%) | >40 | 55 ± 3 | 58 ± 6 | 0.5035 |
| Sperm concentration (106/mL) | >15 | 90.95 ± 15.59 | 75.02 ± 12.87 | 0.6221 |
| Total sperm count (106) | >39 | 359.50 ± 63.86 | 254.95 ± 49.67 | 0.1809 |
| Round cells (106/mL) | <1 | 0.82 ± 0.27 | 1.68 ± 0.76 | 0.6221 |
| Leukocytes (106/mL) | <1 | 0.04 ± 0.04 | 0.04 ± 0.04 | 0.9539 |
| ROS levels (RLU/sec/106 sperm) | - | 46.62 ± 9.67 | 1053.21 ± 441.43 | 0.0001 |
| Protein | Abundance | NSAF Ratio | Expression Profile | p-Value | |
|---|---|---|---|---|---|
| Control | ROS+ | ||||
| SEMG1 | High | High | 0.26 | UE in ROS+ | 0.00074 |
| SEMG2 | High | High | 0.26 | UE in ROS+ | 0.00023 |
| HP | Very Low | Low | 9.03 | OE in ROS+ | 0.00349 |
| PRDX4 | Very Low | Low | 3.39 | OE in ROS+ | 0.00099 |
| SERPINB6 | Low | Low | 2.68 | OE in ROS+ | 0.00424 |
| S100A9 | Very Low | Medium | 3.77 | OE in ROS+ | 0.01707 |
| C3 | Very Low | Medium | 17.22 | OE in ROS+ | 0.00210 |
| Protein | Abundance | NSAF Ratio | Expression Profile | p-Value | |
|---|---|---|---|---|---|
| Control | ROS+ | ||||
| NDUFS1 | Medium | Very Low | 0.02 | UE in ROS+ | 0.00004 |
| PRDX4 | Low | Medium | 4.48 | OE in ROS+ | 0.00134 |
| SOD1 | Low | Medium | 3.99 | OE in ROS+ | 0.02830 |
| TXNRD1 | - | Very Low | - | Unique to ROS+ | 0.00006 |
| TXNRD2 | Very Low | Medium | 10.95 | OE in ROS+ | 0.03640 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, T.R.; Samanta, L.; Agarwal, A.; Pushparaj, P.N.; Panner Selvam, M.K.; Sharma, R. Proteomic Signatures Reveal Differences in Stress Response, Antioxidant Defense and Proteasomal Activity in Fertile Men with High Seminal ROS Levels. Int. J. Mol. Sci. 2019, 20, 203. https://doi.org/10.3390/ijms20010203
Dias TR, Samanta L, Agarwal A, Pushparaj PN, Panner Selvam MK, Sharma R. Proteomic Signatures Reveal Differences in Stress Response, Antioxidant Defense and Proteasomal Activity in Fertile Men with High Seminal ROS Levels. International Journal of Molecular Sciences. 2019; 20(1):203. https://doi.org/10.3390/ijms20010203
Chicago/Turabian StyleDias, Tânia R., Luna Samanta, Ashok Agarwal, Peter N. Pushparaj, Manesh Kumar Panner Selvam, and Rakesh Sharma. 2019. "Proteomic Signatures Reveal Differences in Stress Response, Antioxidant Defense and Proteasomal Activity in Fertile Men with High Seminal ROS Levels" International Journal of Molecular Sciences 20, no. 1: 203. https://doi.org/10.3390/ijms20010203
APA StyleDias, T. R., Samanta, L., Agarwal, A., Pushparaj, P. N., Panner Selvam, M. K., & Sharma, R. (2019). Proteomic Signatures Reveal Differences in Stress Response, Antioxidant Defense and Proteasomal Activity in Fertile Men with High Seminal ROS Levels. International Journal of Molecular Sciences, 20(1), 203. https://doi.org/10.3390/ijms20010203