The Elongation Factor GmEF4 Is Involved in the Response to Drought and Salt Tolerance in Soybean



 and
and
Abstract
:1. Introduction
2. Results
2.1. Identification, Physical Locations, and Conserved Domains of Soybean EF1αs
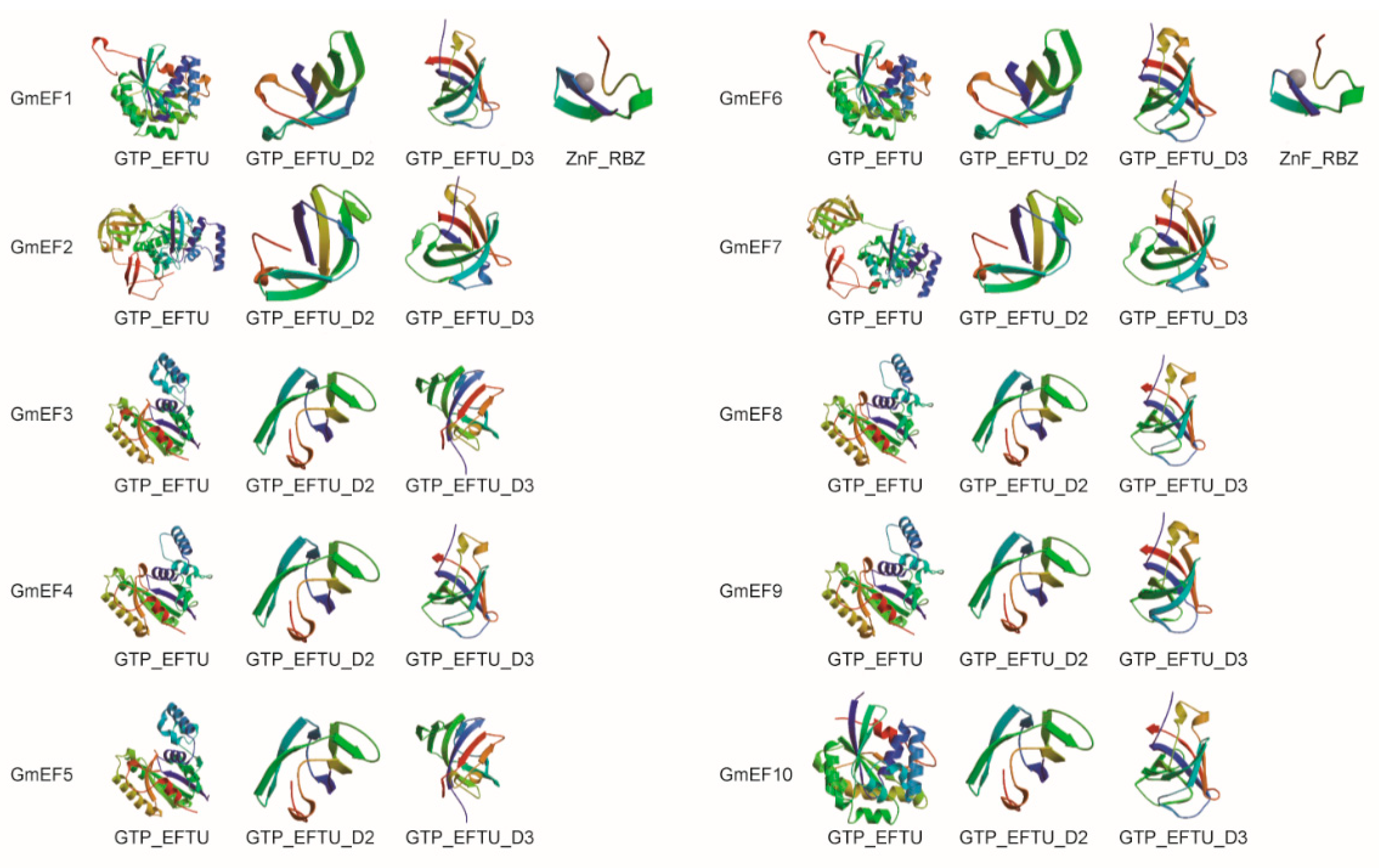
2.2. The Predicted Structures of Domains in Soybean EF1αs
2.3. Phylogenetic and Gene Structural Analysis
2.4. Expression Patterns of Soybean EF1αs
2.5. Soybean EF1αs Contain Various cis-Elements
2.6. GmEF4 Localized in Cell Membranes
2.7. Responses of Soybean EF1αs to Drought, Salt, and ABA Stresses
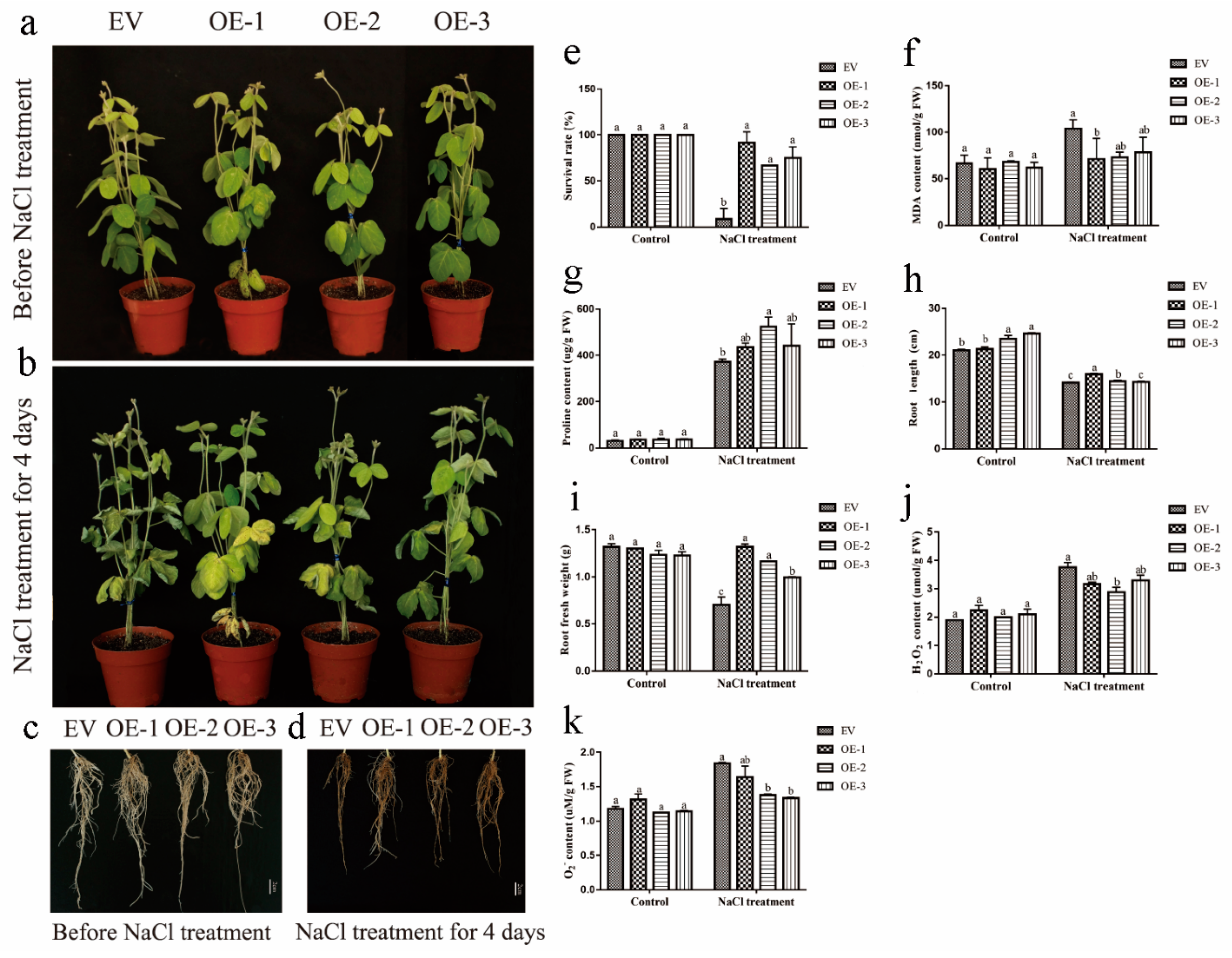
2.8. Overexpression of GmEF4 Significantly Improved Drought Tolerance in Soybean Hairy Roots
2.9. Overexpression of GmEF4 Significantly Improved Salt Tolerance in Soybean Hairy Roots
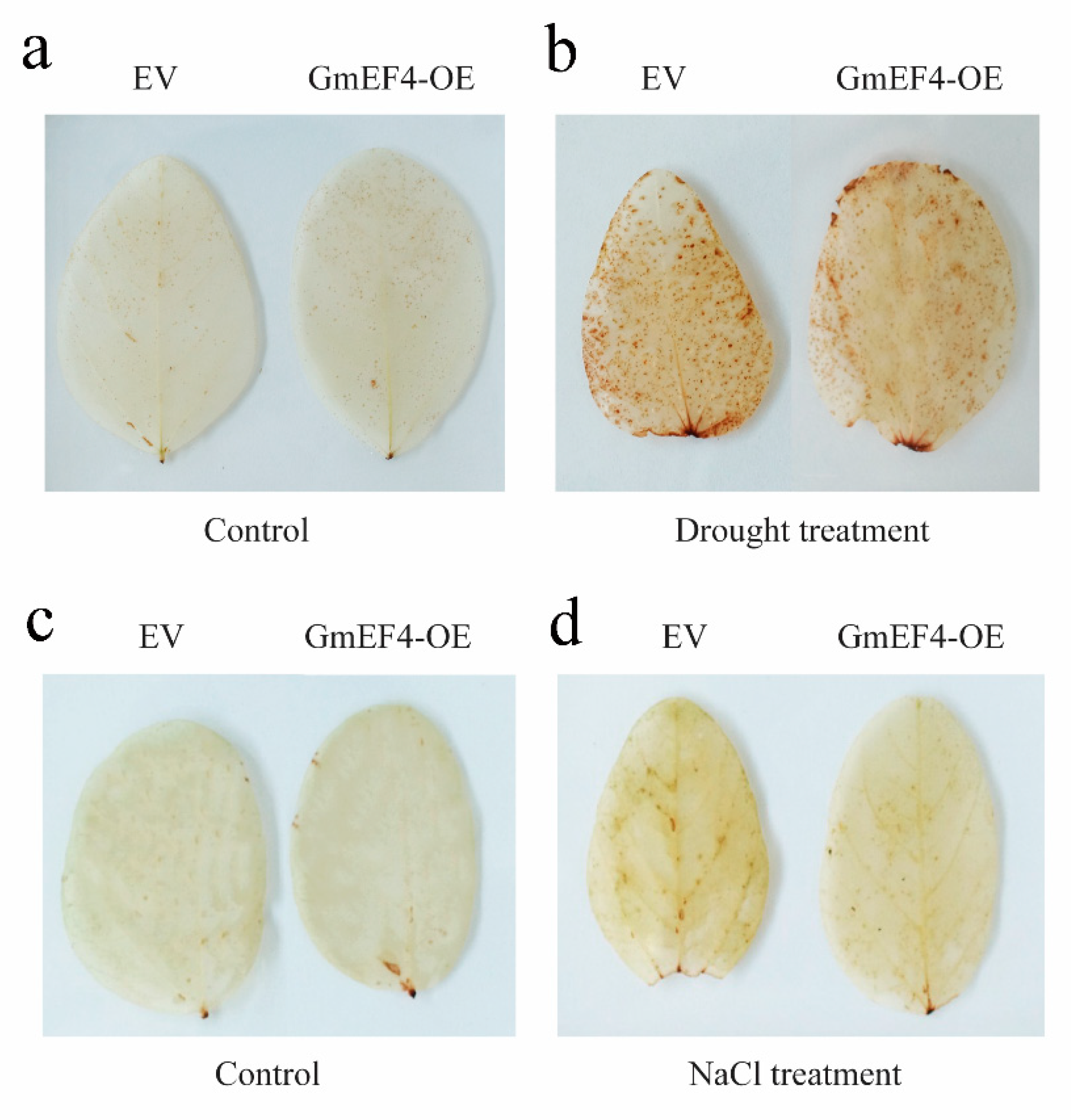
2.10. Overexpression of GmEF4 Reduced the Content of H2O2 under Drought and Salt Stresses
3. Discussion
4. Materials and Methods
4.1. Genomic Location of Soybean EF1α Loci
4.2. Analysis of Gene Structure and cis-Acting Elements
4.3. Genomic and Phylogenetic Relationships and Expression Patterns
4.4. RNA Extraction and qRT-PCR Assays
4.5. Agrobacterium rhizogenes-Mediated Transformation of Soybean Hairy Roots
4.6. Drought and Salt Tolerance Analyses
4.7. Subcellular Localization
4.8. Measurements of Fresh Weight and Main Length
4.9. Measurements of MDA Content, Proline Content, H2O2 Content, and O2− Content
4.10. DAB Staining
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EF1α | elongation factor 1α |
| pI | isoelectric points |
| ABREs | ABA-responsive elements |
| DREs | dehydration-responsive elements |
| EF-Tu | elongation factor thermo unstable |
| ABA | abscisic acid |
| qRT-PCR | quantitative real-time PCR |
| CAMs | calmodulins |
| GFP | green fluorescent protein |
| MDA | malondialdehyde |
| Pro | proline |
| DAF | days after flowering |
References
- Zheng, J.; Liu, H.; Wang, Y.; Wang, L.; Chang, X.; Jing, R.; Hao, C.; Zhang, X. TEF-7A, a transcript elongation factor gene, influences yield-related traits in bread wheat (Triticum aestivum L.). J. Exp. Bot. 2014, 65, 5351–5365. [Google Scholar] [CrossRef] [PubMed]
- Merrick, W.C. Mechanism and regulation of eukaryotic protein synthesis. Microbiol. Rev. 1992, 56, 291–315. [Google Scholar] [PubMed]
- Kazuki, S.; Paul, S. Omnipotent role of archaeal elongation factor 1 alpha (EF1α in translational elongation and termination, and quality control of protein synthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 19242–19247. [Google Scholar]
- Gonen, H.; Dickman, D.; Schwartz, A.L.; Ciechanover, A. Protein synthesis elongation factor EF-1 alpha is an isopeptidase essential for ubiquitin-dependent degradation of certain proteolytic substrates. Adv. Exp. Med. Biol. 1996, 389, 209–219. [Google Scholar] [PubMed]
- Dunn, M.A.; Morris, A.; Jack, P.L.; Hughes, M.A. A low-temperature-responsive translation elongation factor 1 alpha from barley (Hordeum vulgare L.). Plant Mol. Biol. 1993, 23, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Berberich, T.; Sugawara, K.; Harada, M.; Kusano, T. Molecular cloning, characterization and expression of an elongation factor 1 alpha gene in maize. Plant Mol. Biol. 1995, 29, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Momcilović, I.; Prasad, P.V.V. Roles of Protein Synthesis Elongation Factor EF-Tu in Heat Tolerance in Plants. J. Exp. Bot. 2012, 835836, 8. [Google Scholar] [CrossRef]
- Rathod, D.P.; Brestic, M.; Shao, H.B. Chlorophyll a fluorescence determines the drought resistance capabilities in two varieties of mycorrhized and non-mycorrhized Glycine max Linn. Afr. J. Microbil. Res. 2011, 5, 4197–4206. [Google Scholar] [CrossRef]
- Hussain, S.; Iqbal, N.; Rahman, T.; Liu, T.; Brestic, M.; Safdar, M.E.; Asghar, M.A.; Farooq, M.U.; Shafiq, I.; Ali, A.; et al. Shade effect on carbohydrates dynamics and stem strength of soybean genotypes. Environ. Exp. Bot. 2019, 162, 374–382. [Google Scholar] [CrossRef]
- Rose, A.B. Intron-mediated regulation of gene expression. Curr. Top. Microbiol. Immunol. 2008, 326, 277–290. [Google Scholar]
- Chen, Y.; Yang, X.; He, K.; Liu, M.; Li, J.; Gao, Z.; Lin, Z.; Zhang, Y.; Wang, X.; Qiu, X.; et al. The MYB Transcription Factor Superfamily of Arabidopsis: Expression Analysis and Phylogenetic Comparison with the Rice MYB Family. Plant Mol. Biol. 2006, 60, 107–124. [Google Scholar]
- Li, W.; Cui, X.; Meng, Z.; Huang, X.; Xie, Q.; Wu, H.; Jin, H.; Zhang, D.; Liang, W. Transcriptional regulation of Arabidopsis MIR168a and argonaute1 homeostasis in abscisic acid and abiotic stress responses. Plant Physiol. 2012, 158, 1279–1292. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.S.; Xia, L.Q.; Chen, M.; Cheng, X.G.; Zhang, R.Y.; Li, L.C.; Zhao, Y.X.; Lu, Y.; Ni, Z.Y.; Liu, L.; et al. Isolation and molecular characterization of the Triticum aestivum L. ethylene-responsive factor 1 (TaERF1) that increases multiple stress tolerance. Plant Mol. Biol. 2007, 65, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.S.; Chen, M.; Li, L.C.; Ma, Y.Z. Functions and application of the AP2/ERF transcription factor family in crop improvement. J. Integr. Plant Biol. 2011, 53, 570–585. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Tong, J.F.; He, X.L.; Xu, Z.L.; Xu, L.; Wei, P.P.; Huang, Y.H.; Brestic, M.; Ma, H.X.; Shao, H.B. A Novel Soybean Intrinsic Protein Gene, GmTIP2;3, Involved in Responding to Osmotic Stress. Front. Plant Sci. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, J.A.; Dixon, P.; Granger, C.; Grant, D.; Clark, L.; Doyle, J.J.; Shoemaker, R.C. Mining EST databases to resolve evolutionary events in major crop species. Genome 2004, 47, 868–876. [Google Scholar] [CrossRef] [Green Version]
- Blanc, G.; Barakat, A.; Guyot, R.; Cooke, R.; Delseny, M. Extensive duplication and reshuffling in the Arabidopsis genome. Plant Cell 2000, 12, 1093–1102. [Google Scholar] [CrossRef]
- Kaur, K.J.; Ruben, L. Protein translation elongation factor-1 alpha from Trypanosoma brucei binds calmodulin. J. Biol. Chem. 1994, 269, 23045–23050. [Google Scholar]
- Joshi, R.L.; Ravel, J.M.; Haenni, A.L. Interaction of turnip yellow mosaic virus Val-RNA with eukaryotic elongation factor EF-1 [alpha]. Search for a function. EMBO J. 1986, 5, 1143–1148. [Google Scholar] [CrossRef]
- Matsuda, D.; Yoshinari, S.; Dreher, T.W. eEF1A binding to aminoacylated viral RNA represses minus strand synthesis by TYMV RNA-dependent RNA polymerase. J. Virol. 2004, 321, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.S.; Seok, H.; Kwon, T.K.; Min, D.S.; Ahn, B.H.; Lee, Y.H.; Suh, J.W.; Kim, J.W.; Iwashita, S.; Omori, A.; et al. Interaction of elongation factor-1alpha and pleckstrin homology domain of phospholipase C-gamma 1 with activating its activity. J. Biol. Chem. 2002, 277, 19697–19702. [Google Scholar] [CrossRef] [PubMed]
- Gungabissoon, R.A.; Khan, S.; Hussey, P.J.; Maciver, S.K. Interaction of elongation factor 1α from Zea mays (ZmEF-1α) with F-actin and interplay with the maize actin severing protein, ZmADF3. Cytoskeleton 2010, 49, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Urška, B.; Fu, J.; Miranda, B.; Vara Prasad, P.V.; Zoran, R. Heat tolerance and expression of protein synthesis elongation factors, EF-Tu and EF-1α, in spring wheat. Funct. Plant Biol. 2009, 36, 234–241. [Google Scholar]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Xu, Z.S.; Lu, P.P.; Hu, D.; Chen, M.; Li, L.C.; Ma, Y.Z. A wheat PI4K gene whose product possesses threonine autophophorylation activity confers tolerance to drought and salt in Arabidopsis. J. Exp. Bot. 2013, 64, 2915–2927. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K.; et al. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 2011, 68, 302–313. [Google Scholar] [CrossRef]
- Wang, F.; Chen, H.W.; Li, Q.T.; Wei, W.; Li, W.; Zhang, W.K.; Ma, B.; Bi, Y.D.; Lai, Y.C.; Liu, X.L.; et al. GmWRKY27 interacts with GmMYB174 to reduce expression of GmNAC29 for stress tolerance in soybean plants. Plant J. 2015, 83, 224–236. [Google Scholar] [CrossRef]
- Du, Y.T.; Zhao, M.J.; Wang, C.T.; Gao, Y.; Wang, Y.X.; Liu, Y.W.; Chen, M.; Chen, J.; Zhou, Y.B.; Xu, Z.S. Identification and characterization of GmMYB118 responses to drought and salt stress. BMC Plant Biol. 2018, 18, 320. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Cui, X.Y.; Gao, Y.; Guo, J.; Yu, T.F.; Zheng, W.J.; Liu, Y.W.; Chen, J.; Xu, Z.S.; Ma, Y.Z. BES/BZR Transcription Factor TaBZR2 Positively Regulates Drought Responses by Activation of TaGST1. Plant Physiol. 2019, 180, 1605–1620. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID | Amino Acids | MW (Da) | pI | Chromosome | Domain I | Domain II | Domain III |
|---|---|---|---|---|---|---|---|---|
| GmEF1 | Glyma08g12790 | 787 | 85,982.9 | 6.43 | 8 | 360–579 | 605–669 | 678–782 |
| GmEF2 | Glyma08g05570 | 504 | 56,282.4 | 4.89 | 8 | 82–302 | 322–389 | 395–502 |
| GmEF3 | Glyma17g23900 | 447 | 49,387.0 | 9.56 | 17 | 9–227 | 248–314 | 322–429 |
| GmEF4 | Glyma05g24110 | 447 | 49,254.9 | 9.58 | 5 | 9–227 | 248–314 | 322–429 |
| GmEF5 | Glyma05g11630 | 447 | 49,347.0 | 9.56 | 5 | 9–227 | 248–314 | 322–429 |
| GmEF6 | Glyma05g29675 | 792 | 86,366.4 | 6.9 | 5 | 365–584 | 610–674 | 683–787 |
| GmEF7 | Glyma05g34120 | 479 | 53,359.7 | 5.16 | 5 | 57–277 | 297–364 | 370–477 |
| GmEF8 | Glyma19g07240 | 447 | 49,270.9 | 9.58 | 19 | 9–227 | 248–314 | 322–429 |
| GmEF9 | Glyma10g35700 | 448 | 49,491.1 | 9.59 | 10 | 9–227 | 248–314 | 322–429 |
| GmEF10 | Glyma16g07350 | 447 | 49,397.1 | 9.56 | 16 | 9–227 | 248–314 | 322–429 |
| Genes | GmEF1 | GmEF2 | GmEF3 | GmEF4 | GmEF5 | GmEF6 | GmEF7 | GmEF8 | GmEF9 | GmEF10 |
|---|---|---|---|---|---|---|---|---|---|---|
| ABRE | 1 | 1 | 7 | 6 | 6 | 1 | 1 | 8 | 6 | 1 |
| DRE | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| LTRE | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 2 | 1 | 0 |
| MYB | 13 | 11 | 16 | 7 | 9 | 5 | 16 | 10 | 13 | 8 |
| MYC | 16 | 11 | 10 | 16 | 12 | 4 | 12 | 10 | 10 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Ma, J.; Zheng, J.-C.; Chen, J.; Chen, M.; Zhou, Y.-B.; Fu, J.-D.; Xu, Z.-S.; Ma, Y.-Z. The Elongation Factor GmEF4 Is Involved in the Response to Drought and Salt Tolerance in Soybean. Int. J. Mol. Sci. 2019, 20, 3001. https://doi.org/10.3390/ijms20123001
Gao Y, Ma J, Zheng J-C, Chen J, Chen M, Zhou Y-B, Fu J-D, Xu Z-S, Ma Y-Z. The Elongation Factor GmEF4 Is Involved in the Response to Drought and Salt Tolerance in Soybean. International Journal of Molecular Sciences. 2019; 20(12):3001. https://doi.org/10.3390/ijms20123001
Chicago/Turabian StyleGao, Yuan, Jian Ma, Jia-Cheng Zheng, Jun Chen, Ming Chen, Yong-Bin Zhou, Jin-Dong Fu, Zhao-Shi Xu, and You-Zhi Ma. 2019. "The Elongation Factor GmEF4 Is Involved in the Response to Drought and Salt Tolerance in Soybean" International Journal of Molecular Sciences 20, no. 12: 3001. https://doi.org/10.3390/ijms20123001
APA StyleGao, Y., Ma, J., Zheng, J. -C., Chen, J., Chen, M., Zhou, Y. -B., Fu, J. -D., Xu, Z. -S., & Ma, Y. -Z. (2019). The Elongation Factor GmEF4 Is Involved in the Response to Drought and Salt Tolerance in Soybean. International Journal of Molecular Sciences, 20(12), 3001. https://doi.org/10.3390/ijms20123001