Mesenchymal Stem Cells Empowering Tendon Regenerative Therapies




Abstract
:1. Introduction
2. Tendon Pathophysiology
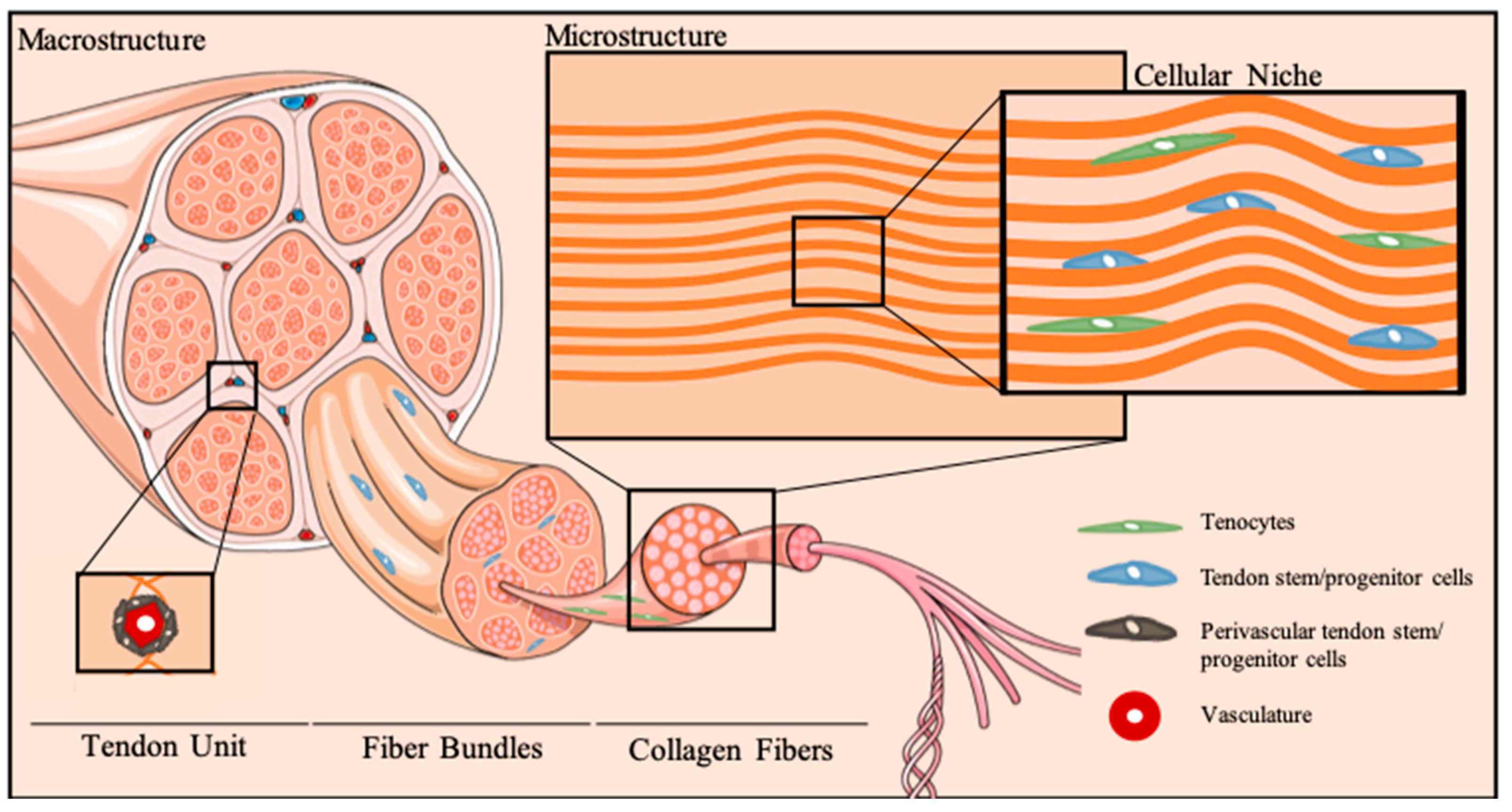
2.1. A Snapshot on Tendon Cellular Niche
2.2. Tendinopathies: A Painful Global Burden and the Need for Novel Therapies
3. Cell-Based Therapies for Tendon Applications
3.1. General Considerations
3.2. Main Challenges
3.3. Mesenchymal Stem Cells for Tendon Regenerative Therapies
3.3.1. Tendon Stem Cells
3.3.2. Mesenchymal Stem Cells from Non-Tendon Tissues
4. Conclusions and Perspectives
Funding
Conflicts of Interest
Abbreviations
| AFSCs | Amniotic fluid stem cells |
| ASCs | Adipose-derived stem cells |
| bFGF | Basic Fibroblast Growth Factor |
| BM-MSCs | Bone marrow-derived mesenchymal stem cells |
| BMP | Bone Morphogenetic Protein |
| ECM | Extracellular matrix |
| EGF | Endothelial growth factor |
| GBD | Global Burden of Disease |
| IGF | Insulin-like Growth Factor |
| IL | Interleukin |
| iPSCs | Induced pluripotent stem cells |
| MMPs | Matrix metalloproteinases |
| MSCs | Mesenchymal stem cells |
| PDGF | Platelet-derived Growth Factor |
| PRHd | Platelet-rich hemoderivatives |
| PRP | Platelet-rich plasma |
| TDSCs | Tendon-derived stem cells |
| TGF | Transforming Growth Factor |
| TSPCs | Tendon stem/progenitor cells |
References
- Bely, A.E.; Nyberg, K.G. Evolution of animal regeneration: Re-emergence of a field. Trends Ecol. Evol. 2010, 25, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Mendes, B.B.; Gómez-Florit, M.; Babo, P.S.; Domingues, R.; Reis, R.L.; Gomes, M.E. Blood derivatives awaken in regenerative medicine strategies to modulate wound healing. Adv. Drug Deliv. Rev. 2018, 12, 376–393. [Google Scholar] [CrossRef] [PubMed]
- Nichols, A.E.C.; Best, K.T.; Loiselle, A.E. The cellular basis of fibrotic tendon healing: Challenges and opportunities. Transl. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Atala, A.; Irvine, D.J.; Moses, M.; Shaunak, S. Wound Healing Versus Regeneration: Role of the Tissue Environment in Regenerative Medicine. MRS Bull. 2010, 35. [Google Scholar] [CrossRef] [PubMed]
- Beredjiklian, P.K.; Favata, M.; Cartmell, J.S.; Flanagan, C.L.; Crombleholme, T.M.; Soslowsky, L.J. Regenerative versus reparative healing in tendon: A study of biomechanical and histological properties in fetal sheep. Ann. Biomed. Eng. 2003, 31, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M.W.; O’Kane, S. Scar-free healing: From embryonic mechanisms to adult therapeutic intervention. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.; Chien, C.; Bell, R.; Laudier, D.; Tufa, S.F.; Keene, D.R.; Andarawis-Puri, N.; Huang, A.H. Novel model of tendon regeneration reveals distinct cell mechanisms underlying regenerative and fibrotic tendon healing. Sci. Rep. 2017, 7, 45238. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, D.; Spanoudes, K.; Holladay, C.; Pandit, A.; Zeugolis, D. Progress in cell-based therapies for tendon repair. Adv. Drug Deliv. Rev. 2015, 84, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.I.; Costa-Almeida, R.; Gershovich, P.; Rodrigues, M.T.; Reis, R.L.; Gomes, M.E. Cell based approaches for tendon regeneration. In Tendon Regeneration: Understanding Tissue Physiology and Development to Engineer Functional Substitutes; Gomes, M.E., Rodrigues, M.T., Reis, R.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Kaux, J.-F.; Crielaard, J.-M. Platelet-rich plasma application in the management of chronic tendinopathies. Acta Orthop. Belg. 2013, 79, 10–15. [Google Scholar] [PubMed]
- Filardo, G.; Di Matteo, B.; Kon, E.; Merli, G.; Marcacci, M. Platelet-rich plasma in tendon-related disorders: Results and indications. Knee Surg. Sports Traumatol. Arthrosc. 2018, 26, 1984–1999. [Google Scholar] [CrossRef]
- Tang, J.B.; Zhou, Y.L.; Wu, Y.F.; Liu, P.Y.; Wang, X.T. Gene therapy strategies to improve strength and quality of flexor tendon healing. Expert Opin. Biol. Ther. 2016, 16, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Freedman, B.R.; Mooney, D.J. Biomaterials to Mimic and Heal Connective Tissues. Adv. Mater. 2019, 31, 1806695. [Google Scholar] [CrossRef] [PubMed]
- Domingues, R.M.A.; Gonçalves, A.I.; Costa-Almeida, R.; Rodrigues, M.T.; Reis, R.L.; Gomes, M.E. Fabrication of hierarchical and biomimetic fibrous structures to support the regeneration of tendon tissues. In Tendon Regeneration: Understanding Tissue Physiology and Development to Engineer Functional Substitutes; Gomes, M.E., Rodrigues, M.T., Reis, R.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 259–280. [Google Scholar]
- Loebel, C.; Burdick, J.A. Engineering stem and stromal cell therapies for musculoskeletal tissue repair. Cell Stem Cell 2018, 22, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Screen, H.R.C.; Berk, D.E.; Kadler, K.E.; Ramirez, F.; Young, M.F. Tendon functional extracellular matrix. J. Orthop. Res. 2015, 33, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Costa-Almeida, R.; Gonçalves, A.I.; Gershovich, P.; Rodrigues, M.T.; Reis, R.L.; Gomes, M.E. Tendon stem cell niche. In Tissue Engineering and Stem Cell Niche; Vol. Stem Cell Niche; Turksen, K., Ed.; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Murchison, N.D.; Price, B.A.; Conner, D.A.; Keene, D.R.; Olson, E.N.; Tabin, C.J.; Schweitzer, R. Regulation of tendon differentiation by scleraxis distinguishes force-transmitting tendons from muscle-anchoring tendons. Development 2007, 134, 2697–2708. [Google Scholar] [CrossRef] [Green Version]
- Shukunami, C.; Takimoto, A.; Oro, M.; Hiraki, Y. Scleraxis positively regulates the expression of tenomodulin, a differentiation marker of tenocytes. Dev. Biol. 2006, 298, 234–247. [Google Scholar] [CrossRef] [Green Version]
- Bi, Y.; Ehirchiou, D.; Kilts, T.M.; Inkson, C.A.; Embree, M.C.; Sonoyama, W.; Li, L.; Leet, A.I.; Seo, B.M.; Zhang, L.; et al. Identification of tendon stem/progenitor cells and the role of the extracellular matrix in their niche. Nat. Med. 2007, 13, 1219–1227. [Google Scholar] [CrossRef]
- Spanoudes, K.; Gaspar, D.; Pandit, A.; Zeugolis, D.I. The biophysical, biochemical, and biological toolbox for tenogenic phenotype maintenance in vitro. Trends Biotechnol. 2014, 32, 474–482. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, Y.; Liu, C. The horizon of Materiobiology: A perspective on material-guided cell behaviors and tissue engineering. Chem. Rev. 2017, 117, 4376–4421. [Google Scholar] [CrossRef]
- Docheva, D.; Müller, S.A.; Majewski, M.; Evans, C.H. Biologics for tendon repair. Adv. Drug Deliv. Rev. 2015, 84, 222–239. [Google Scholar] [CrossRef]
- Evans, C.H. Cytokines and the role they play in the healing of ligaments and tendons. Sports Med. 1999, 28, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Bedi, A.; Maak, T.; Walsh, C.; Rodeo, S.A.; Grande, D.; Dines, D.M.; Dines, J.S. Cytokines in rotator cuff degeneration and repair. J. Shoulder Elb. Surg. 2012, 21, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Millar, N.L.; Murrell, G.A.C.; McInnes, I.B. Inflammatory mechanisms in tendinopathy—Towards translation. Nat. Rev. Rheumatol. 2017, 13, 110–122. [Google Scholar] [CrossRef] [PubMed]
- GBD 2016 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef]
- Briggs, A.M.; Woolf, A.D.; Dreinhöfer, K.; Homb, N.; Hoy, D.G.; Kopansky-Giles, D.; Åkesson, K.; March, L. Reducing the global burden of musculoskeletal conditions. Bull. World Health Organ. 2018, 96, 366–368. [Google Scholar] [CrossRef]
- Lomas, A.J.; Ryan, C.N.M.; Sorushanova, A.; Shologu, N.; Sideri, A.I.; Tsioli, V.; Fthenakis, G.C.; Tzora, A.; Skoufos, I.; Quinlan, L.R.; et al. The past, present and future in scaffold-based tendon treatments. Adv. Drug Deliv. Rev. 2015, 84, 257–277. [Google Scholar] [CrossRef]
- Abbah, S.A.; Spanoudes, K.; O’Brien, T.; Pandit, A.; Zeugolis, D.I. Assessment of stem cell carriers for tendon tissue engineering in pre-clinical models. Stem Cell Res. Ther. 2014, 5, 38. [Google Scholar] [CrossRef]
- Costa-Almeida, R.; Reis, R.L.; Gomes, M.E. Metabolic Disease Epidemics: Emerging Challenges in Regenerative Medicine. Trends Endocrinol. Metab. 2019, 30, 147–149. [Google Scholar] [CrossRef]
- Schneider, M.; Angele, P.; Järvinen, T.A.H.; Docheva, D. Rescue plan for Achilles: Therapeutics steering the fate and functions of stem cells in tendon wound healing. Adv. Drug Deliv. Rev. 2017. [Google Scholar] [CrossRef]
- Chen, J.; Xu, J.; Wang, A.; Zheng, M. Scaffolds for tendon and ligament repair: Review of the efficacy of commercial products. Expert Rev. Med. Devices 2009, 6, 61–73. [Google Scholar] [CrossRef]
- Snedeker, J.G.; Foolen, J. Tendon injury and repair—A perspective on the basic mechanisms of tendon disease and future clinical therapy. Acta Biomater. 2017, 63, 18–36. [Google Scholar] [CrossRef]
- Longo, U.G.; Lamberti, A.; Maffulli, N.; Denaro, V. Tendon augmentation grafts: A systematic review. Br. Med. Bull. 2010, 94, 165–188. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Maffulli, N. Biology of tendon injury: Healing, modeling and remodeling. J. Musculoskelet. Neuronal Interact. 2006, 6, 181–190. [Google Scholar] [PubMed]
- Galatz, L.M.; Gerstenfeld, L.; Heber-Katz, E.; Rodeo, S.A. Tendon regeneration and scar formation: The concept of scarless healing. J. Orthop. Res. 2015, 33, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Meirelles, L.d.S.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. Cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal stem cells: Time to change the name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef]
- Lui, P.P.Y. Stem cell technology for tendon regeneration: Current status, challenges, and future research directions. Stem Cells Cloning Adv. Appl. 2015, 8, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Hindieh, J.; Leong, D.J.; Sun, H.B. Advances of stem cell based-therapeutic approaches for tendon repair. J. Orthop. Transl. 2017, 9, 69–75. [Google Scholar] [CrossRef]
- Deng, D.; Wang, W.; Wang, B.; Zhang, P.; Zhou, G.; Zhang, W.J.; Cao, Y.; Liu, W. Repair of achilles tendon defect with autologous ascs engineered tendon in a rabbit model. Biomaterials 2014, 35, 8801–8809. [Google Scholar] [CrossRef] [PubMed]
- Costa-Almeida, R.; Soares, R.; Granja, P.L. Fibroblasts as maestros orchestrating tissue regeneration. J. Tissue Eng. Regen. Med. 2018, 12, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Dehm, P.; Prockop, D.J. Synthesis and extrusion of collagen by freshly isolated cells from chick embryo tendon. Biochim. Biophys. Acta (BBA) Nucleic Acids Protein Synth. 1971, 240, 358–369. [Google Scholar] [CrossRef]
- Banes, A.J.; Donlon, K.; Link, G.W.; Gillespie, Y.; Bevin, A.G.; Peterson, H.D.; Bynum, D.; Watts, S.; Dahners, L. Cell populations of tendon: A simplified method for isolation of synovial cells and internal fibroblasts: Confirmation of origin and biologic properties. J. Orthop. Res. 1988, 6, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Lui, P.P.Y. Markers for the identification of tendon-derived stem cells in vitro and tendon stem cells in situ—Update and future development. Stem Cell Res. Ther. 2015, 6, 106. [Google Scholar] [CrossRef] [PubMed]
- Birch, H.L.; Sinclair, C.; Goodship, A.E.; Smith, R.K. Tendon and ligament physiology. In Equine Sports Medicine and Surgery, 2nd ed.; Hinchcliff, K.W., Kaneps, A.J., Geor, R.J., Eds.; Saunders Elsevier: Amsterdam, The Netherlands, 2014; pp. 167–188. [Google Scholar]
- Bernard-Beaubois, K.; Hecquet, C.; Houcine, O.; Hayem, G.; Adolphe, M. Culture and characterization of juvenile rabbit tenocytes. Cell Biol. Toxicol. 1997, 13, 103–113. [Google Scholar] [CrossRef]
- Kryger, G.S.; Chong, A.K.; Costa, M.; Pham, H.; Bates, S.J.; Chang, J. A comparison of tenocytes and mesenchymal stem cells for use in flexor tendon tissue engineering. J. Hand Surg. Am. 2007, 32, 597–605. [Google Scholar] [CrossRef]
- Luo, Q.; Song, G.; Song, Y.; Xu, B.; Qin, J.; Shi, Y. Indirect co-culture with tenocytes promotes proliferation and mrna expression of tendon/ligament related genes in rat bone marrow mesenchymal stem cells. Cytotechnology 2009, 61, 1–10. [Google Scholar] [CrossRef]
- Wagget, A.D.; Benjamin, M.; Ralphs, J.R. Connexin 32 and 43 gap junctions differentially modulate tenocyte response to cyclic mechanical load. Eur. J. Cell Biol. 2006, 85, 1145–1154. [Google Scholar] [CrossRef]
- Rui, Y.F.; Lui, P.P.Y.; Li, G.; Fu, S.C.; Lee, Y.W.; Chan, K.M. Isolation and characterization of multipotent rat tendon-derived stem cells. Tissue Eng. Part A 2010, 16, 1549–1558. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.H.-C. Characterization of differential properties of rabbit tendon stem cells and tenocytes. BMC Musculoskelet. Disord. 2010, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Lui, P.P.; Rui, Y.F.; Wong, Y.M. Comparison of potentials of stem cells isolated from tendon and bone marrow for musculoskeletal tissue engineering. Tissue Eng. Part A 2012, 18, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Lui, P.; Wong, Y.; Tan, Q.; Chan, K. Altered fate of tendon-derived stem cells isolated from a failed tendon-healing animal model of tendinopathy. Stem Cells Dev. 2013, 22, 1076–1085. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, I.; Wang, J.H.; Iwasaki, K.; Shimizu, T.; Okano, T. The effect of tendon stem/progenitor cell (TSC) sheet on the early tendon healing in a rat Achilles tendon injury model. Acta Biomater. 2016, 42, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, Q.; Wang, K.; Liu, H.; Ma, C.; Huang, H.; Liu, Y. Isolation and biological characterization of tendon-derived stem cells from fetal bovine. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 846–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Chan, K.M.; Zhang, J.F.; Li, G. Tendon-derived stem cells undergo spontaneous tenogenic differentiation. Exp. Cell Res. 2016, 341, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Clegg, P.D.; Comerford, E.J.; Canty-Laird, E.G. A comparison of the stem cell characteristics of murine tenocytes and tendon-derived stem cells. BMC Musculoskelet. Disord. 2018, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- de Mos, M.; Koevoet, W.J.L.M.; Jahr, H.; Verstegen, M.M.A.; Heijboer, M.P.; Kops, N.; van Leeuwen, J.P.T.M.; Weinans, H.; Verhaar, J.A.N.; van Osch, G.J.V.M. Intrinsic differentiation potential of adolescent human tendon tissue: An in-vitro cell differentiation study. BMC Musculoskelet. Disord. 2007, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Randelli, P.; Conforti, E.; Piccoli, M.; Ragone, V.; Creo, P.; Cirillo, F.; Masuzzo, P.; Tringali, C.; Cabitza, P.; Tettamanti, G.; et al. Isolation and characterization of 2 new human rotator cuff and long head of biceps tendon cells possessing stem cell-like self-renewal and multipotential differentiation capacity. Am. J. Sports Med. 2013, 41, 1653–1664. [Google Scholar] [CrossRef]
- Yin, Z.; Chen, X.; Chen, J.L.; Shen, W.L.; Hieu Nguyen, T.M.; Gao, L.; Ouyang, H.W. The regulation of tendon stem cell differentiation by the alignment of nanofibers. Biomaterials 2010, 31, 2163–2175. [Google Scholar] [CrossRef]
- Ruzzini, L.; Abbruzzese, F.; Rainer, A.; Longo, U.G.; Trombetta, M.; Maffulli, N.; Denaro, V. Characterization of age-related changes of tendon stem cells from adult human tendons. Knee Surg. Sports Traumatol. Arthrosc. 2013, 22, 2856–2866. [Google Scholar] [CrossRef] [PubMed]
- Nagura, I.; Kokubu, T.; Mifune, Y.; Inui, A.; Takase, F.; Ueda, Y.; Kataoka, T.; Kurosaka, M. Characterization of progenitor cells derived from torn human rotator cuff tendons by gene expression patterns of chondrogenesis, osteogenesis, and adipogenesis. J. Orthop. Surg. Res. 2016, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Vigano, M.; Perucca Orfei, C.; Colombini, A.; Stanco, D.; Randelli, P.; Sansone, V.; de Girolamo, L. Different culture conditions affect the growth of human tendon stem/progenitor cells (TSPCs) within a mixed tendon cells (TCs) population. J. Exp. Orthop. 2017, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.I.; Rodrigues, M.T.; Lee, S.J.; Atala, A.; Yoo, J.J.; Reis, R.L.; Gomes, M.E. Understanding the role of growth factors in modulating stem cell tenogenesis. PLoS ONE 2013, 8, e83734. [Google Scholar] [CrossRef] [PubMed]
- Orfei, C.P.; Viganò, M.; Pearson, J.R.; Colombini, A.; Luca, P.D.; Ragni, E.; Santos-Ruiz, L.; de Girolamo, L. In vitro Induction of Tendon-Specific Markers in Tendon Cells, Adipose- and Bone Marrow-Derived Stem Cells is Dependent on TGF3, BMP-12 and Ascorbic Acid Stimulation. Int. J. Mol. Sci. 2019, 20, 149. [Google Scholar] [CrossRef] [PubMed]
- United States Bone and Joint Initiative: The Burden of Musculoskeletal Diseases in the United States (Bmus), 3rd ed. 2014. Available online: http://www.Boneandjointburden.Org (accessed on 10 June 2019).
- Rinoldi, C.; Fallahi, A.; Yazdi, I.K.; Paras, J.C.; Kijeńska-Gawrońska, E.; Santiago, G.T.-D.; Tuoheti, A.; Demarchi, D.; Annabi, N.; Khademhosseini, A.; et al. Mechanical and biochemical stimulation of 3d multilayered scaffolds for tendon tissue engineering. ACS Biomater. Sci. Eng. 2019. [Google Scholar] [CrossRef]
- Uysal, C.A.; Tobita, M.; Hyakusoku, H.; Mizuno, H. Adipose-derived stem cells enhance primary tendon repair: Biomechanical and immunohistochemical evaluation. J. Plast. Reconstr. Aesthet. Surg. 2012, 65, 1712–1719. [Google Scholar] [CrossRef] [PubMed]
- Lovati, A.; Corradetti, B.; Consiglio, A.L.; Recordati, C.; Bonacina, E.; Bizzaro, D.; Cremonesi, F. Characterization and differentiation of equine tendon-derived progenitor cells. J. Biol. Regul. Homeost. Agents 2011, 25, S75–S84. [Google Scholar] [PubMed]
- Al-ani, M.K.; Xu, K.; Sun, Y.; Pan, L.; Xu, Z.; Yang, L. Study of bone marrow mesenchymal and tendon-derived stem cells transplantation on the regenerating effect of achilles tendon ruptures in rats. Stem Cells Int. 2015, 2015, 984146. [Google Scholar] [CrossRef]
- Vermeulen, S.; Vasilevich, A.; Tsiapalis, D.; Roumans, N.; Vroemen, P.; Beijer, N.R.M.; Eren, A.D.; Zeugolis, D.; Boer, J.D. Identification of topographical architectures supporting the phenotype of rat tenocytes. Acta Biomater. 2019, 83, 277–290. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, E.; Yang, L.; Tu, W.; Lin, J.; Yuan, C.; Bunpetch, V.; Chen, X.; Ouyang, H. Histone deacetylase inhibitor treated cell sheet from mouse tendon stem/progenitor cells promotes tendon repair. Biomaterials 2018, 172, 66–82. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, F.; Salamanna, F.; Tschon, M.; Maglio, M.; Aldini, N.N.; Fini, M. Mesenchymal stem cells for tendon healing: What is on the horizon? J. Tissue Eng. Regen. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ekwueme, E.C.; Shah, J.V.; Mohiuddin, M.; Ghebes, C.A.; Crispim, J.F.; Saris, D.L.B.F.; Fernandes, H.A.M.; Freeman, J.W. Cross-talk between human tenocytes and bone marrow stromal cells potentiates extracellular matrix remodeling in vitro. J. Cell. Biochem. 2016, 117, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Awad, H.A.; Butler, D.L.; Boivin, G.P.; Smith, F.N.L.; Malaviya, P.; Huibregtse, B.; Caplan, A.I. Autologous mesenchymal stem cell-mediated repair of tendon. Tissue Eng. 1999, 5, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Liu, Y.; Wang, B.; Sun, Y.; Xu, J.; Lee, W.Y.W.; Xu, L.; Zhang, J.; Li, G. The use of co-cultured mesenchymal stem cells with tendon-derived stem cells as a better cell source for tendon repair. Tissue Engineer. Part A 2016, 19. [Google Scholar]
- Kapacee, Z.; Yeung, C.-Y.C.; Lu, Y.; Crabtree, D.; Holmes, D.F.; Kadler, K.E. Synthesis of embryonic tendon-like tissue by human marrow stromal/mesenchymal stem cells requires a three-dimensional environment and transforming growth factor β3. Matrix Biol. 2010, 29, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Sevivas, N.; Teixeira, F.G.; Portugal, R.; Direito-Santos, B.; Espregueira-Mendes, J.; Oliveira, F.J.; Silva, R.F.; Sousa, N.; Sow, W.T.; Luong, N.; et al. Mesenchymal stem cell secretome improves tendon cell viability in vitro and tendon-bone healing in vivo when a tissue engineering strategy is used in a rat model of chronic massive rotator cuff tear. Am. J. Sports Med. 2018, 46, 449–459. [Google Scholar] [CrossRef]
- Baberg, F.; Geyh, S.; Waldera-Lupa, D.; Stefanski, A.; Zilkens, C.; Haas, R.; Schroeder, T.; Stühler, K. Secretome analysis of human bone marrow derived mesenchymal stromal cells. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 434–441. [Google Scholar] [CrossRef]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived Stem Cells: Isolation, Expansion and Differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef]
- De Francesco, F.; Ricci, G.; D’Andrea, F.; Nicoletti, G.; Ferraro, G. Human Adipose Stem Cells: From Bench to Bedside. Tissue Eng. Part B Rev. 2015, 21, 572–584. [Google Scholar] [CrossRef] [Green Version]
- de Mattos Carvalho, A.; Alves, A.L.G.; Gomes de Oliveira, P.G.; Álvarez, L.E.C.; Amorim, R.L.; Hussni, C.A.; Deffune, E. Use of adipose tissue-derived mesenchymal stem cells for experimental tendinitis therapy in equines. J. Equine Vet. Sci. 2011, 31, 26–34. [Google Scholar] [CrossRef]
- Veronesi, F.; Torricelli, P.; Bella, E.D.; Pagani, S.; Fini, M. In vitro mutual interaction between tenocytes and adipose-derived mesenchymal stromal cells. Cytotherapy 2015, 17, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Costa-Almeida, R.; Calejo, I.; Reis, R.; Gomes, M. Crosstalk between adipose stem cells and tendon cells reveals a temporal regulation of tenogenesis by extracellular matrix deposition and remodeling. J. Cell. Physiol. 2017. [Google Scholar] [CrossRef]
- Costa-Almeida, R.; Berdecka, D.; Rodrigues, M.; Reis, R.; Gomes, M. Tendon explant cultures to study the communication between adipose stem cells and native tendon niche. J. Cell. Biochem. 2017. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Isolation Method | Tissue Origin | Cell Type | Method Description | Culture Medium | Characterization | Differentiation Potential | Ref. | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Morphology | Gene Expression | Protein Expression | Flow Cytometry | ||||||||
| Animal Origin | |||||||||||
| Digestion | Mouse Patellar tendons | TSPCs | Removal of tendon sheath and surrounding paratenon; cut into small pieces; digested with 3 mg/mL collagenase type I + 4 mg/mL dispase/PBS (1 h, 37 °C) | α-MEM + 20% lot-selected FBS + 100 mM 2-mercaptoethanol | Heterogeneous colonies | Scx, cartilage oligomeric protein (Comp), Sox9 and Runx2 | Expression of collagen type I, fibronectin, COMP, tenascin-c Low expression of α-SMA | Positive Sca-1 CD90.2 CD44 | Negative CD34 CD117 CD45 Flk-1 CD18 CD144 | Osteogenic Adipogenic | [20] |
| Digestion | Rat Flexor Tendons | TDSCs | Removal of peritendinous connective tissue; tissue minced and digested with 3 mg/mL collagenase type I (2.5 h, 37 °C); strained through a 70 µm cell strainer; suspension washed in PBS (centrifuge 300× g, 5 min); cells resuspended in medium | DMEM + 10% FBS + 100 U/mL penicillin + 100 µg/mL streptomycin + 2 mM L-glutamine | P0: large polygonal and star-shaped cells; P1: flat and slender cells; P3: fibroblast-like cells; Heterogeneous colonies | Expression of α-SMA, tenascin-c, tenomodulin and aggrecan | Positive CD90 CD44 | Negative CD34 CD31 | Osteogenic Chondrogenic Adipogenic | [54] | |
| Digestion | Rabbit Patellar and Achilles Tendons | TSCs and tenocytes | Tendon portions (1 mm3) minced; 100 mg of tissue sample digested with 3 mg collagenase type I + 4 mg dispase/1 mL of PBS (1 h, 37 °C); suspensions were centrifuged (1500× g, 15 min); cell pellet resuspended; tenocytes were obtained by application of local trypsin to colonies and culture in T25 flasks | DMEM + 20% FBS + 100 U/mL penicillin + 100 µg/mL streptomycin Tenocytes: DMEM + 10% FBS | TSCs: Cobblestone shape; unequal colonies formation Tenocytes: elongated shape | TSCs differentiated: Expression of Runx2, Sox2 and Col2A1 | Expression of Oct-4, SSEA-4 and nucleostemin in TSCs Tenocytes exhibited an absence for the same markers | TSCs: Osteogenic Chondrogenic Adipogenic | [55] | ||
| Digestion | Rat Patellar Tendons | TDSCs | Removal of peritendinous connective tissue; tissue was minced; digested with 3 mg/mL of collagenase type I; strained through a 70 µm cell strainer; cell suspension resuspended in culture medium | LG-DMEM + 10% FBS + 100 U/mL penicillin + 100 µg/mL streptomycin + 2 mM L-glutamine | Colonies formation | Expression of Scx, Col1A1, Dcn, Alp, Col2A1, Col3A1, Tnc, Aca, Tnmd, Oct4, Sox2, Nanog | Positive CD90 CD73 | Negative CD45 | Osteogenic Chondrogenic Adipogenic | [56] | |
| Digestion | Rat Patellar Tendons (Diseased and healthy) | TDSCs | Removal of peritendinous connective tissue; tissue was minced; digested with 3 mg/mL of collagenase type I; strained through a 70 µm cell strainer; cell suspension resuspended in culture medium | LG-DMEM + 10% FBS + 100 U/mL penicillin + 100 µg/mL streptomycin + 2 mM L-glutamine | Colonies formation | Higher expression of Col1A1, Scx and Tnmd in healthy TDSCs | Expression of SOX9 | Positive CD90 CD44* CD73* *Lower levels in diseased TDSCs | Negative CD31 CD34 CD45 CD11b | Osteogenic Chondrogenic Adipogenic | [57] |
| Digestion | Rat Achilles tendons | TSCs/TSCs Sheets | Removal of tendon sheath and paratenon; tissue minced into small pieces; about 100 mg of tissue digested with 3 mg/mL of collagenase type I and 4 mg/mL of dispase in PBS (2.5 h); centrifuged at 1500× g, for 15 min; cell suspension strained through a 70 µm cell strainer; cell pellet resuspended in medium in T25 flasks | DMEM + 10% FBS + 100 U/mL penicillin + 100 µg/mL streptomycin | Heterogeneous colonies; P3: homogeneous and cobblestone-shape cells | Expression of collagen type I, collagen type II and tenomodulin | Positive CD90 CD44 | Negative CD31 CD34 CD45 | Osteogenic Adipogenic | [58] | |
| Digestion | Fetal Bovine Achilles Tendons | TDSCs | Removal of peritendinous connective tissue; washed with PBS; tissue trimmed into 1 mm3 pieces; digested with 0.1% collagenase type I (1 h, 37 °C); sample strained through a 70 µm cell strainer and added complete medium to stop reaction; cell suspension centrifuged (1200 rpm, 10 min); cells resuspended in culture medium | LG-DMEM + 15% FBS + 100 U/mL penicillin + 100 µg/mL streptomycin + 2.5 ng/mL bFGF and 2 mM L-glutamine | Spindle-shaped or fusoid cells; Colonies formation | Expression of Col1A1, Tnc, Col3A1 and Cd44 | Expression of Collagen type I and III, CD44 and Tenascin-C | Osteogenic Adipogenic Chondrogenic | [59] | ||
| Digestion | Rat Achilles Tendons | TDSCs | Removal of tendon sheath and paratenon; tissue cut into small pieces; digested with 2 mg/mL collagenase type I (2.5 h, 37 °C); resuspend in culture medium; | DMEM + 10% FBS + 50 µg/mL penicillin + 50 µg/mL streptomycin +100 µg/mL neomycin | Up-regulation of CD90, nucleostemin (NS), Col3A1, lysyl oxidase (Lox), Tnc; Low expression of Tnmd, Dcn and Fmod | Tenogenic | [60] | ||||
| Digestion | Murine | TDSCs and tenocytes | Tissue digested for 3 h at 37 °C in 20 mL 375 U/mL collagenase type I and 0.05% trypsin; cell suspension strained and centrifuged at 1200× g for 10 min; cells resuspended in medium | DMEM + 20% FCS +100 U/mL penicillin + 100 μg/mL streptomycin and 2 μg/mL amphotericin B | TDCs: smaller and round shaped; Tenocytes: large and flat fibroblast-like cells | TDCs: Nanog, CD73, CD45, Scx and Mkx; Tenocytes: Tnc, thrombospondin-4 and Tnmd | Osteogenic Adipogenic | [61] | |||
| Human Origin | |||||||||||
| Digestion | Hamstring tendons | TSPCs | Removal of tendon sheath and surrounding paratenon; cut into small pieces; digested with 3 mg/mL collagenase type I + 4 mg/mL dispase/PBS (1 h, 37 °C) | α-MEM + 20% lot-selected FBS + 100 mM 2-mercaptoethanol | Expression of Tnmd, Comp, Runx2 | Positive Stro-1 CD146 CD90 CD44 | Negative CD18 CD34 CD45 CD117 CD106 | Osteogenic Adipogenic Chondrogenic | [20] | ||
| Explant | Hamstring Tendons | TDCs | Removal of peritendineum; cut into 3 mm3 pieces and placed in culture medium | DMEM + 10% FCS + 50 µg/mL gentamicin + 1.5 µg/mL fungizone | Spindle-shape | Positive CD105 | Negative CD34 | Osteogenic Adipogenic Chondrogenic | [62] | ||
| Digestion | Supraspinatus and long head of biceps tendons | TDSCs | Washed with PBS; tissue was cut into small pieces and digested with 3 mg/mL collagenase type I and 4 mg/mL dispase, in PBS (1.5 h, 37 °C); sample was centrifuged; cell pellet resuspended in culture medium | α-MEM + 20% lot-selected FBS + 2 mM L-glutamine + 100 U/mL penicillin + 100 µg/mL streptomycin | Fibroblast-like shape; Colonies formation | Expression of Col1A1, Col3A1, Mmp-2, TGFB1, Cx43 | Positive CD90 CD44 CD146 | Negative CD2 CD3 CD11b CD14 CD15 CD16 CD19 CD56 CD123 CD235a CD18 CD34 CD117 | Osteogenic Adipogenic Myogenic | [63] | |
| Digestion | Fetal Achilles Tendon | TSPC | Tissues were cut into 1–2 mm3 pieces and washed 3x with PBS; digested with 0.25% collagenase (37 °C, overnight); cell suspension was cultured in culture medium; | DMEM low glucose + 10% FBS + 1% penicillin-streptomycin | Fibroblast-like morphology | Positive CD105 CD90 CD146 | Negative CD44 CD18 CD34 | Osteogenic Chondrogenic Adipogenic | [64] | ||
| Digestion | Hamstring Tendons | TSC | Tissue was harvested in 8 mm3 blocks; surrounding adipose and muscle tissues were cleaned off; samples were cut into small pieces and digested with 3 mg/mL collagenase type I + 4 mg/mL dispase in PBS (1 h, 37 °C); cell suspension was centrifuged (2000 rpm, 15 min); cell pellet was resuspended in culture medium | α-MEM + 20% FBS + 1% penicillin and streptomycin + 100 mM 2-mercaptoethanol | Elongated fibroblastic-like cells and polygonal-shaped cells | Expression of CD146, STRO1, α-SMA and Tenomodulin | Positive CD44 | Osteogenic Adipogenic Chondrogenic | [65] | ||
| Explants | Rotator cuff Tendons | TSPC | Tissue was minced into 1 mm3 pieces, placed on a 10-mm diameter culture dish and cultured in a monolayer with medium; minced tissue was removed after 1 week; after 2–3 weeks, the cells were harvested with 0.05% trypsin/EDTA and cultured in non-coated flasks | α-MEM + 10% heat-inactivated FBS + 2 mM L-glutamine + antibiotics | Fibroblast-like spindle shape; | Positive CD29 CD44 CD105 CD166 | Negative CD14 CD34 CD45 | Osteogenic Adipogenic Chondrogenic | [66] | ||
| Digestion | Gracilis and semitendinosus tendons | TSPCs within tendon resident cells | Tissue was fragmented and digested with 0.3 % (w/v) collagenase type I (16 h, 37 °C) in HG-DMEM; samples were strained through a 100 µm cell strainer and centrifuged (300× g, 5 min); cells were plated in culture medium; | HG-DMEM + 10% FBS 50 µg/mL penicillin + 50 µg/mL streptomycin +2 mM L-glutamine Cultured in the presence/absence of 5 ng/mL bFGF | Fibroblast-like shape; Low colony forming capability | Expression of Scx, Tnc, Mkx, Oct4 (in presence and absence of bFGF) | Positive CD13 CD73 CD90 CD54 | Negative CD34 CD45 | Osteogenic Chondrogenic | [67] | |
| Cells | Condition | Strategy | Reference |
|---|---|---|---|
| BM-MSCs | Rotator cuff tears | Arthroscopic repair combined with bone marrow aspirate | NCT03688308 |
| Non-retracted supraspinatus tendon tear | Regenexx-SD injection: Bone marrow aspirate injection into the area of the damaged tendon | NCT01788683 | |
| Full thickness rotator cuff tears | Arthroscopic repair combined with bone marrow aspirate | NCT02484950 | |
| ASCs | Intractable common extensor tendinosis | Intra-tendon injection of allogeneic ASCs combined with fibrin glue | NCT03449082 |
| Rotator cuff tear Lateral epicondylitis | Injection of autologous ASCs (1 million cells/10 kg body weight) once a week, three times | NCT03279796 | |
| Partial-thickness rotator cuff tear Rotator cuff tendinitis | Single injection of adipose-derived regenerative cells (ADRCs) | NCT03752827 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa-Almeida, R.; Calejo, I.; Gomes, M.E. Mesenchymal Stem Cells Empowering Tendon Regenerative Therapies. Int. J. Mol. Sci. 2019, 20, 3002. https://doi.org/10.3390/ijms20123002
Costa-Almeida R, Calejo I, Gomes ME. Mesenchymal Stem Cells Empowering Tendon Regenerative Therapies. International Journal of Molecular Sciences. 2019; 20(12):3002. https://doi.org/10.3390/ijms20123002
Chicago/Turabian StyleCosta-Almeida, Raquel, Isabel Calejo, and Manuela E. Gomes. 2019. "Mesenchymal Stem Cells Empowering Tendon Regenerative Therapies" International Journal of Molecular Sciences 20, no. 12: 3002. https://doi.org/10.3390/ijms20123002
APA StyleCosta-Almeida, R., Calejo, I., & Gomes, M. E. (2019). Mesenchymal Stem Cells Empowering Tendon Regenerative Therapies. International Journal of Molecular Sciences, 20(12), 3002. https://doi.org/10.3390/ijms20123002