Reactive Oxygen Species Are Key Mediators of Demyelination in Canine Distemper Leukoencephalitis but not in Theiler’s Murine Encephalomyelitis


Abstract
:1. Introduction
2. Results
2.1. Lesion Profile in CDV-DL
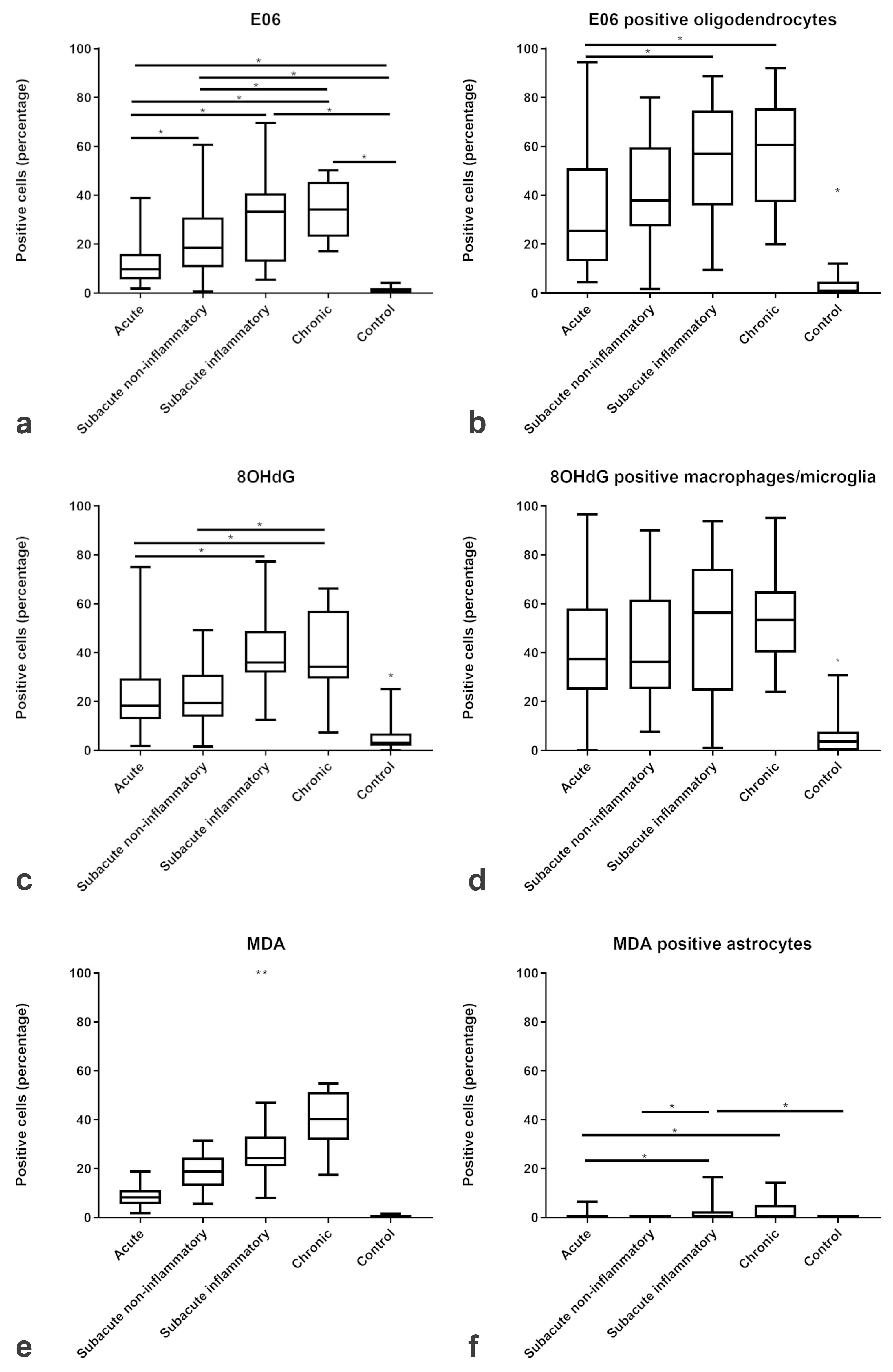
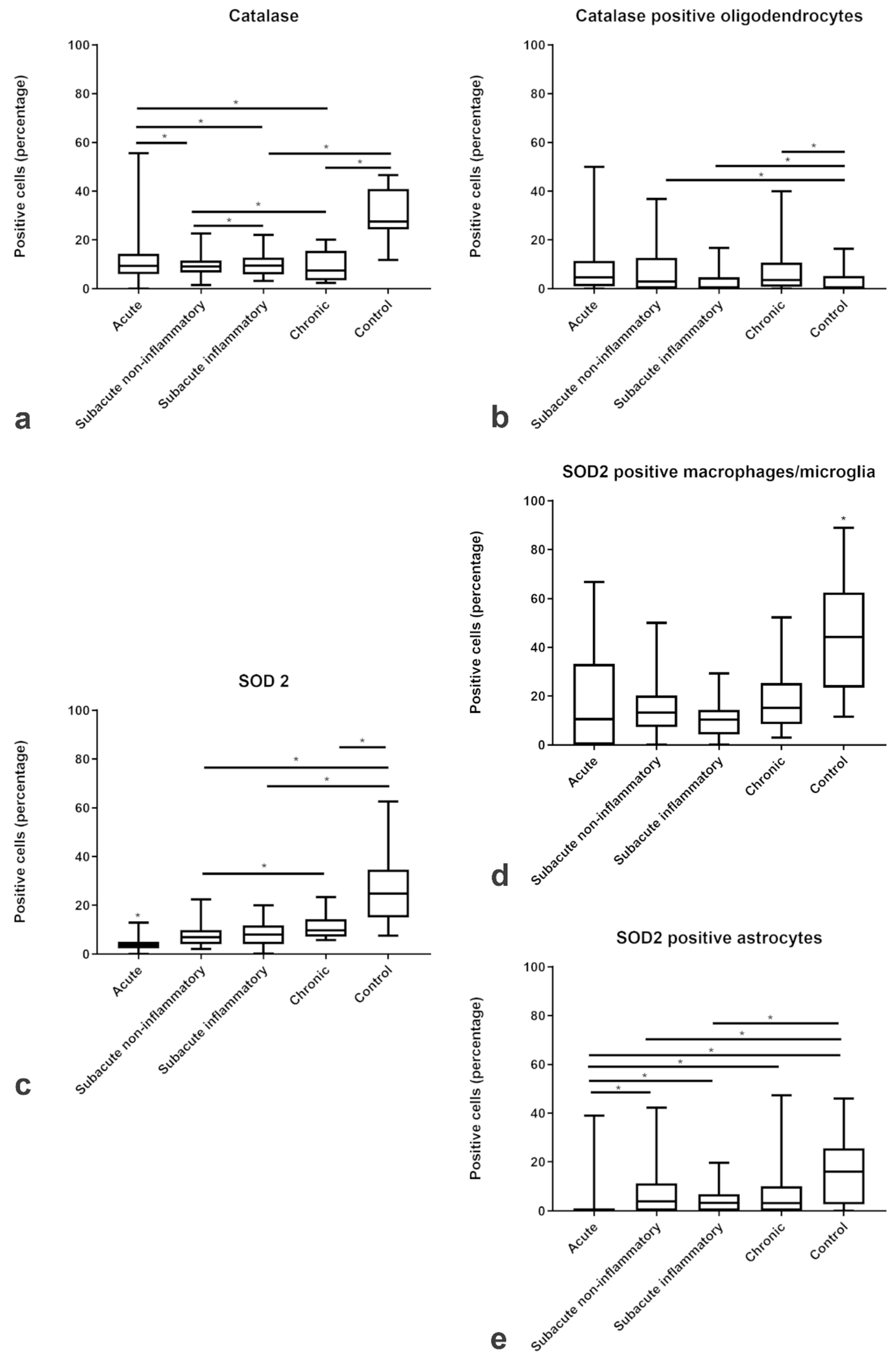
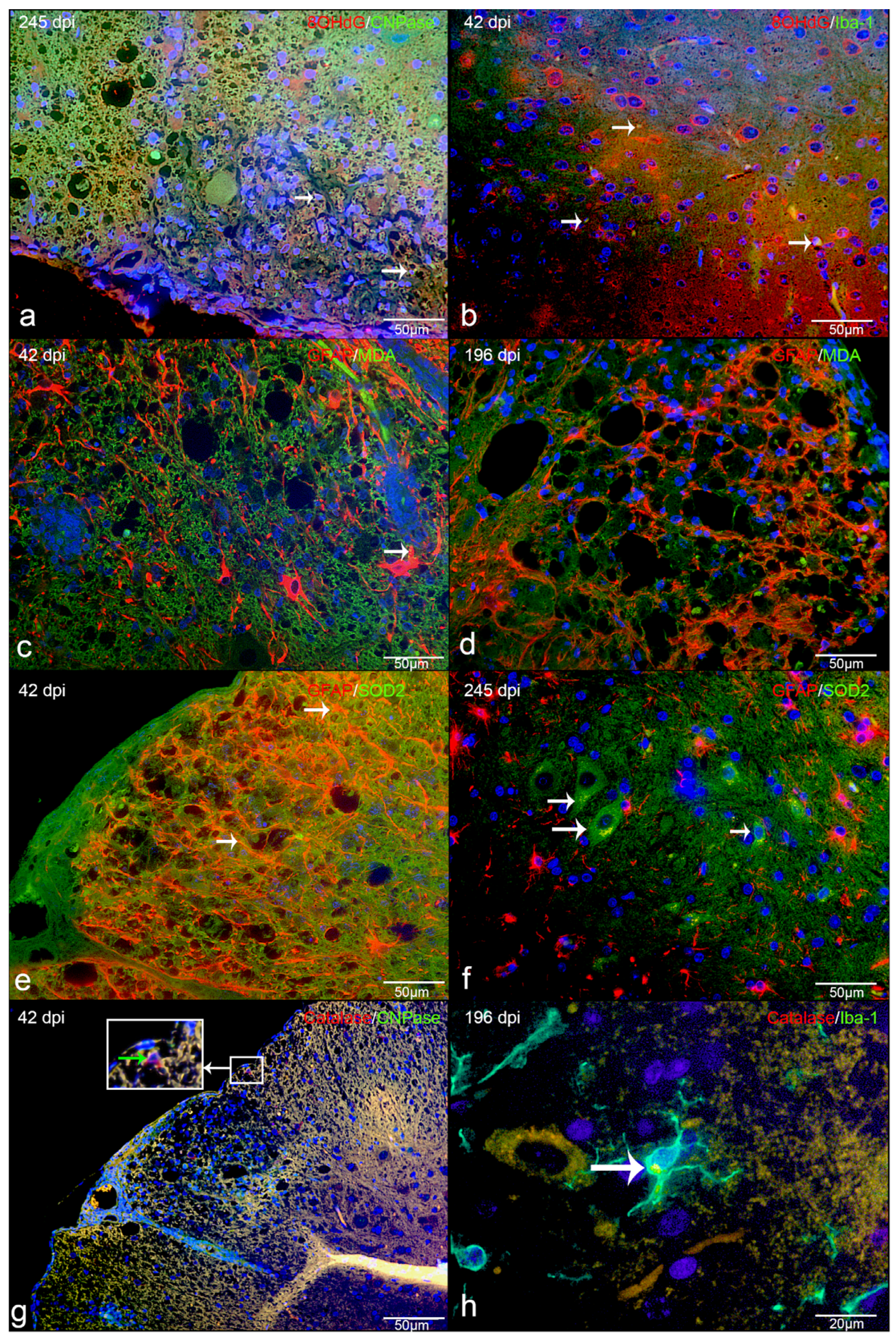
2.2. Amount and Localization ROS-Induced Metabolites in CDV-DL
2.3. Lesion Profile in TMEV-DL
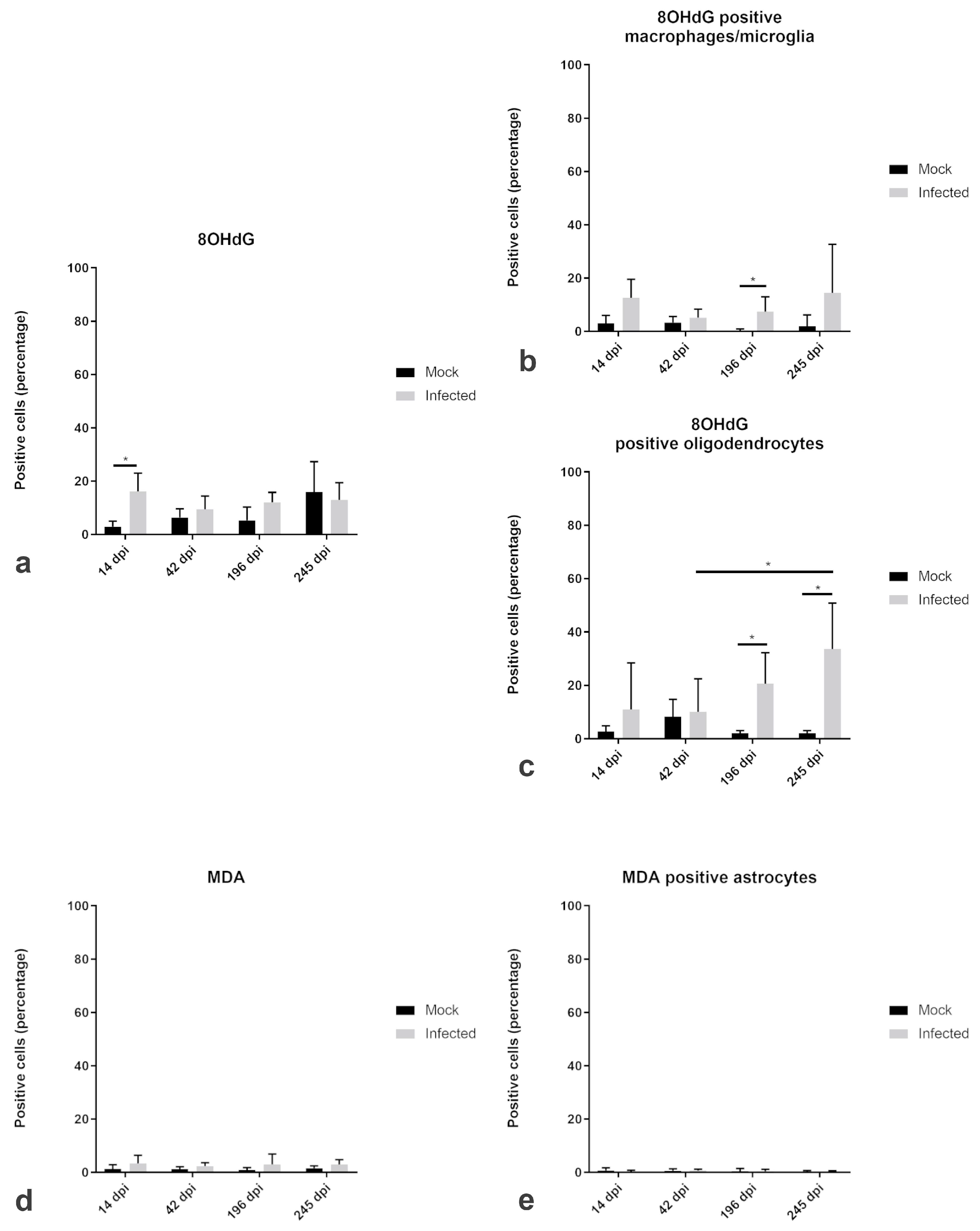
2.4. Amount and Localization ROS-Induced Metabolites and Antioxidant Enzymes in TMEV-DL
2.5. Transcriptional Activity of ROS-Generating and Detoxifying Pathways in CDV-DL
2.6. Transcriptional Activity of ROS-Generating and Detoxifying Pathways in TMEV-DL
2.7. Intersections Between Differentially Expressed Genes in CDV-DL and TMEV-DL
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Design
4.3. Histology and Immunofluorescence
4.4. Microarray Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CDV | Canine distemper virus |
| CDV-DL | Canine distemper virus -induced demyelinating leukoencephalitis |
| TME | Theiler’s murine encephalomyelitis |
| TMEV-DL | Theiler’s murine encephalomyelitis virus—induced demyelinating leukomyelitis |
| TMEV | Theiler’s murine encephalomyelitis virus |
| MS | Multiple Sclerosis |
| ROS | Reactive oxygen species |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| ATP | Adenosine triphosphate |
| MDA | Malondialdehyde |
| 8-OHdG | 8-hydroxydeoxyguanosine |
| 8-OHG | 8-hydroxyguanosine |
| CNS | Central nervous system |
| PUFAs | Polyunsaturated fatty acids |
| SOD | Superoxide dismutases |
| MnSOD, aka SOD2 | Manganese SOD |
| O2• − | superoxide |
| H2O2 | hydrogen peroxide |
| CSF | Cerebrospinal fluid |
| LFB | Luxol fast blue |
| dpi | Days post infection |
| CNPase | 2′,3′-cyclic-nucleotide 3′-phosphodiesterase |
| GFAP | Glial fibrillary acidic protein |
| Iba-1 | Ionized calcium-binding adapter molecule 1 |
| ND5 | NADH dehydrogenase subunit 5 |
| RAC2 | Ras-related C3 botulinum toxin substrate 2 |
| CYBB | Cytochrome b-245, beta polypeptide |
| XDH | Xanthine dehydrogenase |
| NOX2 | NADPH oxidase |
| EAE | experimental autoimmune encephalomyelitis |
| HE | Hematoxylin and eosin |
| LFB | Luxol fast blue |
| PBS | Phosphate buffered saline |
| FCE | Fibrocartilaginous embolus |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
References
- Witte, M.E.; Mahad, D.J.; Lassmann, H.; van Horssen, J. Mitochondrial dysfunction contributes to neurodegeneration in multiple sclerosis. Trends Mol. Med. 2014, 20, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [CrossRef] [Green Version]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.J.; Kapoor, R.; Felts, P.A. Demyelination: The role of reactive oxygen and nitrogen species. Brain Pathol. 1999, 9, 69–92. [Google Scholar] [CrossRef] [PubMed]
- Gilgun-Sherki, Y.; Melamed, E.; Offen, D. The role of oxidative stress in the pathogenesis of multiple sclerosis: The need for effective antioxidant therapy. J. Neurol. 2004, 251, 261–268. [Google Scholar] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Palinski, W.; Yla-Herttuala, S.; Rosenfeld, M.E.; Butler, S.W.; Socher, S.A.; Parthasarathy, S.; Curtiss, L.K.; Witztum, J.L. Antisera and monoclonal antibodies specific for epitopes generated during oxidative modification of low density lipoprotein. Arteriosclerosis 1990, 10, 325–335. [Google Scholar] [CrossRef]
- Palinski, W.; Horkko, S.; Miller, E.; Steinbrecher, U.P.; Powell, H.C.; Curtiss, L.K.; Witztum, J.L. Cloning of monoclonal autoantibodies to epitopes of oxidized lipoproteins from apolipoprotein E-deficient mice. Demonstration of epitopes of oxidized low density lipoprotein in human plasma. J. Clin. Investig. 1996, 98, 800–814. [Google Scholar] [CrossRef]
- Horkko, S.; Bird, D.A.; Miller, E.; Itabe, H.; Leitinger, N.; Subbanagounder, G.; Berliner, J.A.; Friedman, P.; Dennis, E.A.; Curtiss, L.K.; et al. Monoclonal autoantibodies specific for oxidized phospholipids or oxidized phospholipid-protein adducts inhibit macrophage uptake of oxidized low-density lipoproteins. J. Clin. Investig. 1999, 103, 117–128. [Google Scholar] [CrossRef]
- Hametner, S.; Wimmer, I.; Haider, L.; Pfeifenbring, S.; Bruck, W.; Lassmann, H. Iron and neurodegeneration in the multiple sclerosis brain. Ann. Neurol. 2013, 74, 848–861. [Google Scholar] [CrossRef]
- Haider, L.; Fischer, M.T.; Frischer, J.M.; Bauer, J.; Hoftberger, R.; Botond, G.; Esterbauer, H.; Binder, C.J.; Witztum, J.L.; Lassmann, H. Oxidative damage in multiple sclerosis lesions. Brain 2011, 134, 1914–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.L.; Chiou, C.C.; Chang, P.Y.; Wu, J.T. Urinary 8-OHdG: A marker of oxidative stress to DNA and a risk factor for cancer, atherosclerosis and diabetics. Clin. Chim Acta 2004, 339, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Finean, J.; Robertson, J. Lipids and the structure of myelin. Br. Med. Bull 1958, 14, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Yurlova, L.; Simons, M. Central nervous system myelin: Structure, synthesis and assembly. Trends Cell Biol. 2011, 21, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Chrast, R.; Saher, G.; Nave, K.-A.; Verheijen, M.H. Lipid metabolism in myelinating glial cells: Lessons from human inherited disorders and mouse models. J. Lipid Res. 2011, 52, 419–434. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.M.; Melov, S. SOD2 in mitochondrial dysfunction and neurodegeneration. Free Radic. Biol. Med. 2013, 62, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chance, B.; Greenstein, D.; Roughton, F. The mechanism of catalase action. I. Steady-state analysis. Arch. Biochem. Biophys 1952, 37, 301–321. [Google Scholar] [CrossRef]
- Baumgärtner, W.; Örvell, C.; Reinacher, M. Naturally occurring canine distemper virus encephalitis: Distribution and expression of viral polypeptides in nervous tissues. Acta Neuropathol. 1989, 78, 504–512. [Google Scholar] [CrossRef]
- Moreno, S.; Mugnaini, E.; Cerù, M.P. Immunocytochemical localization of catalase in the central nervous system of the rat. J. Histochem Cytochem. 1995, 43, 1253–1267. [Google Scholar] [CrossRef]
- Dunkin, G.; Laidlaw, P. Studies in dog-distemper: II.—Experimental distemper in the dog. J. Comp. Pathol. 1926, 39, 213–221. [Google Scholar] [CrossRef]
- Tipold, A.; Vandevelde, M.; Jaggy, A. Neurological manifestations of canine distemper virus infection. J. Small Anim Pr. 1992, 33, 466–470. [Google Scholar] [CrossRef]
- Carvalho, O.V.; Botelho, C.V.; Ferreira, C.G.T.; Scherer, P.O.; Soares-Martins, J.A.P.; Almeida, M.R.; Silva Júnior, A. Immunopathogenic and neurological mechanisms of canine distemper virus. Adv. Virus Res. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Amude, A.; Alfieri, A.; Alfieri, A. Canine distemper virus and multiple sclerosis: A real or an anecdotal association. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial. Biotechnology. Sao Paulo: Formatex, 2nd ed.; Mendez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2010; pp. 737–745. [Google Scholar]
- Beineke, A.; Puff, C.; Seehusen, F.; Baumgartner, W. Pathogenesis and immunopathology of systemic and nervous canine distemper. Vet. Immunol. Immunopathol. 2009, 127, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wyss-Fluehmann, G.; Zurbriggen, A.; Vandevelde, M.; Plattet, P. Canine distemper virus persistence in demyelinating encephalitis by swift intracellular cell-to-cell spread in astrocytes is controlled by the viral attachment protein. Acta Neuropathol. 2010, 119, 617–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alldinger, S.; Wünschmann, A.; Baumgärtner, W.; Voss, C.; Kremmer, E. Up-regulation of major histocompatibility complex class II antigen expression in the central nervous system of dogs with spontaneous canine distemper virus encephalitis. Acta Neuropathol. 1996, 92, 273–280. [Google Scholar] [CrossRef]
- Summers, B.; Greisen, H.; Appel, M. Early events in canine distemper demyelinating encephalomyelitis. Acta Neuropathol. 1979, 46, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lempp, C.; Spitzbarth, I.; Puff, C.; Cana, A.; Kegler, K.; Techangamsuwan, S.; Baumgartner, W.; Seehusen, F. New aspects of the pathogenesis of canine distemper leukoencephalitis. Viruses 2014, 6, 2571–2601. [Google Scholar] [CrossRef]
- Vandevelde, M.; Zurbriggen, A. Demyelination in canine distemper virus infection: A review. Acta Neuropathol. 2005, 109, 56–68. [Google Scholar] [CrossRef]
- Zurbriggen, A.; Schmid, I.; Graber, H.; Vandevelde, M. Oligodendroglial pathology in canine distemper. Acta Neuropathol. 1997, 95, 71–77. [Google Scholar] [CrossRef]
- Ulrich, R.; Puff, C.; Wewetzer, K.; Kalkuhl, A.; Deschl, U.; Baumgartner, W. Transcriptional changes in canine distemper virus-induced demyelinating leukoencephalitis favor a biphasic mode of demyelination. PLoS ONE 2014, 9, e95917. [Google Scholar] [CrossRef]
- Ozden, S.; Tangy, F.; Chamorro, M.; Brahic, M. Theiler’s virus genome is closely related to that of encephalomyocarditis virus, the prototype cardiovirus. J. Virol. 1986, 60, 1163–1165. [Google Scholar] [PubMed]
- Pevear, D.; Calenoff, M.; Rozhon, E.; Lipton, H. Analysis of the complete nucleotide sequence of the picornavirus Theiler’s murine encephalomyelitis virus indicates that it is closely related to cardioviruses. J. Virol. 1987, 61, 1507–1516. [Google Scholar] [PubMed]
- Oleszak, E.L.; Chang, J.R.; Friedman, H.; Katsetos, C.D.; Platsoucas, C.D. Theiler’s virus infection: A model for multiple sclerosis. Clin. Microbiol. Rev. 2004, 17, 174–207. [Google Scholar] [CrossRef] [PubMed]
- Theiler, M. Spontaneous Encephalomyelitis of Mice, A New Virus Disease. J. Exp. Med. 1937, 65, 705–719. [Google Scholar] [CrossRef] [PubMed]
- Mecha, M.; Carrillo-Salinas, F.J.; Mestre, L.; Feliu, A.; Guaza, C. Viral models of multiple sclerosis: Neurodegeneration and demyelination in mice infected with Theiler’s virus. Prog. Neurobiol. 2013, 101, 46–64. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, R.; Kalkuhl, A.; Deschl, U.; Baumgartner, W. Machine learning approach identifies new pathways associated with demyelination in a viral model of multiple sclerosis. J. Cell Mol. Med. 2010, 14, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Vandevelde, M.; Zurbriggen, A. The neurobiology of canine distemper virus infection. Vet. Microbiol. 1995, 44, 271–280. [Google Scholar] [CrossRef]
- Ulrich, R.; Seeliger, F.; Kreutzer, M.; Germann, P.G.; Baumgärtner, W. Limited remyelination in Theiler’s murine encephalomyelitis due to insufficient oligodendroglial differentiation of nerve/glial antigen 2 (NG2)-positive putative oligodendroglial progenitor cells. Neuropathol. Appl. Neurobiol. 2008, 34, 603–620. [Google Scholar] [CrossRef]
- Fischer, M.T.; Sharma, R.; Lim, J.L.; Haider, L.; Frischer, J.M.; Drexhage, J.; Mahad, D.; Bradl, M.; van Horssen, J.; Lassmann, H. NADPH oxidase expression in active multiple sclerosis lesions in relation to oxidative tissue damage and mitochondrial injury. Brain 2012, 135, 886–899. [Google Scholar] [CrossRef] [Green Version]
- Bains, M.; Hall, E.D. Antioxidant therapies in traumatic brain and spinal cord injury. Biochim. Biophys Acta 2012, 1822, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Denic, A.; Johnson, A.J.; Bieber, A.J.; Warrington, A.E.; Rodriguez, M.; Pirko, I. The relevance of animal models in multiple sclerosis research. Pathophysiology 2011, 18, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Procaccini, C.; De Rosa, V.; Pucino, V.; Formisano, L.; Matarese, G. Animal models of multiple sclerosis. Eur. J. Pharm. 2015, 759, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Koestner, A. Animal model of human disease: Subacute sclerosing panencephalitis, multiple sclerosis; animal model: Distemper-associated demyelinating encephalomyelitis. Am. J. Pathol. 1975, 78, 361. [Google Scholar] [PubMed]
- Schuh, C.; Wimmer, I.; Hametner, S.; Haider, L.; Van Dam, A.M.; Liblau, R.S.; Smith, K.J.; Probert, L.; Binder, C.J.; Bauer, J.; et al. Oxidative tissue injury in multiple sclerosis is only partly reflected in experimental disease models. Acta Neuropathol. 2014, 128, 247–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Canto, M.C.; Rabinowitz, S.G. Experimental models of virus-induced demyelination of the central nervous system. Ann. Neurol. 1982, 11, 109–127. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, K.B. Oxidative stress during viral infection: A review. Free Radic Biol. Med. 1996, 21, 641–649. [Google Scholar] [CrossRef]
- Chang, J.R.; Zaczynska, E.; Katsetos, C.D.; Platsoucas, C.D.; Oleszak, E.L. Differential expression of TGF-β, IL-2, and other cytokines in the CNS of Theiler’s murine encephalomyelitis virus-infected susceptible and resistant strains of mice. Virology 2000, 278, 346–360. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Mao, P.; Reddy, P.H. Is multiple sclerosis a mitochondrial disease? Biochim Biophys Acta 2010, 1802, 66–79. [Google Scholar] [CrossRef]
- Bokoch, G.M. Regulation of innate immunity by Rho GTPases. Trends Cell Biol. 2005, 15, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Asahara, S.; Shibutani, Y.; Teruyama, K.; Inoue, H.; Kawada, Y.; Etoh, H.; Matsuda, T.; Kimura-Koyanagi, M.; Hashimoto, N.; Sakahara, M. Ras-related C3 botulinum toxin substrate 1 (RAC1) regulates glucose-stimulated insulin secretion via modulation of F-actin. Diabetologia 2013, 56, 1088–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridley, A.J. Rho GTPases and cell migration. J. Cell Sci. 2001, 114, 2713–2722. [Google Scholar] [PubMed]
- Bustelo, X.R.; Sauzeau, V.; Berenjeno, I.M. GTP-binding proteins of the Rho/Rac family: Regulation, effectors and functions in vivo. Bioessays 2007, 29, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Gaasch, J.A.; Lockman, P.R.; Geldenhuys, W.J.; Allen, D.D.; Van der Schyf, C.J. Brain iron toxicity: Differential responses of astrocytes, neurons, and endothelial cells. Neurochem. Res. 2007, 32, 1196–1208. [Google Scholar] [CrossRef] [PubMed]
- Sadrzadeh, S.M.; Saffari, Y. Iron and brain disorders. Am. J. Clin. Pathol. 2004, 121, S64–S70. [Google Scholar] [CrossRef]
- Lassmann, H.; van Horssen, J.; Mahad, D. Progressive multiple sclerosis: Pathology and pathogenesis. Nat. Rev. Neurosci 2012, 8, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef]
- Mahad, D.J.; Ziabreva, I.; Campbell, G.; Lax, N.; White, K.; Hanson, P.S.; Lassmann, H.; Turnbull, D.M. Mitochondrial changes within axons in multiple sclerosis. Brain 2009, 132, 1161–1174. [Google Scholar] [CrossRef] [Green Version]
- Mahad, D.; Ziabreva, I.; Lassmann, H.; Turnbull, D. Mitochondrial defects in acute multiple sclerosis lesions. Brain 2008, 131, 1722–1735. [Google Scholar] [CrossRef]
- Kirkinezos, I.G.; Moraes, C.T. Reactive oxygen species and mitochondrial diseases. Semin. Cell Dev. Biol. 2001, 12, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Schad, A.; Fahimi, H.D.; Völkl, A.; Baumgart, E. Expression of catalase mRNA and protein in adult rat brain: Detection by nonradioactive in situ hybridization with signal amplification by catalyzed reporter deposition (ISH–CARD) and immunohistochemistry (IHC)/immunofluorescence (IF). J. Histochem. Cytochem. 2003, 51, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Guy, J.; Ellis, E.A.; Hope, G.M.; Rao, N.A. Antioxidant enzyme suppression of demyelination in experimental optic neuritis. Curr. Eye Res. 1989, 8, 467–477. [Google Scholar] [CrossRef] [PubMed]
- van Horssen, J.; Schreibelt, G.; Drexhage, J.; Hazes, T.; Dijkstra, C.D.; van der Valk, P.; de Vries, H.E. Severe oxidative damage in multiple sclerosis lesions coincides with enhanced antioxidant enzyme expression. Free Radic. Biol. Med. 2008, 45, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- Seehusen, F.; Orlando, E.A.; Wewetzer, K.; Baumgärtner, W. Vimentin-positive astrocytes in canine distemper: A target for canine distemper virus especially in chronic demyelinating lesions? Acta Neuropathol. 2007, 114, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lehmbecker, A.; Kalkuhl, A.; Deschl, U.; Sun, W.; Rohn, K.; Tzvetanova, I.D.; Nave, K.A.; Baumgärtner, W.; Ulrich, R. STAT3 represents a molecular switch possibly inducing astroglial instead of oligodendroglial differentiation of oligodendroglial progenitor cells in Theiler’s murine encephalomyelitis. Neuropathol. Appl. Neurobiol. 2015, 41, 347–370. [Google Scholar] [CrossRef] [PubMed]
- Olby, N. The pathogenesis and treatment of acute spinal cord injuries in dogs. Vet. Clin. North. Am. Food Anim Pr. 2010, 40, 791–807. [Google Scholar] [CrossRef]
- Freundt-Revilla, J.; Kegler, K.; Baumgärtner, W.; Tipold, A. Spatial distribution of cannabinoid receptor type 1 (CB1) in normal canine central and peripheral nervous system. PLoS ONE 2017, 12, e0181064. [Google Scholar] [CrossRef]
- Uhde, A.-K.; Herder, V.; Khan, M.A.; Ciurkiewicz, M.; Schaudien, D.; Teich, R.; Floess, S.; Baumgärtner, W.; Huehn, J.; Beineke, A. Viral Infection of the Central Nervous System Exacerbates Interleukin-10 Receptor Deficiency-Mediated Colitis in SJL Mice. PLoS ONE 2016, 11, e0161883. [Google Scholar] [CrossRef]
- Bolstad, B.M.; Irizarry, R.A.; Åstrand, M.; Speed, T.P. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Raddatz, B.B.; Spitzbarth, I.; Matheis, K.A.; Kalkuhl, A.; Deschl, U.; Baumgärtner, W.; Ulrich, R. Microarray-based gene expression analysis for veterinary pathologists: A review. Vet. Pathol. 2017, 54, 734–755. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, M.; Lepow, T.S.; Billingham, L.K.; Murphy, M.P.; Siegel, R.M. New tricks from an old dog: Mitochondrial redox signaling in cellular inflammation. Semin Immunol. 2012, 24, 384–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, G.K.; Scaini, G.; Carvalho-Silva, M.; Gomes, L.M.; Borges, L.S.; Vieira, J.S.; Constantino, L.S.; Ferreira, G.C.; Schuck, P.F.; Streck, E.L. Effect of l-Tyrosine In Vitro and In Vivo on Energy Metabolism Parameters in Brain and Liver of Young Rats. Neurotox. Res. 2013, 23, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Adam-Vizi, V. Production of reactive oxygen species in brain mitochondria: Contribution by electron transport chain and non–electron transport chain sources. Antioxid Redox Signal. 2005, 7, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Mastrogiacoma, F.; Lindsay, J.G.; Bettendorff, L.; Rice, J.; Kish, S.J. Brain protein and alpha-ketoglutarate dehydrogenase complex activity in Alzheimer’s disease. Ann. Neurol. 1996, 39, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Wynn, R.M.; Chuang, J.L.; Tso, S.-C.; Machius, M.; Li, J.; Chuang, D.T. Structural basis for inactivation of the human pyruvate dehydrogenase complex by phosphorylation: Role of disordered phosphorylation loops. Structure 2008, 16, 1849–1859. [Google Scholar] [CrossRef]
- Michel, H. The mechanism of proton pumping by cytochrome c oxidasex127e [comments]. Proc. Natl. Acad Sci. USA 1998, 95, 12819–12824. [Google Scholar] [CrossRef]
- Michel, H.; Behr, J.; Harrenga, A.; Kannt, A. Cytochrome c oxidase: Structure and spectroscopy. Annu. Rev. Biophys Biomol. Struct. 1998, 27, 329–356. [Google Scholar] [CrossRef]
- Wikstrom, M. Proton translocation by cytochrome c oxidase: A rejoinder to recent criticism. Biochemistry 2000, 39, 3515–3519. [Google Scholar] [CrossRef]
- van Horssen, J.; Witte, M.E.; Schreibelt, G.; de Vries, H.E. Radical changes in multiple sclerosis pathogenesis. Biochim. Biophys Acta Mol. Basis Dis. 2011, 1812, 141–150. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Genes (N) | Differentially Expressed Genes | |||
|---|---|---|---|---|---|
| Direction of change | 2 vs. 1 | 3 vs. 1 | 4 vs. 1 | ||
| Mitochondrial respiratory chain | 50 | Up | 0 | 0 | 0 |
| Down | 7 (14%) | 17 (34%) | 6 (12%) | ||
| NADPH oxidase enzyme system | 13 | Up | 5 (38.5%) | 4 (30.8%) | 2 (15.38%) |
| Down | 0 | 0 | 0 | ||
| ROS detoxification system | 12 | Up | 3 (25%) | 3 (25%) | 1 (8.33%) |
| Down | 1 (8.33%) | 1 (8.33%) | 0 | ||
| Catabolic enzymes with oxidase activity | 3 | Up | 1 (33.33%) | 1 (33.33%) | 1 (33.33%) |
| Down | 0 | 1 (33.33%) | 1 (33.33%) | ||
| Pyruvate dehydrogenase complex) | 5 | Up | 0 | 0 | 0 |
| Down | 0 | 1 (20%) | 0 | ||
| ROS toxification system | 4 | Up | 0 | 0 | 0 |
| Down | 0 | 0 | 0 |
| Function | Genes (N) | Differentially Expressed Genes | ||||
|---|---|---|---|---|---|---|
| Direction of change | 14dpi | 42dpi | 98dpi | 196dpi | ||
| Mitochondrial respiratory chain | 55 | Up | 0 | 0 | 0 | 0 |
| Down | 7(12.7%) | 31(56.36%) | 30(54.55%) | 36 (65.45%) | ||
| NADPH oxidase enzyme system | 9 | Up | 5 (55.5%) | 6 (66.7%) | 6 (66.7%) | 6 (66.7%) |
| Down | 0 | 0 | 0 | 0 | ||
| ROS detoxification system | 17 | Up | 0 | 2 (11.76%) | 1 (5.88%) | 4 (23.53%) |
| Down | 1 (5.88%) | 2 (11.76%) | 3 (17.65%) | 3 (17.65%) | ||
| Catabolic enzymes with oxidase activity | 3 | Up | 1 (33.3%) | 2 (66.7%) | 2 (66.7%) | 1 (33.3%) |
| Down | 0 | 1 (33.3%) | 0 | 0 | ||
| Pyruvate dehydrogenase complex) | 5 | Up | 0 | 0 | 0 | 0 |
| Down | 0 | 3 (60%) | 2 (40%) | 0 | ||
| ROS toxification system | 4 | Up | 0 | 0 | 0 | 0 |
| Down | 0 | 0 | 1 (25.0%) | 1 (25.0%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attig, F.; Spitzbarth, I.; Kalkuhl, A.; Deschl, U.; Puff, C.; Baumgärtner, W.; Ulrich, R. Reactive Oxygen Species Are Key Mediators of Demyelination in Canine Distemper Leukoencephalitis but not in Theiler’s Murine Encephalomyelitis. Int. J. Mol. Sci. 2019, 20, 3217. https://doi.org/10.3390/ijms20133217
Attig F, Spitzbarth I, Kalkuhl A, Deschl U, Puff C, Baumgärtner W, Ulrich R. Reactive Oxygen Species Are Key Mediators of Demyelination in Canine Distemper Leukoencephalitis but not in Theiler’s Murine Encephalomyelitis. International Journal of Molecular Sciences. 2019; 20(13):3217. https://doi.org/10.3390/ijms20133217
Chicago/Turabian StyleAttig, Friederike, Ingo Spitzbarth, Arno Kalkuhl, Ulrich Deschl, Christina Puff, Wolfgang Baumgärtner, and Reiner Ulrich. 2019. "Reactive Oxygen Species Are Key Mediators of Demyelination in Canine Distemper Leukoencephalitis but not in Theiler’s Murine Encephalomyelitis" International Journal of Molecular Sciences 20, no. 13: 3217. https://doi.org/10.3390/ijms20133217
APA StyleAttig, F., Spitzbarth, I., Kalkuhl, A., Deschl, U., Puff, C., Baumgärtner, W., & Ulrich, R. (2019). Reactive Oxygen Species Are Key Mediators of Demyelination in Canine Distemper Leukoencephalitis but not in Theiler’s Murine Encephalomyelitis. International Journal of Molecular Sciences, 20(13), 3217. https://doi.org/10.3390/ijms20133217