Scratching Counteracts IL-13 Signaling by Upregulating the Decoy Receptor IL-13Rα2 in Keratinocytes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
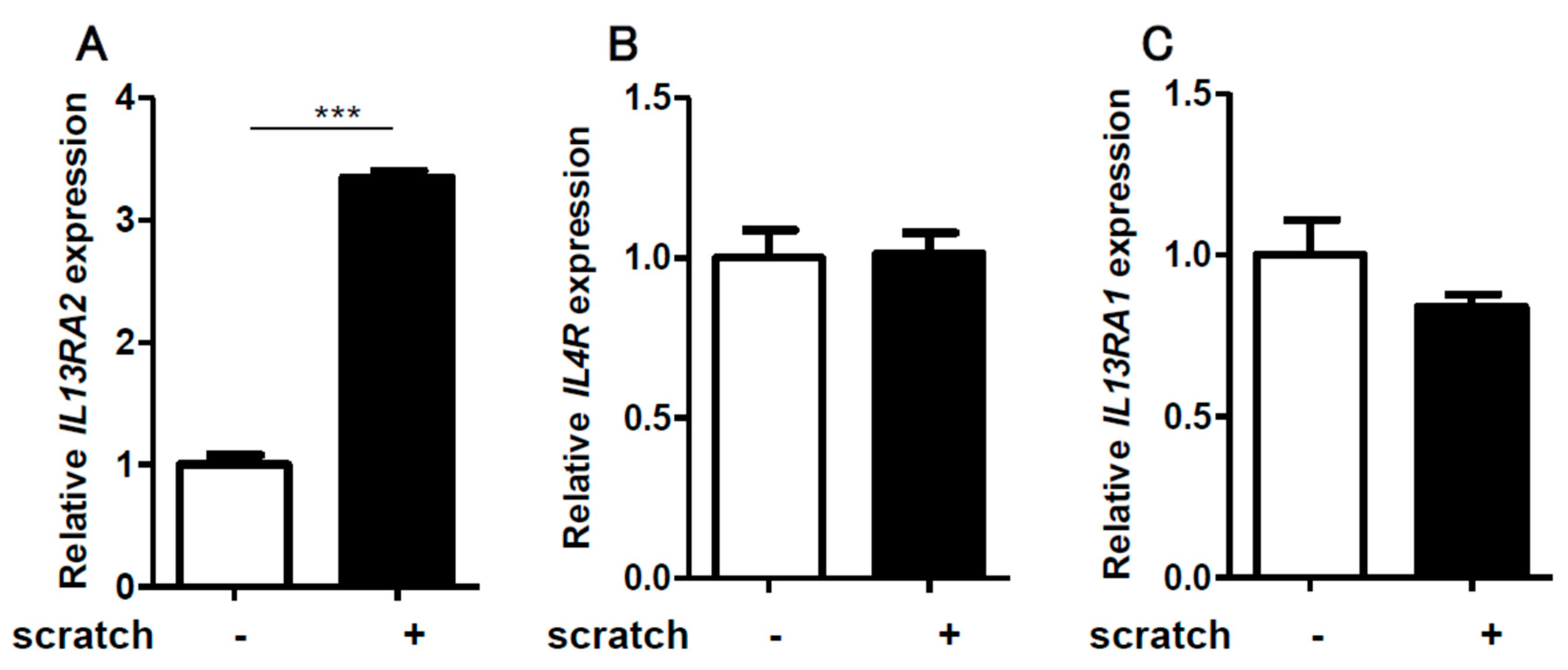
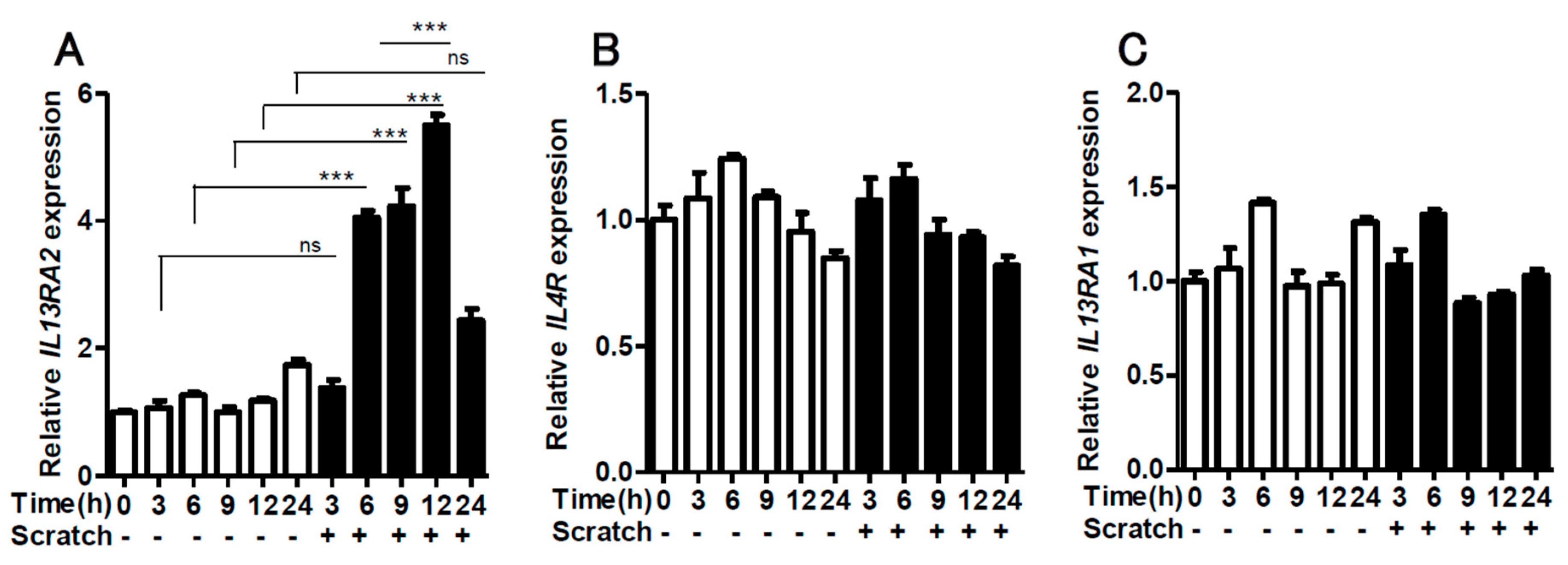
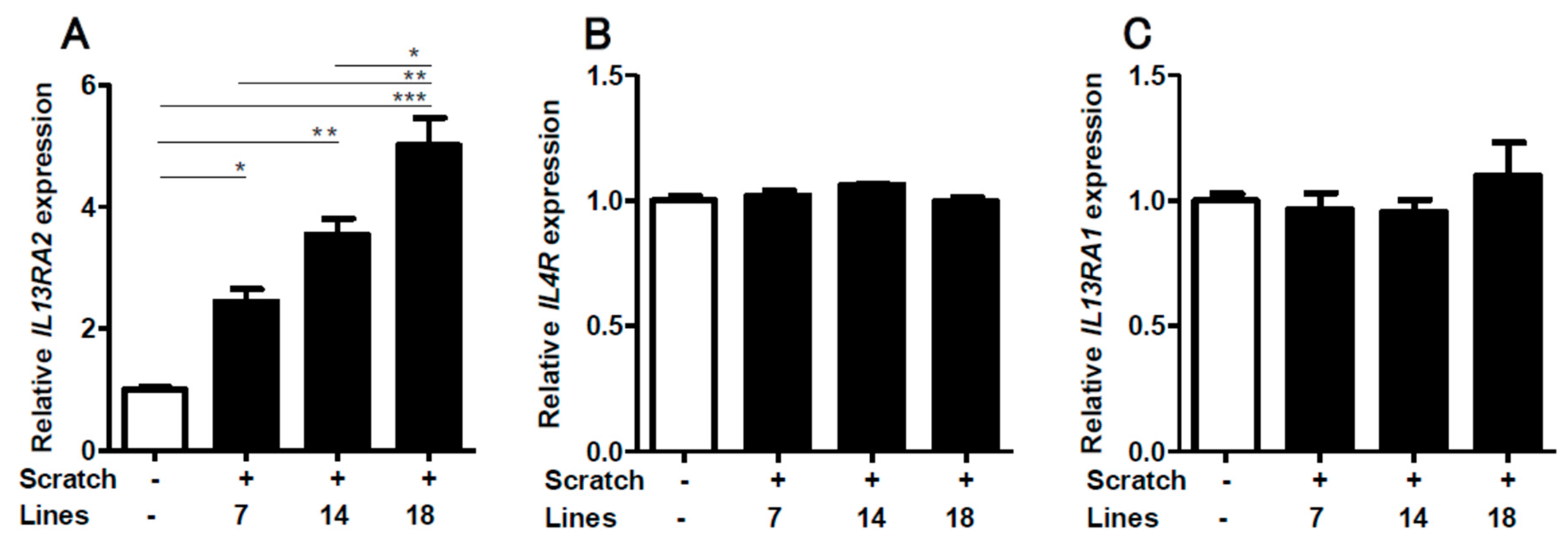
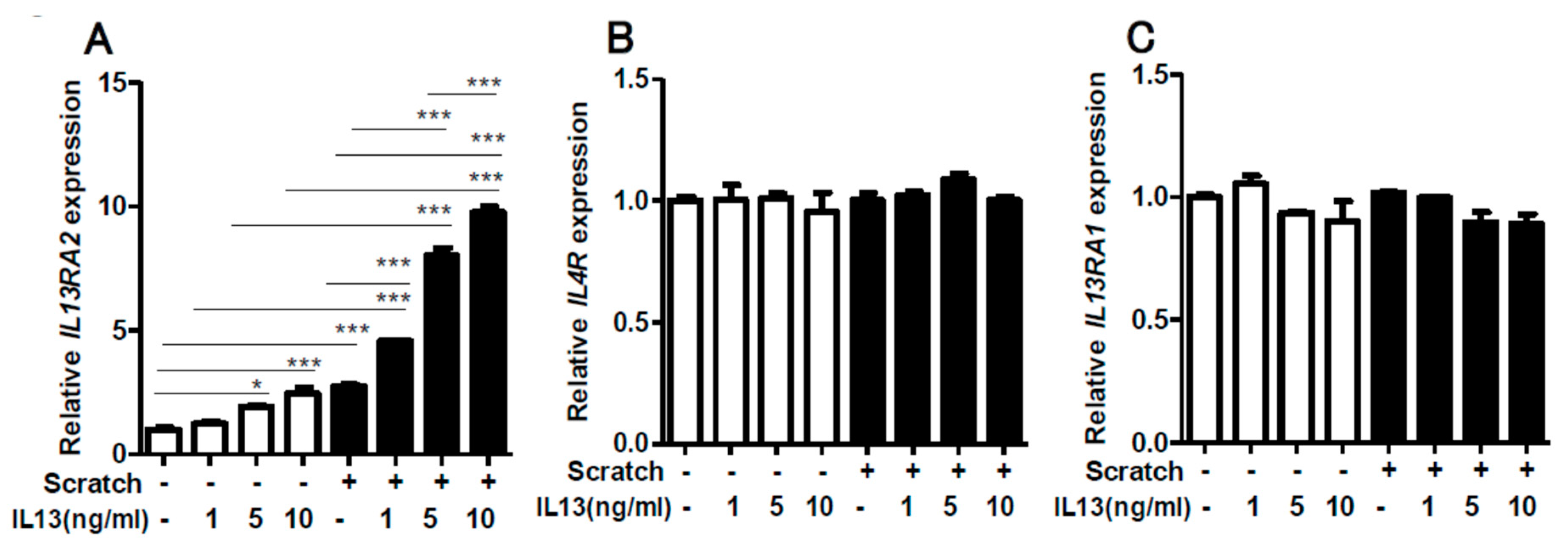
2.1. Scratching Upregulates the Expression of IL13RA2, Which is Further Enhanced by IL-13
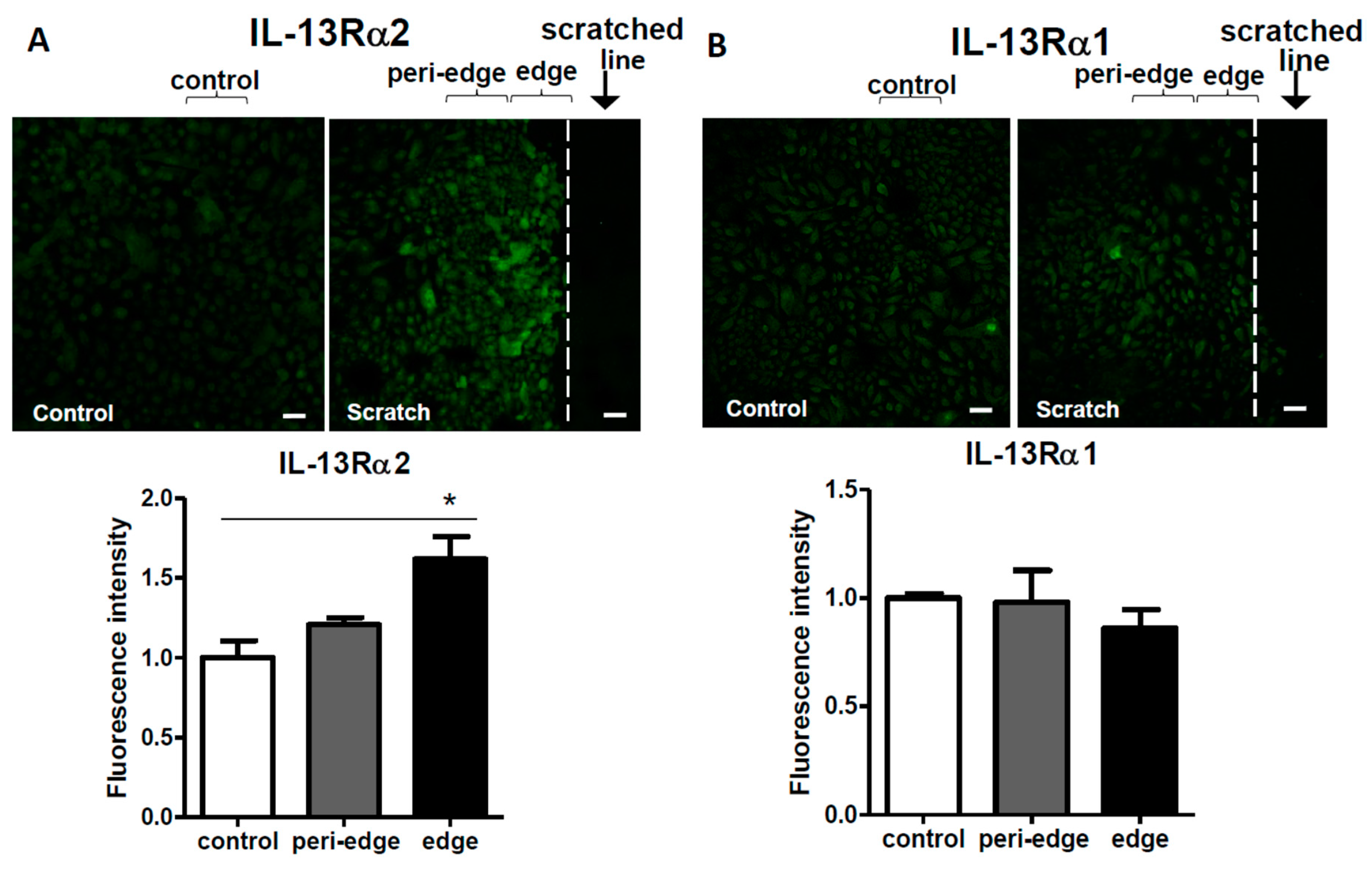
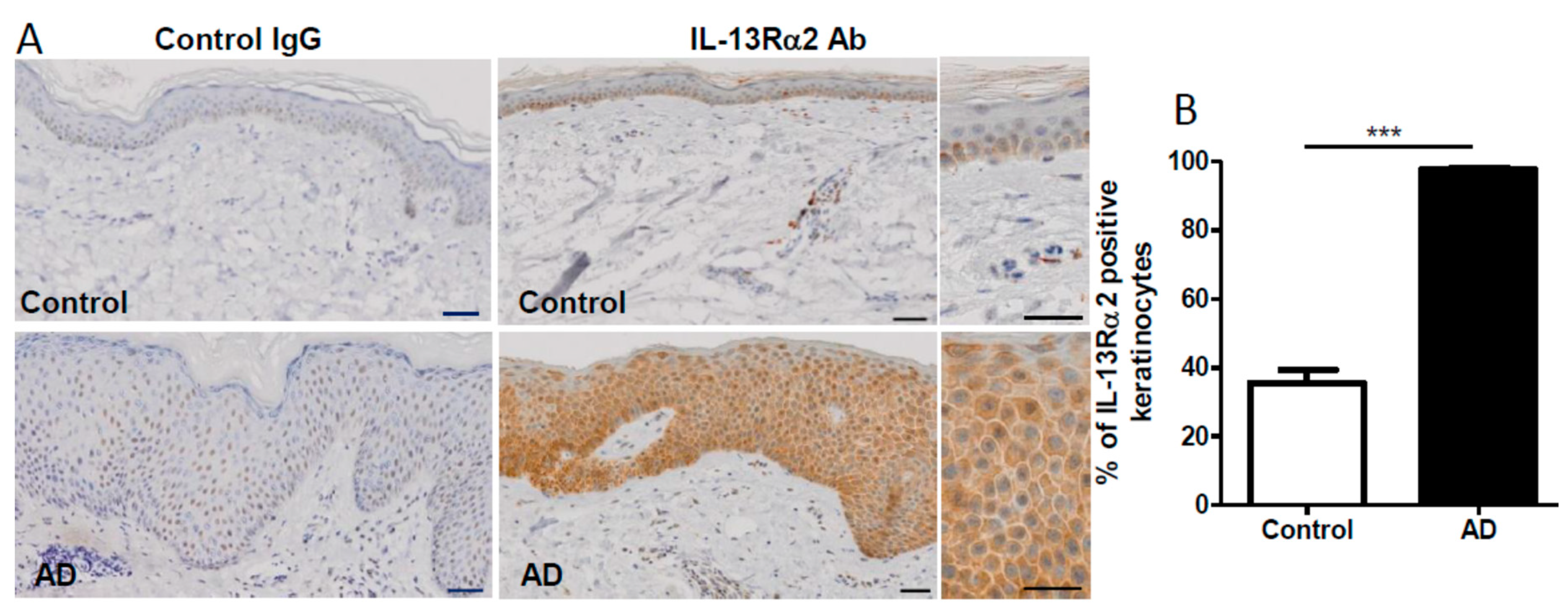
2.2. Upregulation of IL-13Rα2 Protein in a Scratched Edge Area In Vitro as well as in Lesional AD Skin
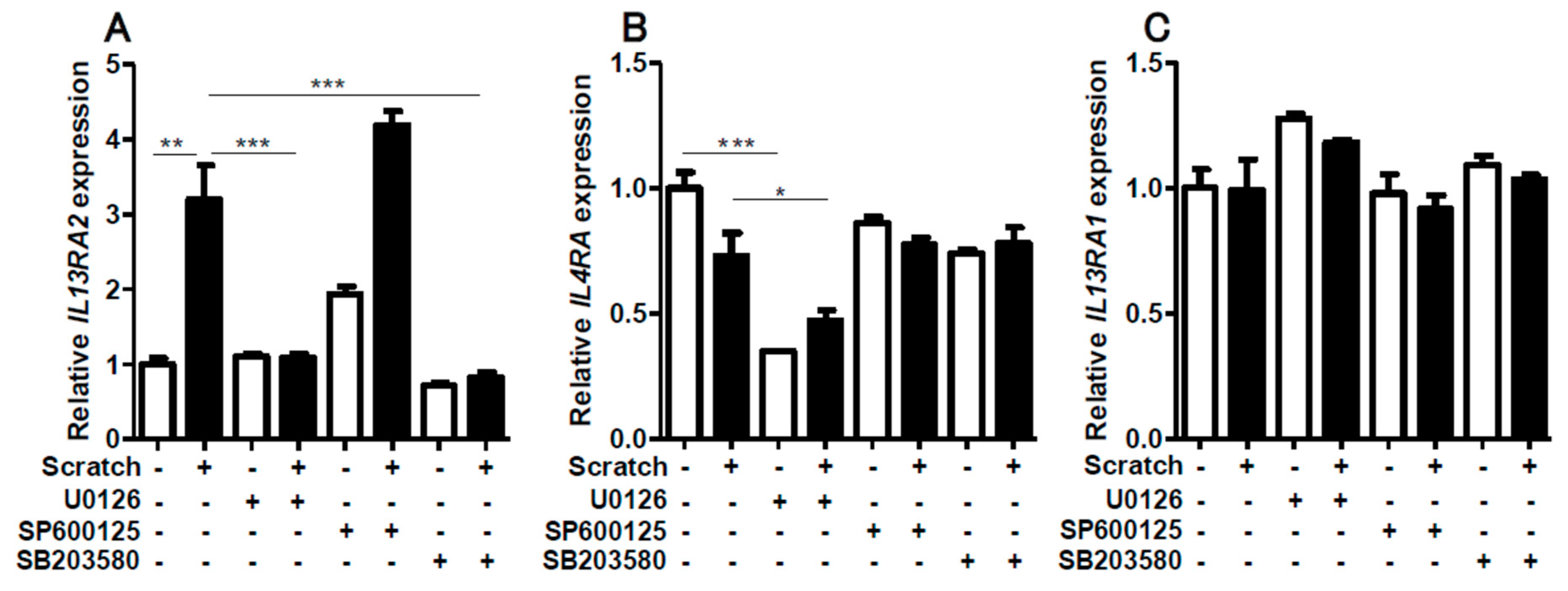
2.3. Contribution of ERK1/2 and P38MAPK Activation to Scratch-Induced IL13RA2 Upregulation
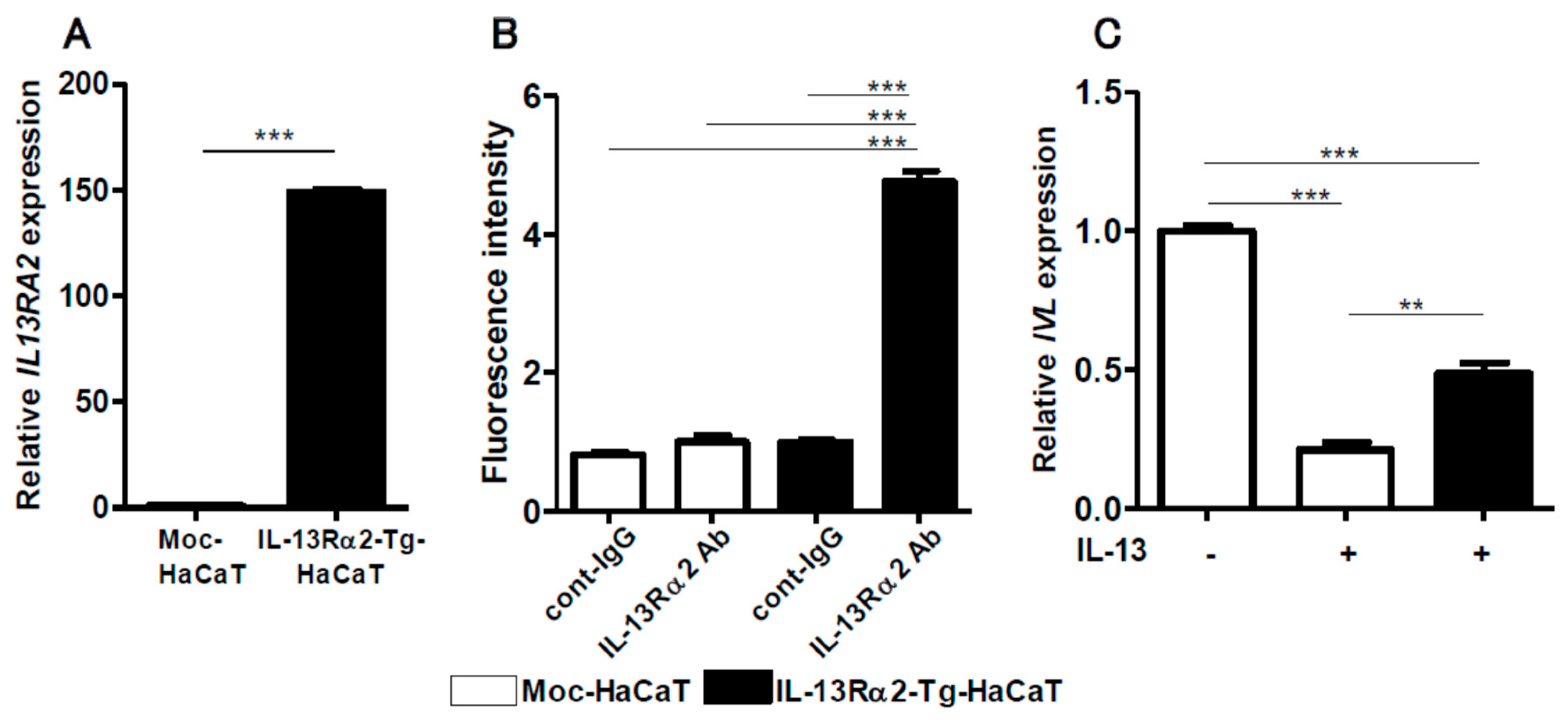
2.4. IL-13-Mediated IVL Downregulation is Restored in IL-13Rα2-Overexpressed HaCaT Keratinocytes
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture
4.3. In Vitro Scratched Keratinocyte Model
4.4. Quantitative Real-Time PCR (qRT-PCR)
4.5. Immunofluorescence Analysis
4.6. Immunohistochemistry
4.7. Western Blotting
4.8. Plasmid DNA and Transfection of Plasmids
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Conflicts of Interest
Abbreviations
| AD | atopic dermatitis |
| ERK | extracellular signal-regulated kinase |
| FLG | filaggrin |
| IL | interleukin |
| IL-4Rα | IL-4 receptor α |
| IL-13Rα1 | IL-13 receptor α1 |
| IL-13Rα2 | IL-13 receptor α2 |
| IVL | involucrin |
| JNK | c-Jun N-terminal kinase |
| MAPK | mitogen-activated protein kinase |
| MEK | mitogen-activated protein kinase kinase |
| NHEK | normal human epidermal keratinocyte |
| TH2 cell | T helper 2 cells |
References
- Bieber, T. Atopic dermatitis 2.0: From the clinical phenotype to the molecular taxonomy and stratified medicine. Allergy 2012, 67, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Chiba, T.; Tsuji, G.; Ulzii, D.; Kido-Nakahara, M.; Nakahara, T.; Kadono, T. Atopic dermatitis: Immune deviation, barrier dysfunction, IgE autoreactivity and new therapies. Allergol. Int. 2017, 66, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, J.; Apfelbacher, C.; Spuls, P.I.; Thomas, K.S.; Simpson, E.L.; Furue, M.; Chalmers, J.; Williams, H.C. The Harmonizing Outcome Measures for Eczema (HOME) roadmap: A methodological framework to develop core sets of outcome measurements in dermatology. J. Invest. Dermatol. 2015, 135, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.L.; Bruin-Weller, M.; Flohr, C.; Ardern-Jones, M.R.; Barbarot, S.; Deleuran, M.; Bieber, T.; Vestergaard, C.; Brown, S.J.; Cork, M.J.; et al. When does atopic dermatitis warrant systemic therapy? Recommendations from an expert panel of the International Eczema Council. J. Am. Acad. Dermatol. 2017, 77, 623–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, M.; Chiba, T.; Takeuchi, S. Current status of atopic dermatitis in Japan. Asia Pac. Allergy 2011, 1, 64–72. [Google Scholar] [CrossRef]
- Williams, H.; Stewart, A.; von Mutius, E.; Cookson, W.; Anderson, H.R. International Study of Asthma and Allergies in Childhood (ISAAC) Phase One and Three Study Groups. Is eczema really on the increase worldwide? J. Allergy Clin. Immunol. 2008, 121, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Saeki, H.; Nakahara, T.; Tanaka, A.; Kabashima, K.; Sugaya, M.; Murota, H.; Ebihara, T.; Kataoka, Y.; Aihara, M.; Etoh, T.; et al. Committee for Clinical Practice Guidelines for the Management of Atopic Dermatitis of Japanese Dermatological Association. Clinical Practice Guidelines for the Management of Atopic Dermatitis 2016. J. Dermatol. 2016, 43, 1117–1145. [Google Scholar] [CrossRef]
- Pavlis, J.; Yosipovitch, G. Management of Itch in Atopic Dermatitis. Am. J. Clin. Dermatol. 2018, 19, 319–332. [Google Scholar] [CrossRef]
- Takeuchi, S.; Yasukawa, F.; Furue, M.; Katz, S.I. Collared mice: A model to assess the effects of scratching. J. Dermatol. Sci. 2010, 57, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Hamid, Q.; Boguniewicz, M.; Leung, D.Y. Differential in situ cytokine gene expression in acute versus chronic atopic dermatitis. J. Clin. Invest. 1994, 94, 870–876. [Google Scholar] [CrossRef]
- Czarnowicki, T.; Esaki, H.; Gonzalez, J.; Malajian, D.; Shemer, A.; Noda, S.; Talasila, S.; Berry, A.; Gray, J.; Becker, L.; et al. Early pediatric atopic dermatitis shows only a cutaneous lymphocyte antigen (CLA)(+) TH2/TH1 cell imbalance, whereas adults acquire CLA(+) TH22/TC22 cell subsets. J. Allergy Clin. Immunol. 2015, 136, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Esaki, H.; Brunner, P.M.; Renert-Yuval, Y.; Czarnowicki, T.; Huynh, T.; Tran, G.; Lyon, S.; Rodriguez, G.; Immaneni, S.; Johnson, D.B.; et al. Early-onset pediatric atopic dermatitis is T(H)2 but also T(H)17 polarized in skin. J. Allergy Clin. Immunol. 2016, 138, 1639–1651. [Google Scholar] [CrossRef] [PubMed]
- Gittler, J.K.; Shemer, A.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Gulewicz, K.J.; Wang, C.Q.; Mitsui, H.; Cardinale, I.; de Guzman Strong, C.; Krueger, J.G.; et al. Progressive activation of T(H)2/T(H)22 cytokines and selective epidermal proteins characterizes acute and chronic atopic dermatitis. J. Allergy Clin. Immunol. 2012, 130, 1344–1354. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, G.; Hashimoto-Hachiya, A.; Kiyomatsu-Oda, M.; Takemura, M.; Ohno, F.; Ito, T.; Morino-Koga, S.; Mitoma, C.; Nakahara, T.; Uchi, H.; et al. Aryl hydrocarbon receptor activation restores filaggrin expression via OVOL1 in atopic dermatitis. Cell Death Dis. 2017, 8, e2931. [Google Scholar] [CrossRef] [PubMed]
- Van den Bogaard, E.H.; Bergboer, J.G.; Vonk-Bergers, M.; van Vlijmen-Willems, I.M.; Hato, S.V.; van der Valk, P.G.; Schröder, J.M.; Joosten, I.; Zeeuwen, P.L.; Schalkwijk, J. Coal tar induces AHR-dependent skin barrier repair in atopic dermatitis. J. Clin. Invest. 2013, 123, 917–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oetjen, L.K.; Mack, M.R.; Feng, J.; Whelan, T.M.; Niu, H.; Guo, C.J.; Chen, S.; Trier, A.M.; Xu, A.Z.; Tripathi, S.V.; et al. Sensory Neurons Co-opt Classical Immune Signaling Pathways to Mediate Chronic Itch. Cell 2017, 171, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.L.; Bieber, T.; Guttman-Yassky, E.; Beck, L.A.; Blauvelt, A.; Cork, M.J.; Silverberg, J.I.; Deleuran, M.; Kataoka, Y.; Lacour, J.P.; et al. SOLO 1 and SOLO 2 Investigators. Two Phase 3 Trials of Dupilumab versus Placebo in Atopic Dermatitis. N. Engl. J. Med. 2016, 375, 2335–2348. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, L.C.; Rodriguez, E.; Degenhardt, F.; Baurecht, H.; Wehkamp, U.; Volks, N.; Szymczak, S.; Swindell, W.R.; Sarkar, M.K.; Raja, K.; et al. Atopic dermatitis is an IL-13 dominant disease with greater molecular heterogeneity compared to psoriasis. J. Invest. Dermatol. 2019, 139, 1480–1489. [Google Scholar] [CrossRef]
- Lumsden, R.V.; Worrell, J.C.; Boylan, D.; Walsh, S.M.; Cramton, J.; Counihan, I.; O’Beirne, S.; Medina, M.F.; Gauldie, J.; Fabre, A.; et al. Modulation of pulmonary fibrosis by IL-13Rα2. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 308, L710–L718. [Google Scholar] [CrossRef]
- Ranasinghe, C.; Trivedi, S.; Wijesundara, D.K.; Jackson, R.J. IL-4 and IL-13 receptors: Roles in immunity and powerful vaccine adjuvants. Cytokine Growth Factor Rev. 2014, 25, 437–442. [Google Scholar] [CrossRef]
- Kasaian, M.T.; Raible, D.; Marquette, K.; Cook, T.A.; Zhou, S.; Tan, X.Y.; Tchistiakova, L. IL-13 antibodies influence IL-13 clearance in humans by modulating scavenger activity of IL-13Rα2. J. Immunol. 2011, 187, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.; Whitters, M.J.; Jacobson, B.A.; Witek, J.; Sypek, J.P.; Kasaian, M.; Eppihimer, M.J.; Unger, M.; Tanaka, T.; Goldman, S.J.; et al. Enhanced interleukin (IL)-13 responses in mice lacking IL-13 receptor alpha 2. J. Exp. Med. 2003, 197, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Akaiwa, M.; Yu, B.; Umeshita-Suyama, R.; Terada, N.; Suto, H.; Koga, T.; Arima, K.; Matsushita, S.; Saito, H.; Ogawa, H.; et al. Localization of human interleukin 13 receptor in non-haematopoietic cells. Cytokine 2001, 13, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Sivaprasad, U.; Warrier, M.R.; Gibson, A.M.; Chen, W.; Tabata, Y.; Bass, S.A.; Rothenberg, M.E.; Khurana Hershey, G.K. IL-13Rα2 has a protective role in a mouse model of cutaneous inflammation. J. Immunol. 2010, 185, 6802–6808. [Google Scholar] [CrossRef] [PubMed]
- Kido-Nakahara, M.; Furue, M.; Ulzii, D.; Nakahara, T. Itch in Atopic Dermatitis. Immunol. Allergy Clin. N. Am. 2017, 37, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Hadi, S.; Guttman-Yassky, E. Current and emerging biologic and small molecule therapies for atopic dermatitis. Expert. Opin. Biol. Ther. 2019, 19, 367–380. [Google Scholar] [CrossRef]
- Vierow, V.; Forster, C.; Vogelgsang, R.; Dörfler, A.; Handwerker, H.O. Cerebral Networks Linked to Itch-related Sensations Induced by Histamine and Capsaicin. Acta. Derm. Venereol. 2015, 95, 645–652. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.; Onozuka, D.; Takeuchi, S.; Murota, H.; Sugaya, M.; Masuda, K.; Hiragun, T.; Kaneko, S.; Saeki, H.; Shintani, Y.; et al. Poor adherence to oral and topical medication in 3096 dermatological patients as assessed by the Morisky Medication Adherence Scale-8. Br. J. Dermatol. 2015, 172, 272–275. [Google Scholar] [CrossRef]
- Kabashima, K.; Furue, M.; Hanifin, J.M.; Pulka, G.; Wollenberg, A.; Galus, R.; Etoh, T.; Mihara, R.; Nakano, M.; Ruzicka, T. Nemolizumab in patients with moderate-to-severe atopic dermatitis: Randomized, phase II, long-term extension study. J. Allergy Clin. Immunol. 2018, 142, 1121–1130. [Google Scholar] [CrossRef]
- Nakahara, T.; Fujita, H.; Arima, K.; Taguchi, Y.; Motoyama, S.; Furue, M. Treatment satisfaction in atopic dermatitis relates to patient-reported severity: A cross-sectional study. Allergy 2018, 74, 1179–1181. [Google Scholar] [CrossRef]
- Morino-Koga, S.; Uchi, H.; Mitoma, C.; Wu, Z.; Kiyomatsu, M.; Fuyuno, Y.; Nagae, K.; Yasumatsu, M.; Suico, M.A.; Kai, H.; et al. 6-Formylindolo[3,2-b]Carbazole Accelerates Skin Wound Healing via Activation of ERK, but Not Aryl Hydrocarbon Receptor. J. Invest. Dermatol. 2017, 137, 2217–2226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasca, M.R.; O’Toole, E.A.; Palicharla, P.; West, D.P.; Woodley, D.T. Thalidomide increases human keratinocyte migration and proliferation. J. Invest. Dermatol. 1999, 113, 720–724. [Google Scholar] [PubMed]
- Patruno, A.; Ferrone, A.; Costantini, E.; Franceschelli, S.; Pesce, M.; Speranza, L.; Amerio, P.; D’Angelo, C.; Felaco, M.; Grilli, A.; et al. Extremely low-frequency electromagnetic fields accelerates wound healing modulating MMP-9 and inflammatory cytokines. Cell Prolif. 2018, 51, e12432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, M.D.; Kim, B.E.; Gao, P.; Grant, A.V.; Boguniewicz, M.; DeBenedetto, A.; Schneider, L.; Beck, L.A.; Barnes, K.C.; Leung, D.Y. Cytokine modulation of atopic dermatitis filaggrin skin expression. J. Allergy Clin. Immunol. 2009, 124, R7–R12. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulzii, D.; Kido-Nakahara, M.; Nakahara, T.; Tsuji, G.; Furue, K.; Hashimoto-Hachiya, A.; Furue, M. Scratching Counteracts IL-13 Signaling by Upregulating the Decoy Receptor IL-13Rα2 in Keratinocytes. Int. J. Mol. Sci. 2019, 20, 3324. https://doi.org/10.3390/ijms20133324
Ulzii D, Kido-Nakahara M, Nakahara T, Tsuji G, Furue K, Hashimoto-Hachiya A, Furue M. Scratching Counteracts IL-13 Signaling by Upregulating the Decoy Receptor IL-13Rα2 in Keratinocytes. International Journal of Molecular Sciences. 2019; 20(13):3324. https://doi.org/10.3390/ijms20133324
Chicago/Turabian StyleUlzii, Dugarmaa, Makiko Kido-Nakahara, Takeshi Nakahara, Gaku Tsuji, Kazuhisa Furue, Akiko Hashimoto-Hachiya, and Masutaka Furue. 2019. "Scratching Counteracts IL-13 Signaling by Upregulating the Decoy Receptor IL-13Rα2 in Keratinocytes" International Journal of Molecular Sciences 20, no. 13: 3324. https://doi.org/10.3390/ijms20133324
APA StyleUlzii, D., Kido-Nakahara, M., Nakahara, T., Tsuji, G., Furue, K., Hashimoto-Hachiya, A., & Furue, M. (2019). Scratching Counteracts IL-13 Signaling by Upregulating the Decoy Receptor IL-13Rα2 in Keratinocytes. International Journal of Molecular Sciences, 20(13), 3324. https://doi.org/10.3390/ijms20133324