Kidney Injury Molecule-1 Is Upregulated in Renal Lipotoxicity and Mediates Palmitate-Induced Tubular Cell Injury and Inflammatory Response

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
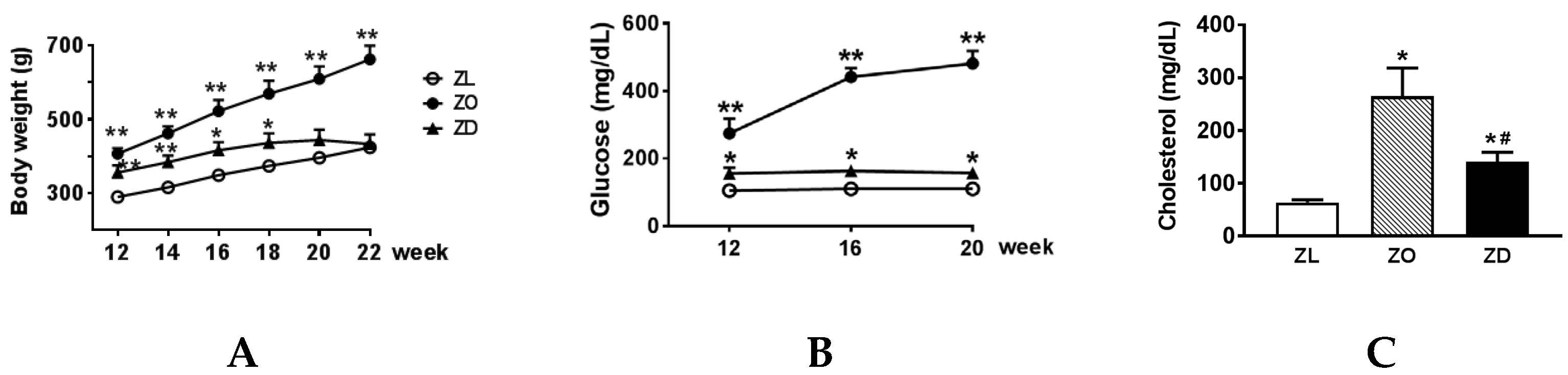
2.1. Metabolic Parameters
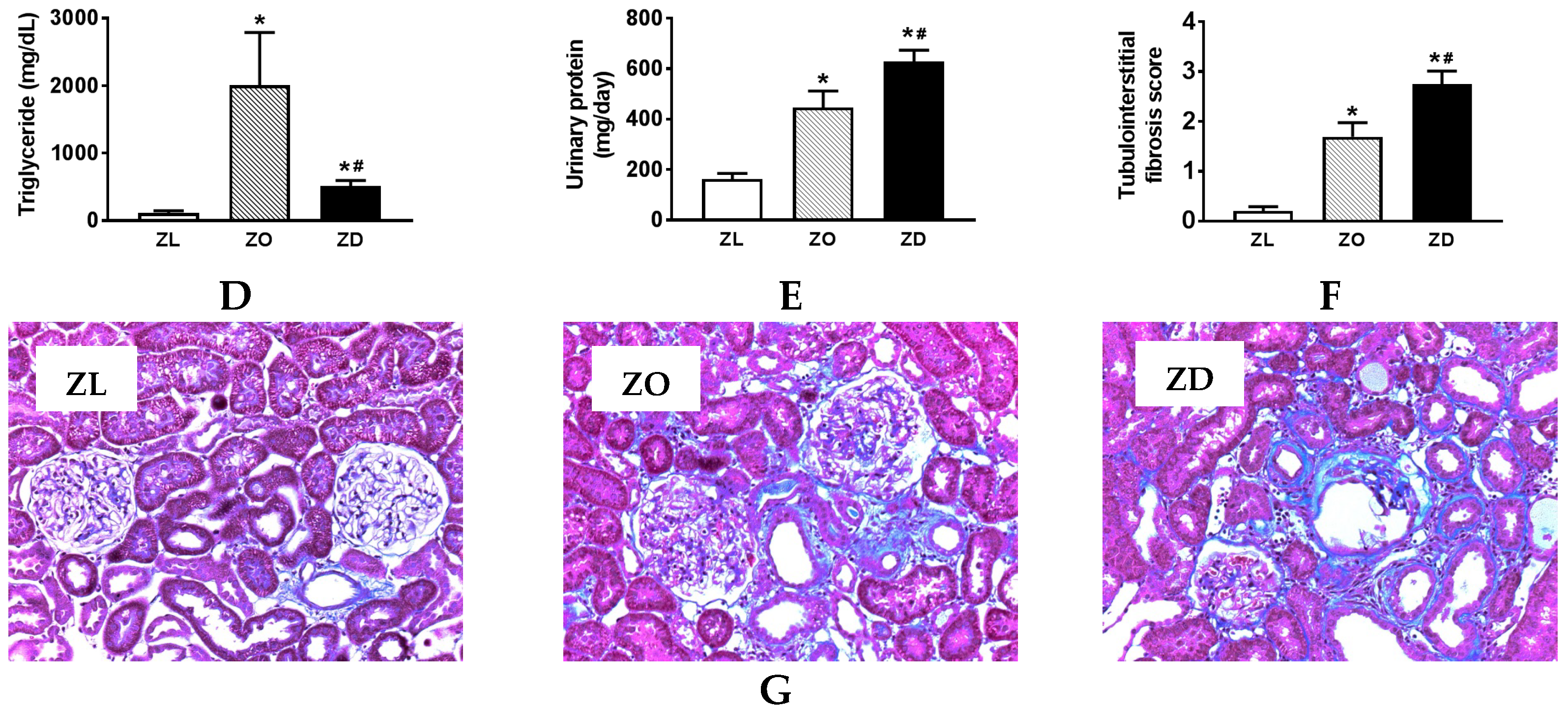
2.2. Kidney Injury in ZO and ZD Rats
2.3. Genes Commonly Impacted in ZO and ZD Kidneys
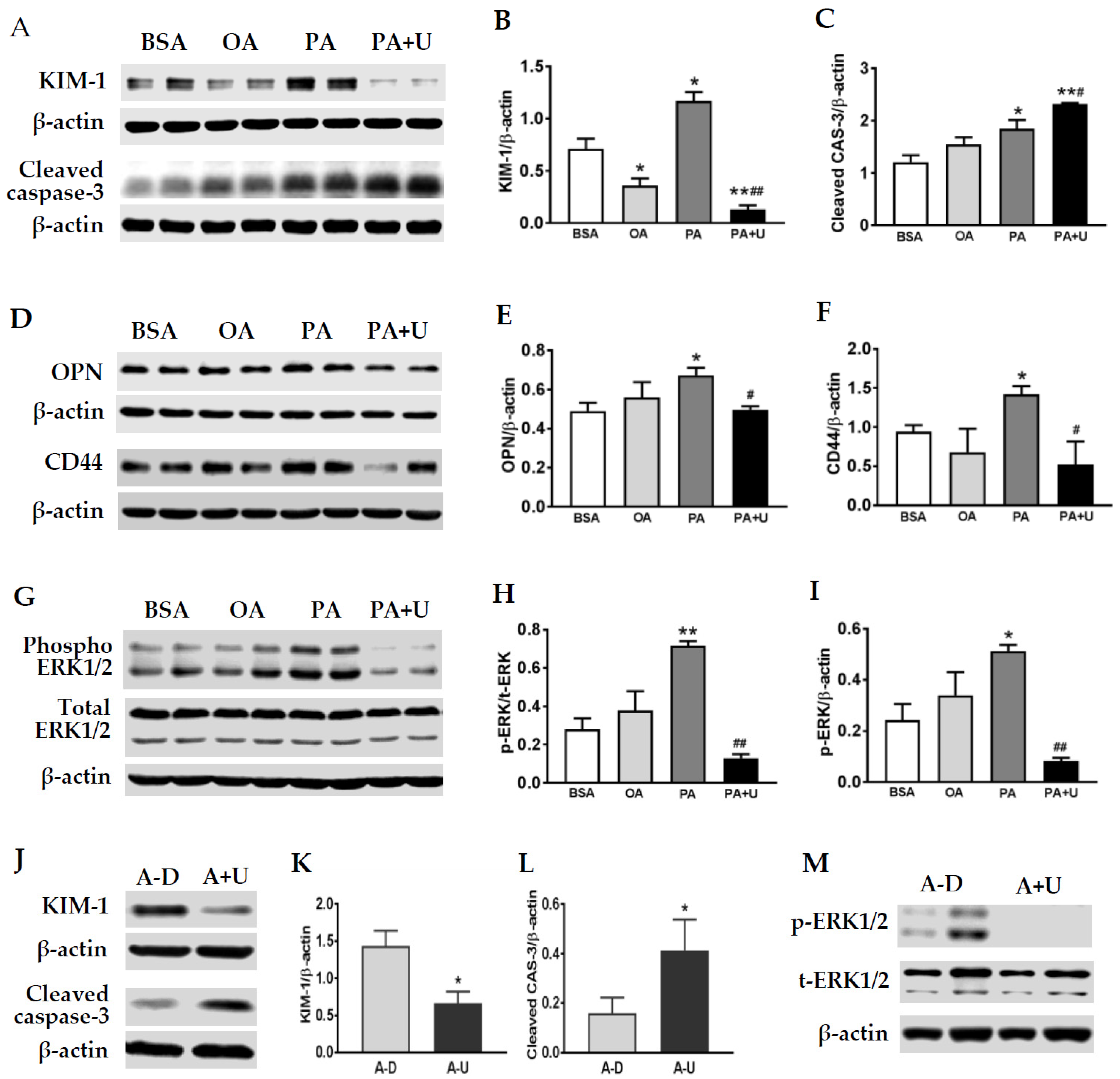
2.4. An Upregulation of KIM-1 Was Associated with Lipid Accumulation and CD44 Induction in Injured Proximal Tubular Cells of ZO and ZD Rats
2.5. Saturated FA Palmitate Increased KIM-1, OPN, and CD44 Expression via the ERK1/2 Signaling Pathway in Primary Rat TECs
2.6. KIM-1 Is Involved in Palmitate-Induced Caspase-3 Activation and Upregulation of OPN and CD44
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. RNA Isolation
4.3. Gene Microarray
4.4. Quantitative Real-Time PCR
4.5. Light Microscopy of Kidney Sections
4.6. Immunostaining
4.7. Culture of Primary Tubular Epithelial Cells and FA Treatment
4.8. RNA Interference
4.9. Western Blot Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Boden, G. Obesity and free fatty acids. Endocrinol. Metab. Clin. N. Am. 2008, 37, 635–636. [Google Scholar] [CrossRef]
- Fagot-Campagna, A.; Balkau, B.; Simon, D.; Warnet, J.M.; Claude, J.R.; Ducimetiere, P.; Eschwege, E. High free fatty acid concentration: An independent risk factor for hypertension in the Paris Prospective Study. Int. J. Epidemiol. 1998, 27, 808–813. [Google Scholar] [CrossRef]
- Reaven, G.M.; Hollenbeck, C.; Jeng, C.Y.; Wu, M.S.; Chen, Y.D. Measurement of plasma glucose, free fatty acid, lactate, and insulin for 24 h in patients with NIDDM. Diabetes 1988, 37, 1020–1024. [Google Scholar] [CrossRef]
- Shulman, G.I. Ectopic fat in insulin resistance, dyslipidemia, and cardiometabolic disease. N. Engl. J. Med. 2014, 371, 1131–1141. [Google Scholar] [CrossRef]
- Kamijo, A.; Kimura, K.; Sugaya, T.; Yamanouchi, M.; Hase, H.; Kaneko, T.; Hirata, Y.; Goto, A.; Fujita, T.; Omata, M. Urinary free fatty acids bound to albumin aggravate tubulointerstitial damage. Kidney Int. 2002, 62, 1628–1637. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Abu Jawdeh, B.G.; Goel, M.; Schilling, W.P.; Parker, M.D.; Puchowicz, M.A.; Yadav, S.P.; Harris, R.C.; El-Meanawy, A.; Hoshi, M.; et al. Lipotoxic disruption of NHE1 interaction with PI(4,5)P2 expedites proximal tubule apoptosis. J. Clin. Investig. 2014, 124, 1057–1068. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Cabral, P.D.; Schilling, W.P.; Schmidt, Z.W.; Uddin, A.N.; Gingras, A.; Madhavan, S.M.; Garvin, J.L.; Schelling, J.R. Kidney Proximal Tubule Lipoapoptosis Is Regulated by Fatty Acid Transporter-2 (FATP2). J. Am. Soc. Nephrol. 2018, 29, 81–91. [Google Scholar] [CrossRef]
- Ichimura, T.; Asseldonk, E.J.; Humphreys, B.D.; Gunaratnam, L.; Duffield, J.S.; Bonventre, J.V. Kidney injury molecule-1 is a phosphatidylserine receptor that confers a phagocytic phenotype on epithelial cells. J. Clin. Investig. 2008, 118, 1657–1668. [Google Scholar] [CrossRef]
- Kuehn, E.W.; Park, K.M.; Somlo, S.; Bonventre, J.V. Kidney injury molecule-1 expression in murine polycystic kidney disease. Am. J. Physiol. Renal. Physiol. 2002, 283, F1326–F1336. [Google Scholar] [CrossRef] [Green Version]
- Van Timmeren, M.M.; Bakker, S.J.; Vaidya, V.S.; Bailly, V.; Schuurs, T.A.; Damman, J.; Stegeman, C.A.; Bonventre, J.V.; Van, G.H. Tubular kidney injury molecule-1 in protein-overload nephropathy. Am. J. Physiol. Renal. Physiol. 2006, 291, F456–F464. [Google Scholar] [CrossRef]
- Li, L.; Emmett, N.; Mann, D.; Zhao, X. Fenofibrate attenuates tubulointerstitial fibrosis and inflammation through suppression of nuclear factor-kappaB and transforming growth factor-beta1/Smad3 in diabetic nephropathy. Exp. Biol. Med. 2010, 235, 383–391. [Google Scholar] [CrossRef]
- Zhang, Y.; George, J.; Li, Y.; Olufade, R.; Zhao, X. Matrix metalloproteinase-9 expression is enhanced in renal parietal epithelial cells of zucker diabetic Fatty rats and is induced by albumin in in vitro primary parietal cell culture. PLoS ONE 2015, 10, e0123276. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, Y.; Li, L.; Mann, D.; Imig, J.D.; Emmett, N.; Gibbons, G.; Jin, L.M. Glomerular expression of kidney injury molecule-1 and podocytopenia in diabetic glomerulopathy. Am. J. Nephrol. 2011, 34, 268–280. [Google Scholar] [CrossRef]
- Nowak, N.; Skupien, J.; Niewczas, M.A.; Yamanouchi, M.; Major, M.; Croall, S.; Smiles, A.; Warram, J.H.; Bonventre, J.V.; Krolewski, A.S. Increased plasma kidney injury molecule-1 suggests early progressive renal decline in non-proteinuric patients with type 1 diabetes. Kidney Int. 2016, 89, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Nowak, N.; Skupien, J.; Smiles, A.M.; Yamanouchi, M.; Niewczas, M.A.; Galecki, A.T.; Duffin, K.L.; Breyer, M.D.; Pullen, N.; Bonventre, J.V.; et al. Markers of early progressive renal decline in type 2 diabetes suggest different implications for etiological studies and prognostic tests development. Kidney Int. 2018, 93, 1198–1206. [Google Scholar] [CrossRef]
- Zylka, A.; Dumnicka, P.; Kusnierz-Cabala, B.; Gala-Bladzinska, A.; Ceranowicz, P.; Kucharz, J.; Zabek-Adamska, A.; Maziarz, B.; Drozdz, R.; Kuzniewski, M. Markers of Glomerular and Tubular Damage in the Early Stage of Kidney Disease in Type 2 Diabetic Patients. Mediat. Inflamm. 2018, 2018, 7659243. [Google Scholar] [CrossRef]
- Panduru, N.M.; Sandholm, N.; Forsblom, C.; Saraheimo, M.; Dahlstrom, E.H.; Thorn, L.M.; Gordin, D.; Tolonen, N.; Waden, J.; Harjutsalo, V.; et al. Kidney injury molecule-1 and the loss of kidney function in diabetic nephropathy: A likely causal link in patients with type 1 diabetes. Diabetes Care 2015, 38, 1130–1137. [Google Scholar] [CrossRef]
- Han, W.K.; Bailly, V.; Abichandani, R.; Thadhani, R.; Bonventre, J.V. Kidney Injury Molecule-1 (KIM-1): A novel biomarker for human renal proximal tubule injury. Kidney Int. 2002, 62, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Han, W.K.; Waikar, S.S.; Johnson, A.; Betensky, R.A.; Dent, C.L.; Devarajan, P.; Bonventre, J.V. Urinary biomarkers in the early diagnosis of acute kidney injury. Kidney Int. 2008, 73, 863–869. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, B.D.; Xu, F.; Sabbisetti, V.; Grgic, I.; Movahedi, N.S.; Wang, N.; Chen, G.; Xiao, S.; Patel, D.; Henderson, J.M.; et al. Chronic epithelial kidney injury molecule-1 expression causes murine kidney fibrosis. J. Clin. Investig. 2013, 123, 4023–4035. [Google Scholar] [CrossRef] [Green Version]
- Ichimura, T.; Bonventre, J.V.; Bailly, V.; Wei, H.; Hession, C.A.; Cate, R.L.; Sanicola, M. Kidney injury molecule-1 (KIM-1), a putative epithelial cell adhesion molecule containing a novel immunoglobulin domain, is up-regulated in renal cells after injury. J. Biol. Chem. 1998, 273, 4135–4142. [Google Scholar] [CrossRef]
- Wagoner, M.P.; Yang, Y.; McDuffie, J.E.; Klapczynski, M.; Buck, W.; Cheatham, L.; Eisinger, D.; Sace, F.; Lynch, K.M.; Sonee, M.; et al. Evaluation of Temporal Changes in Urine-based Metabolomic and Kidney Injury Markers to Detect Compound Induced Acute Kidney Tubular Toxicity in Beagle Dogs. Curr. Top. Med. Chem. 2017, 17, 2767–2780. [Google Scholar] [CrossRef]
- Chen, X.; Cobbs, A.; George, J.; Chima, A.; Tuyishime, F.; Zhao, X. Endocytosis of Albumin Induces Matrix Metalloproteinase-9 by Activating the ERK Signaling Pathway in Renal Tubule Epithelial Cells. Int. J. Mol. Sci. 2017, 18, 1758. [Google Scholar] [CrossRef]
- Cobbs, A.; Ballou, K.; Chen, X.; George, J.; Zhao, X. Saturated fatty acids bound to albumin enhance osteopontin expression and cleavage in renal proximal tubular cells. Int. J. Physiol. Pathophysiol. Pharmacol. 2018, 10, 29–38. [Google Scholar]
- Van Timmeren, M.M.; Gross, M.L.; Hanke, W.; Klok, P.A.; Van, G.H.; Stegeman, C.A.; Bakker, S.J. Oleic acid loading does not add to the nephrotoxic effect of albumin in an amphibian and chronic rat model of kidney injury. Nephrol. Dial. Transplant. 2008, 23, 3814–3823. [Google Scholar] [CrossRef] [Green Version]
- Collier, J.B.; Schnellmann, R.G. Extracellular Signal-Regulated Kinase 1/2 Regulates Mouse Kidney Injury Molecule-1 Expression Physiologically and Following Ischemic and Septic Renal Injury. J. Pharmacol. Exp. Ther. 2017, 363, 419–427. [Google Scholar] [CrossRef]
- Arici, M.; Chana, R.; Lewington, A.; Brown, J.; Brunskill, N.J. Stimulation of proximal tubular cell apoptosis by albumin-bound fatty acids mediated by peroxisome proliferator activated receptor-gamma. J. Am. Soc. Nephrol. 2003, 14, 17–27. [Google Scholar] [CrossRef]
- Guder, W.G.; Wagner, S.; Wirthensohn, G. Metabolic fuels along the nephron: Pathways and intracellular mechanisms of interaction. Kidney Int. 1986, 29, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Nieth, H.; Schollmeyer, P. Substrate-utilization of the human kidney. Nature 1966, 209, 1244–1245. [Google Scholar] [CrossRef]
- Proctor, G.; Jiang, T.; Iwahashi, M.; Wang, Z.; Li, J.; Levi, M. Regulation of renal fatty acid and cholesterol metabolism, inflammation, and fibrosis in Akita and OVE26 mice with type 1 diabetes. Diabetes 2006, 55, 2502–2509. [Google Scholar] [CrossRef]
- Sun, L.; Halaihel, N.; Zhang, W.; Rogers, T.; Levi, M. Role of sterol regulatory element-binding protein 1 in regulation of renal lipid metabolism and glomerulosclerosis in diabetes mellitus. J. Biol. Chem. 2002, 277, 18919–18927. [Google Scholar] [CrossRef]
- Kang, H.M.; Ahn, S.H.; Choi, P.; Ko, Y.A.; Han, S.H.; Chinga, F.; Park, A.S.; Tao, J.; Sharma, K.; Pullman, J.; et al. Defective fatty acid oxidation in renal tubular epithelial cells has a key role in kidney fibrosis development. Nat. Med. 2015, 21, 37–46. [Google Scholar] [CrossRef]
- Moorhead, J.F.; Chan, M.K.; El-Nahas, M.; Varghese, Z. Lipid nephrotoxicity in chronic progressive glomerular and tubulo-interstitial disease. Lancet 1982, 320, 1309–1311. [Google Scholar] [CrossRef]
- Roeder, S.S.; Barnes, T.J.; Lee, J.S.; Kato, I.; Eng, D.G.; Kaverina, N.V.; Sunseri, M.W.; Daniel, C.; Amann, K.; Pippin, J.W.; et al. Activated ERK1/2 increases CD44 in glomerular parietal epithelial cells leading to matrix expansion. Kidney Int. 2017, 91, 896–913. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, X.; Chima, A.; Zhang, Y.; George, J.; Cobbs, A.; Emmett, N. Albumin induces CD44 expression in glomerular parietal epithelial cells by activating extracellular signal-regulated kinase 2018, 1/2 pathway. J. Cell. Physiol. 2018, 234, 7224–7235. [Google Scholar] [CrossRef]
- Sauvant, C.; Holzinger, H.; Gekle, M. Proximal tubular toxicity of ochratoxin A is amplified by simultaneous inhibition of the extracellular signal-regulated kinases 1/2. J. Pharmacol. Exp. Ther. 2005, 313, 234–241. [Google Scholar] [CrossRef]
- Chang, H.K.; Kim, D.S.; Chae, J.J.; Kim, M.; Myong, J.P.; Lee, K.H.; Lee, M.W.; Park, T.C. Inhibition of ERK activity enhances the cytotoxic effect of peroxisome proliferator-activated receptor gamma (PPARgamma) agonists in HeLa cells. Biochem. Biophys. Res. Commun. 2017, 482, 843–848. [Google Scholar] [CrossRef]
- Yang, L.; Brooks, C.R.; Xiao, S.; Sabbisetti, V.; Yeung, M.Y.; Hsiao, L.L.; Ichimura, T.; Kuchroo, V.; Bonventre, J.V. KIM-1-mediated phagocytosis reduces acute injury to the kidney. J. Clin. Investig. 2015, 125, 1620–1636. [Google Scholar] [CrossRef]
- Tian, L.; Shao, X.; Xie, Y.; Wang, Q.; Che, X.; Zhang, M.; Xu, W.; Xu, Y.; Mou, S.; Ni, Z. Kidney Injury Molecule-1 is Elevated in Nephropathy and Mediates Macrophage Activation via the Mapk Signalling Pathway. Cell. Physiol. Biochem. 2017, 41, 769–783. [Google Scholar] [CrossRef]
- Yin, W.; Naini, S.M.; Chen, G.; Hentschel, D.M.; Humphreys, B.D.; Bonventre, J.V. Mammalian Target of Rapamycin Mediates Kidney Injury Molecule 1-Dependent Tubule Injury in a Surrogate Model. J. Am. Soc. Nephrol. 2016, 27, 1943–1957. [Google Scholar] [CrossRef]
- He, L.; Weber, K.J.; Diwan, A.; Schilling, J.D. Inhibition of mTOR reduces lipotoxic cell death in primary macrophages through an autophagy-independent mechanism. J. Leukoc. Biol. 2016, 100, 1113–1124. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Fold-Change | GenBank ID | |
|---|---|---|---|
| ZO | ZD | ||
| Interleukin 24 | 32.02 | 19.73 | NM_133311 |
| Alpha-2-macroglobulin | 11.92 | 4.60 | NM_012488 |
| Adrenergic receptor, alpha 1d | 10.61 | 7.60 | NM_024483 |
| Early growth response 2 | 6.31 | 3.41 | NM_053633 |
| Kininogen 1 | 4.54 | 6.31 | NM_012696 |
| Activating transcription factor 3 | 3.68 | 2.16 | NM_012912 |
| Fibronectin 1 | 3.47 | 3.15 | NM_019143 |
| Poliovirus receptor | 2.86 | 2.56 | NM_017076 |
| Chemokine (C-X-C motif) ligand 1 | 2.73 | 2.81 | NM_030845 |
| Chemokine (C-X-C motif) ligand 2 | 2.62 | 4.95 | NM_053647 |
| Myelocytomatosis viral oncogene homolog (avian) | 2.61 | 2.29 | NM_012603 |
| Fos-like antigen 1 | 2.46 | 2.45 | NM_012953 |
| Nuclear protein 1 | 2.39 | 2.13 | NM_053611 |
| Lysyl oxidase | 2.17 | 2.10 | NM_017061 |
| Parathyroid hormone-like peptide | 2.16 | 2.04 | NM_012636 |
| Gene | Fold-Change | GenBank ID | |
|---|---|---|---|
| ZO | ZD | ||
| Tubular Necrosis Related | |||
| Kidney injury molecule 1 (HAVCR1) | 21.30 | 20.67 | NM_173149 |
| Secreted phosphoprotein 1 (SPP1, osteopontin) | 6.34 | 7.82 | NM_012881 |
| Fibrosis and Organ Damage Related | |||
| LIM/homeobox protein Lhx2 (Homeobox protein LH-2) | 4.98 | 2.69 | ENSRNOT00000014452 |
| Matrix metallopeptidase 12 (Mmp12) | 4.55 | 4.38 | NM_053963 |
| Troponin T2, cardiac (Tnnt2) | 3.86 | 3.01 | NM_012676 |
| Activating transcription factor 3 (Atf3) | 3.68 | 2.16 | NM_012912 |
| Serine (or cysteine) peptidase inhibitor, clade E, member 1 (Serpine1) | 3.65 | 2.86 | NM_012620 |
| Adenosine A1 receptor | 3.35 | 3.16 | ENSRNOT00000004602 |
| CD44 antigen (CD44) | 3.10 | 2.58 | NM_012924 |
| Interleukin 1 receptor antagonist (Il1rn) | 2.57 | 2.27 | NM_022194 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Chen, X.; Zhang, Y.; George, J.; Cobbs, A.; Wang, G.; Li, L.; Emmett, N. Kidney Injury Molecule-1 Is Upregulated in Renal Lipotoxicity and Mediates Palmitate-Induced Tubular Cell Injury and Inflammatory Response. Int. J. Mol. Sci. 2019, 20, 3406. https://doi.org/10.3390/ijms20143406
Zhao X, Chen X, Zhang Y, George J, Cobbs A, Wang G, Li L, Emmett N. Kidney Injury Molecule-1 Is Upregulated in Renal Lipotoxicity and Mediates Palmitate-Induced Tubular Cell Injury and Inflammatory Response. International Journal of Molecular Sciences. 2019; 20(14):3406. https://doi.org/10.3390/ijms20143406
Chicago/Turabian StyleZhao, Xueying, Xiaoming Chen, Yuanyuan Zhang, Jasmine George, Alyssa Cobbs, Guoshen Wang, Lingyun Li, and Nerimiah Emmett. 2019. "Kidney Injury Molecule-1 Is Upregulated in Renal Lipotoxicity and Mediates Palmitate-Induced Tubular Cell Injury and Inflammatory Response" International Journal of Molecular Sciences 20, no. 14: 3406. https://doi.org/10.3390/ijms20143406
APA StyleZhao, X., Chen, X., Zhang, Y., George, J., Cobbs, A., Wang, G., Li, L., & Emmett, N. (2019). Kidney Injury Molecule-1 Is Upregulated in Renal Lipotoxicity and Mediates Palmitate-Induced Tubular Cell Injury and Inflammatory Response. International Journal of Molecular Sciences, 20(14), 3406. https://doi.org/10.3390/ijms20143406