Sphingolipid-Transporting Proteins as Cancer Therapeutic Targets


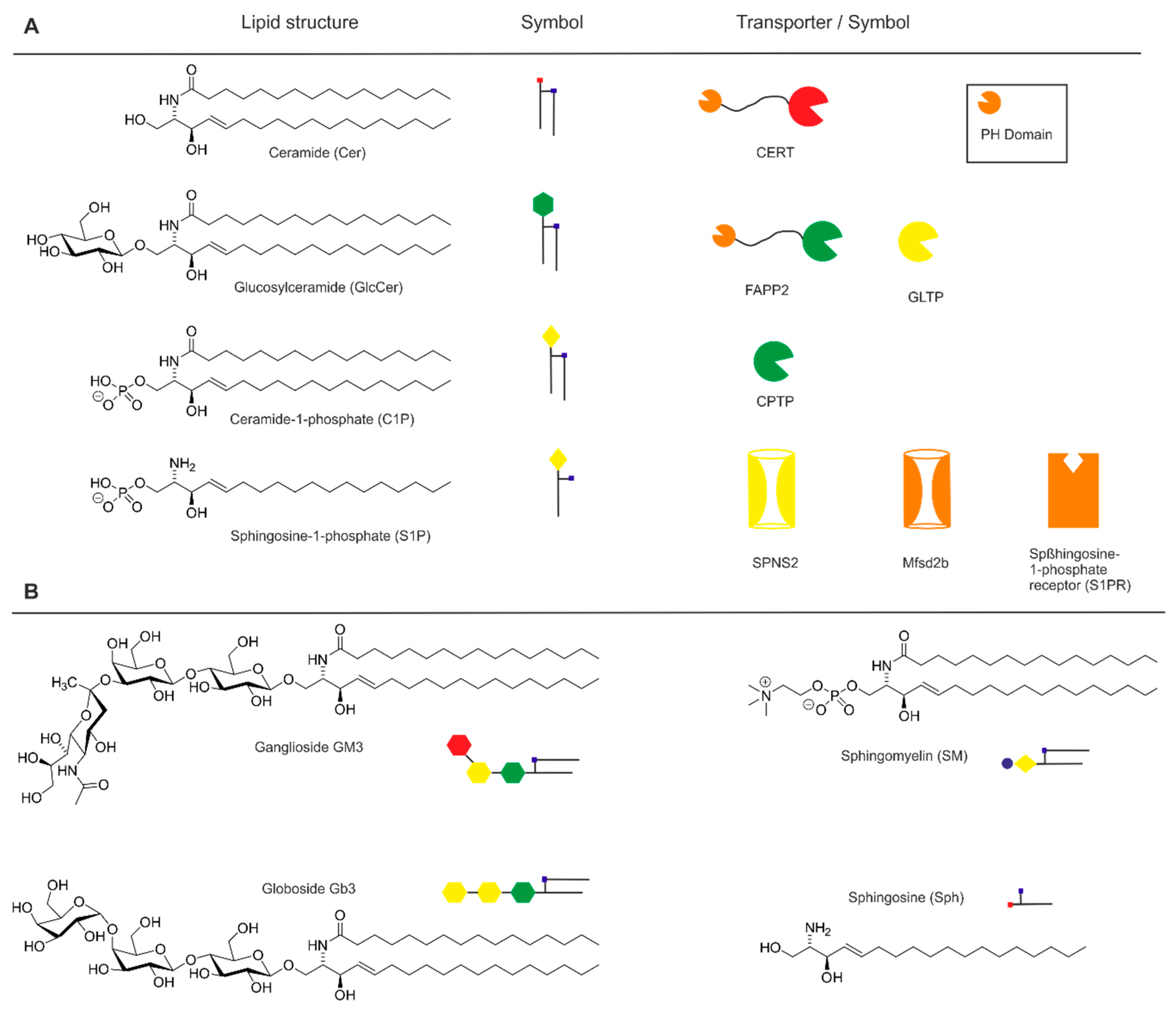{kind=link}
{kind=link}
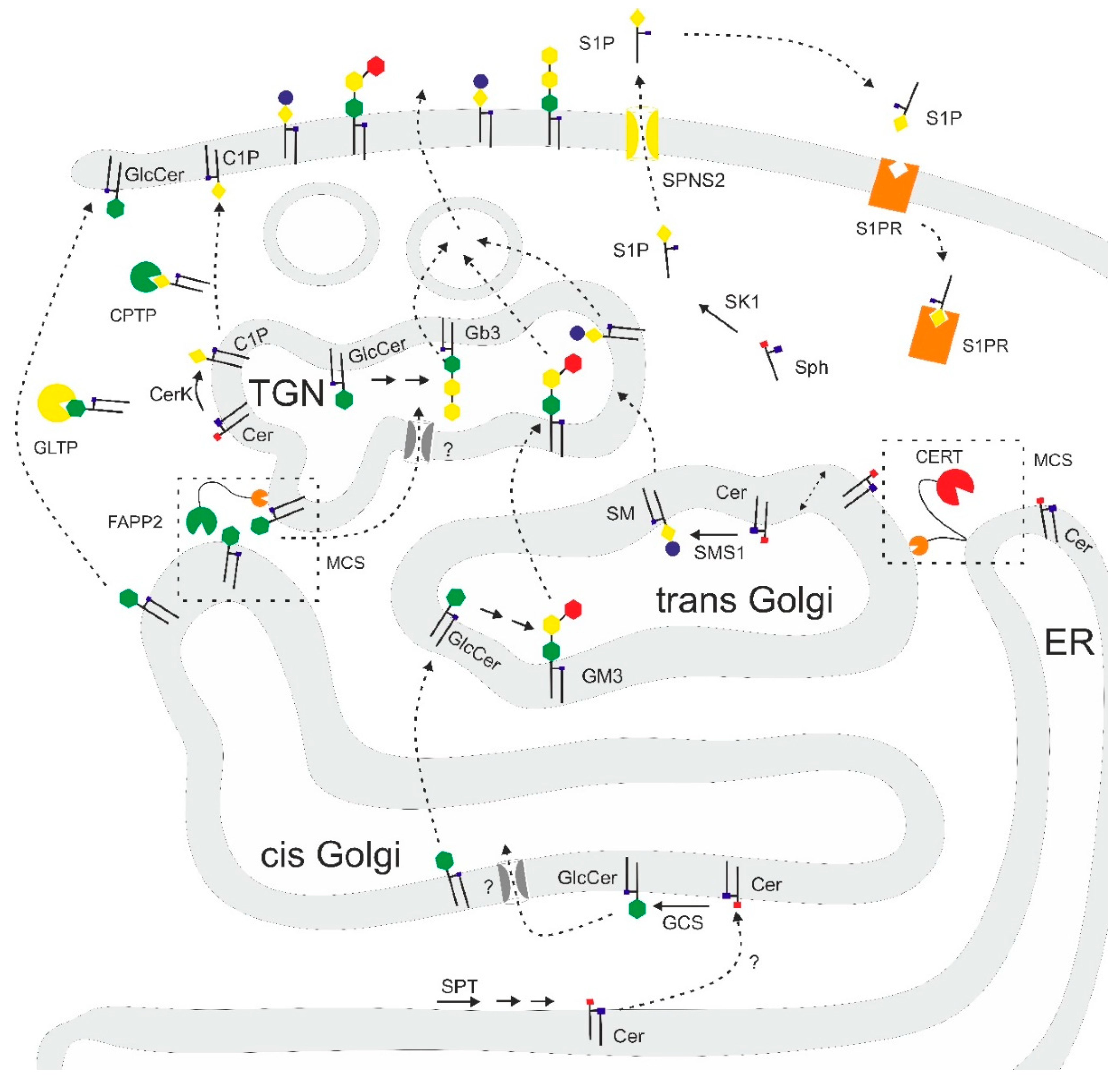{kind=link}
Abstract
:1. Introduction
2. Ceramide Transfer Protein (CERT)
2.1. CERT and Cancer
2.2. CERT Inhibitors
3. FAPP2
3.1. FAPP2 in Cancer
3.2. FAPP2 Inhibitors
4. Glycolipid Transfer Protein (GLTP)
4.1. GLTP and Cancer
4.2. GLTP Inhibitors
5. Ceramide-1-Phosphate Transport Protein (CPTP)
5.1. CPTP and Cancer
5.2. CPTP Inhibitors
6. Sphingosine-1-Phosphate Transporters
7. Spinster Homolog 2 (SPNS2)
7.1. SPNS2 and Cancer
7.2. SPNS2 Inhibitors
8. Mfsd2b
8.1. Mfsd2b
8.2. Mfsd2b Inhibitors
9. Conclusions
Acknowledgments
Conflicts of Interest
References
- Wong, L.H.; Copic, A.; Levine, T.P. Advances on the Transfer of Lipids by Lipid Transfer Proteins. Trends Biochem. Sci. 2017, 42, 516–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, L.H.; Gatta, A.T.; Levine, T.P. Lipid transfer proteins: The lipid commute via shuttles, bridges and tubes. Nat. Rev. Mol. Cell Biol. 2019, 20, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Füllekrug, J.; Simons, K. Lipid Rafts and Apical Membrane Traffic. Ann. N. Y. Acad. Sci. 2004, 1014, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golub, T.; Wacha, S.; Caroni, P. Spatial and temporal control of signaling through lipid rafts. Curr. Opin. Neurobiol. 2004, 14, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Kraft, M.L. Plasma membrane organization and function: Moving past lipid rafts. Mol. Biol. Cell 2013, 24, 2765–2768. [Google Scholar] [CrossRef] [PubMed]
- Sevcsik, E.; Schutz, G.J. With or without rafts? Alternative views on cell membranes. Bioessays 2016, 38, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Holthuis, J.C.M.; Levine, T.P.; Levine, T. Lipid traffic: Floppy drives and a superhighway. Nat. Rev. Mol. Cell Biol. 2005, 6, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Ogushi, F.; Ishitsuka, R.; Kobayashi, T.; Sugita, Y. Rapid flip-flop motions of diacylglycerol and ceramide in phospholipid bilayers. Chem. Phys. Lett. 2012, 522, 96–102. [Google Scholar] [CrossRef]
- Airenne, T.T.; Kidron, H.; Nymalm, Y.; Nylund, M.; West, G.; Mattjus, P.; Salminen, T.A. Structural Evidence for Adaptive Ligand Binding of Glycolipid Transfer Protein. J. Mol. Biol. 2006, 355, 224–236. [Google Scholar] [CrossRef]
- Hanada, K. Lipid transfer proteins rectify inter-organelle flux and accurately deliver lipids at membrane contact sites. J. Lipid Res. 2018, 59, 1341–1366. [Google Scholar] [CrossRef] [Green Version]
- García-Pacios, M.; Collado, M.I.; Busto, J.V.; Sot, J.; Alonso, A.; Arrondo, J.-L.R.; Goñi, F.M. Sphingosine-1-Phosphate as an Amphipathic Metabolite: Its Properties in Aqueous and Membrane Environments. Biophys. J. 2009, 97, 1398–1407. [Google Scholar] [CrossRef] [Green Version]
- Hanada, K.; Kumagai, K.; Yasuda, S.; Miura, Y.; Kawano, M.; Fukasawa, M.; Nishijima, M. Molecular machinery for non-vesicular trafficking of ceramide. Nature 2003, 426, 803–809. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Lahiri, S.; Futerman, A.H.; Futerman, A. The metabolism and function of sphingolipids and glycosphingolipids. Cell. Mol. Life Sci. 2007, 64, 2270–2284. [Google Scholar] [CrossRef]
- Smyth, M.J.; Obeid, L.M.; Hannunf, Y.A. Ceramide: A Novel Lipid Mediator of Apoptosis. Adv. Pharmacol. 1997, 41, 133–154. [Google Scholar] [PubMed]
- Bartke, N.; Hannun, Y.A. Bioactive sphingolipids: Metabolism and function. J. Lipid Res. 2009, 50, 17. [Google Scholar] [CrossRef]
- Young, M.M.; Kester, M.; Wang, H.G. Sphingolipids: Regulators of crosstalk between apoptosis and autophagy. J. Lipid. Res. 2013, 54, 5–19. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Tumor suppressive functions of ceramide: Evidence and mechanisms. Apoptosis 2015, 20, 689–711. [Google Scholar] [CrossRef]
- Raya, A.; Revert, F.; Navarro, S.; Saus, J. Characterization of a Novel Type of Serine/Threonine Kinase That Specifically Phosphorylates the Human Goodpasture Antigen. J. Biol. Chem. 1999, 274, 12642–12649. [Google Scholar] [CrossRef] [Green Version]
- Revert, F.; Ventura, I.; Martinez-Martinez, P.; Granero-Moltó, F.; Revert-Ros, F.; Macias, J.; Saus, J. Goodpasture Antigen-binding Protein Is a Soluble Exportable Protein That Interacts with Type IV Collagen. J. Biol. Chem. 2008, 283, 30246–30255. [Google Scholar] [CrossRef]
- Mencarelli, C.; Losen, M.; Hammels, C.; De Vry, J.; Hesselink, M.K.C.; Steinbusch, H.W.M.; De Baets, M.H.; Martínez-Martínez, P. The ceramide transporter and the Goodpasture antigen binding protein: One protein—One function? J. Neurochem. 2010, 113, 1369–1386. [Google Scholar] [CrossRef] [PubMed]
- Kawano, M.; Kumagai, K.; Nishijima, M.; Hanada, K. Efficient Trafficking of Ceramide from the Endoplasmic Reticulum to the Golgi Apparatus Requires a VAMP-associated Protein-interacting FFAT Motif of CERT. J. Biol. Chem. 2006, 281, 30279–30288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, S.; Matsui, H.; Kawano, M.; Kumagai, K.; Tomishige, N.; Hanada, K.; Echigo, S.; Tamura, S.; Kobayashi, T. Protein phosphatase 2Cepsilon is an endoplasmic reticulum integral membrane protein that dephosphorylates the ceramide transport protein CERT to enhance its association with organelle membranes. J. Biol. Chem. 2008, 283, 6584–6593. [Google Scholar] [CrossRef] [PubMed]
- Tomishige, N.; Kumagai, K.; Kusuda, J.; Nishijima, M.; Hanada, K. Casein kinase I{gamma}2 down-regulates trafficking of ceramide in the synthesis of sphingomyelin. Mol. Biol. Cell 2009, 20, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.P.; Scheffer, L.; Srideshikan, S.M.; Parthibane, V.; Kosakowska-Cholody, T.; Masood, M.A.; Nagashima, K.; Gudla, P.; Lockett, S.; Acharya, U.; et al. Ceramide Transfer Protein Deficiency Compromises Organelle Function and Leads to Senescence in Primary Cells. PLoS ONE 2014, 9, e92142. [Google Scholar] [CrossRef] [PubMed]
- Swanton, C.; Marani, M.; Pardo, O.; Warne, P.H.; Kelly, G.; Sahai, E.; Elustondo, F.; Chang, J.; Temple, J.; Ahmed, A.A.; et al. Regulators of Mitotic Arrest and Ceramide Metabolism Are Determinants of Sensitivity to Paclitaxel and Other Chemotherapeutic Drugs. Cancer Cell 2007, 11, 498–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juul, N.; Szallasi, Z.; Eklund, A.C.; Li, Q.; A Burrell, R.; Gerlinger, M.; Valero, V.; Andreopoulou, E.; Esteva, F.J.; Symmans, W.F.; et al. Assessment of an RNA interference screen-derived mitotic and ceramide pathway metagene as a predictor of response to neoadjuvant paclitaxel for primary triple-negative breast cancer: A retrospective analysis of five clinical trials. Lancet Oncol. 2010, 11, 358–365. [Google Scholar] [CrossRef]
- Lee, A.J.; Roylance, R.; Sander, J.; Gorman, P.; Endesfelder, D.; Kschischo, M.; Jones, N.P.; East, P.; Nicke, B.; Spassieva, S.; et al. CERT depletion predicts chemotherapy benefit and mediates cytotoxic and polyploid-specific cancer cell death through autophagy induction. J. Pathol. 2012, 226, 482–494. [Google Scholar] [CrossRef]
- Heering, J.; Weis, N.; Holeiter, M.; Neugart, F.; Staebler, A.; Fehm, T.N.; Bischoff, A.; Schiller, J.; Duss, S.; Schmid, S.; et al. Loss of the Ceramide Transfer Protein Augments EGF Receptor Signaling in Breast Cancer. Cancer Res. 2012, 72, 2855–2866. [Google Scholar] [CrossRef]
- Berkeš, D.; Daïch, A.; Santos, C.; Ballereau, S.; Génisson, Y. Chemistry and Biology of HPAs: A Family of Ceramide Trafficking Inhibitors. Chem. A Eur. J. 2016, 22, 17514–17525. [Google Scholar] [CrossRef]
- Yasuda, S.; Kitagawa, H.; Ueno, M.; Ishitani, H.; Fukasawa, M.; Nishijima, M.; Kobayashi, S.; Hanada, K. A Novel Inhibitor of Ceramide Trafficking from the Endoplasmic Reticulum to the Site of Sphingomyelin Synthesis. J. Biol. Chem. 2001, 276, 43994–44002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saied, E.M.; Diederich, S.; Arenz, C. Facile Synthesis of the CERT Inhibitor HPA-12 and Some Novel Derivatives. Chem. Asian J. 2014, 9, 2092–2094. [Google Scholar] [CrossRef] [PubMed]
- Nakao, N.; Ueno, M.; Sakai, S.; Egawa, D.; Hanzawa, H.; Kawasaki, S.; Kumagai, K.; Suzuki, M.; Kobayashi, S.; Hanada, K. Natural ligand-nonmimetic inhibitors of the lipid-transfer protein CERT. Commun. Chem. 2019, 2, 20. [Google Scholar] [CrossRef]
- D’Angelo, G.; Polishchuk, E.; Di Tullio, G.; Santoro, M.; Di Campli, A.; Godi, A.; West, G.; Bielawski, J.; Chuang, C.-C.; Van Der Spoel, A.C.; et al. Glycosphingolipid synthesis requires FAPP2 transfer of glucosylceramide. Nature 2007, 449, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Lizarralde, B.; Gao, Y.-G.; Popov, A.N.; Samygina, V.R.; Zhai, X.; Mishra, S.K.; Boldyrev, I.A.; Molotkovsky, J.G.; Simanshu, D.K.; Patel, D.J.; et al. Structural analyses of 4-phosphate adaptor protein 2 yield mechanistic insights into sphingolipid recognition by the glycolipid transfer protein family. J. Biol. Chem. 2018, 293, 16709–16723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Angelo, G.; Uemura, T.; Chuang, C.-C.; Polishchuk, E.; Santoro, M.; Ohvo-Rekilä, H.; Sato, T.; Di Tullio, G.; Varriale, A.; D’Auria, S.; et al. Vesicular and non-vesicular transport feed distinct glycosylation pathways in the Golgi. Nature 2013, 501, 116–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Angelo, G.; Rega, L.R.; De Matteis, M.A. Connecting vesicular transport with lipid synthesis: FAPP2. Biochim. Biophys. Acta 2012, 1821, 1089–1095. [Google Scholar] [CrossRef] [Green Version]
- Chalat, M.; Menon, I.; Turan, Z.; Menon, A.K. Reconstitution of glucosylceramide flip-flop across endoplasmic reticulum: Implications for mechanism of glycosphingolipid biosynthesis. J. Biol. Chem. 2012, 287, 15523–15532. [Google Scholar] [CrossRef]
- Khan, I.; Katikaneni, D.S.; Han, Q.; Sanchez-Felipe, L.; Hanada, K.; Ambrose, R.L.; MacKenzie, J.M.; Konan, K.V.; Williams, B. Modulation of Hepatitis C Virus Genome Replication by Glycosphingolipids and Four-Phosphate Adaptor Protein 2. J. Virol. 2014, 88, 12276–12295. [Google Scholar] [CrossRef] [Green Version]
- Tritz, R.; Hickey, M.J.; Lin, A.H.; Hadwiger, P.; Sah, D.W.; Neuwelt, E.A.; Mueller, B.M.; Kruse, C.A. FAPP2 Gene Downregulation Increases Tumor Cell Sensitivity to Fas-Induced Apoptosis. Biochem. Biophys. Res. Commun. 2009, 383, 167–171. [Google Scholar] [CrossRef]
- Chen, J.; Li, L.; Zhou, Z.; Yu, S.; Li, Y.; Gao, Y. FAPP2 promotes tumor cell growth in human colon cancer through activation of Wnt signaling. Exp. Cell Res. 2019, 374, 12–18. [Google Scholar] [CrossRef]
- La Montagna, R.; De Matteis, M.A.; D’angelo, G. Inhibitors of fapp2 and uses thereof. U.S. Patent Application 14/907, 172, 1 September 2016. [Google Scholar]
- Arenz, C. Recent advances and novel treatments for sphingolipidoses. Future Med. Chem. 2017, 9, 1685–1698. [Google Scholar] [CrossRef]
- Zhai, X.; Malakhova, M.L.; Pike, H.M.; Benson, L.M.; Bergen, H.R., 3rd; Sugar, I.P.; Malinina, L.; Patel, D.J.; Brown, R.E. Glycolipid acquisition by human glycolipid transfer protein dramatically alters intrinsic tryptophan fluorescence: Insights into glycolipid binding affinity. J. Biol. Chem. 2009, 284, 13620–13628. [Google Scholar] [CrossRef]
- Malinina, L.; Simanshu, D.K.; Zhai, X.; Samygina, V.R.; Kamlekar, R.; Kenoth, R.; Ochoa-Lizarralde, B.; Malakhova, M.L.; Molotkovsky, J.G.; Patel, D.J.; et al. Sphingolipid transfer proteins defined by the GLTP-fold. Q. Rev. Biophys. 2015, 48, 281–322. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.E.; Mattjus, P. Glycolipid transfer proteins. Biochim. Biophy. Acta 2007, 1771, 746–760. [Google Scholar] [CrossRef] [Green Version]
- Malinina, L.; Malakhova, M.L.; Teplov, A.; Brown, R.E.; Patel, D.J. Structural basis for glycosphingolipid transfer specificity. Nature 2004, 430, 1048–1053. [Google Scholar] [CrossRef] [Green Version]
- Rao, C.S.; Lin, X.; Pike, H.M.; Molotkovsky, J.G.; Brown, R.E. Glycolipid transfer protein mediated transfer of glycosphingolipids between membranes: A model for action based on kinetic and thermodynamic analyses. Biochemistry 2004, 43, 13805–13815. [Google Scholar] [CrossRef]
- Rao, C.S.; Chung, T.; Pike, H.M.; Brown, R.E. Glycolipid Transfer Protein Interaction with Bilayer Vesicles: Modulation by Changing Lipid Composition. Biophys. J. 2005, 89, 4017–4028. [Google Scholar] [CrossRef] [Green Version]
- Kjellberg, M.A.; Backman, A.P.E.; Ohvo-Rekilä, H.; Mattjus, P. Alternation in the Glycolipid Transfer Protein Expression Causes Changes in the Cellular Lipidome. PLoS ONE 2014, 9, e97263. [Google Scholar] [CrossRef]
- West, G.; Viitanen, L.; Alm, C.; Mattjus, P.; Salminen, T.A.; Edqvist, J. Identification of a glycosphingolipid transfer protein GLTP1 in Arabidopsis thaliana. FEBS J. 2008, 275, 3421–3437. [Google Scholar] [CrossRef]
- Helmkamp, G.M.; Wirtz, K.W.; Van Deenen, L.L. Phosphatidylinositol exchange protein effects of membrane structure on activity and evidence for a ping-pong mechanism. Arch. Biochem. Biophys. 1976, 174, 592–602. [Google Scholar] [CrossRef]
- Gao, Y.; Chung, T.; Zou, X.; Pike, H.M.; Brown, R.E. Human Glycolipid Transfer Protein (GLTP) Expression Modulates Cell Shape. PLoS ONE 2011, 6, e19990. [Google Scholar] [CrossRef]
- Kjellberg, M.A.; Mattjus, P. Glycolipid Transfer Protein Expression Is Affected by Glycosphingolipid Synthesis. PLoS ONE 2013, 8, e70283. [Google Scholar] [CrossRef]
- Hakomori, S.I. Glycosynaptic microdomains controlling tumor cell phenotype through alteration of cell growth, adhesion, and motility. FEBS Lett. 2010, 584, 1901–1906. [Google Scholar] [CrossRef]
- Camp, E.R.; Patterson, L.D.; Kester, M.; Voelkel-Johnson, C. Therapeutic implications of bioactive sphingolipids: A focus on colorectal cancer. Cancer Biol. Ther. 2017, 18, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Halter, D.; Neumann, S.; Van Dijk, S.M.; Wolthoorn, J.; De Mazière, A.M.; Vieira, O.V.; Mattjus, P.; Klumperman, J.; Van Meer, G.; Sprong, H. Pre- and post-Golgi translocation of glucosylceramide in glycosphingolipid synthesis. J. Cell Biol. 2007, 179, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Mo, J.-S.; Alam, K.J.; Kang, I.-H.; Park, W.C.; Seo, G.-S.; Choi, S.-C.; Kim, H.-S.; Moon, H.-B.; Yun, K.-J.; Chae, S.-C. MicroRNA 196B regulates FAS-mediated apoptosis in colorectal cancer cells. Oncotarget 2015, 6, 2843–2855. [Google Scholar] [CrossRef]
- Mo, J.-S.; Park, Y.-R.; Chae, S.-C. MicroRNA 196B Regulates HOXA5, HOXB6 and GLTP Expression Levels in Colorectal Cancer Cells. Pathol. Oncol. Res. 2018, 25, 953–959. [Google Scholar] [CrossRef]
- Mishra, S.K.; Stephenson, D.J.; Chalfant, C.E.; Brown, R.E. Upregulation of human glycolipid transfer protein (GLTP) induces necroptosis in colon carcinoma cells. Biochim. Biophys. Acta 2019, 1864, 158–167. [Google Scholar] [CrossRef]
- He, S.; Wang, L.; Miao, L.; Wang, T.; Du, F.; Zhao, L.; Wang, X. Receptor interacting protein kinase-3 determines cellular necrotic response to TNF-alpha. Cell 2009, 137, 1100–1111. [Google Scholar] [CrossRef]
- Simanshu, D.K.; Kamlekar, R.K.; Wijesinghe, D.S.; Zou, X.; Zhai, X.; Mishra, S.K.; Molotkovsky, J.G.; Malinina, L.; Hinchcliffe, E.H.; Chalfant, C.E.; et al. Non-vesicular trafficking by a ceramide-1-phosphate transfer protein regulates eicosanoids. Nature 2013, 500, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Zhai, X.; Gao, Y.G.; Mishra, S.K.; Simanshu, D.K.; Boldyrev, I.A.; Benson, L.M.; Bergen, H.R., 3rd; Malinina, L.; Mundy, J.; Molotkovsky, J.G.; et al. Phosphatidylserine Stimulates Ceramide 1-Phosphate (C1P) Intermembrane Transfer by C1P Transfer Proteins. J. Biol. Chem. 2017, 292, 2531–2541. [Google Scholar] [CrossRef] [Green Version]
- Arana, L.; Gangoiti, P.; Ouro, A.; Trueba, M.; Gómez-Muñoz, A.; Arana, L.A. Ceramide and ceramide 1-phosphate in health and disease. Lipids Health Dis. 2010, 9, 15. [Google Scholar] [CrossRef]
- Gómez-Muñoz, A. The Role of Ceramide 1-Phosphate in Tumor Cell Survival and Dissemination. Adv. Cancer Res. 2018, 140, 217–234. [Google Scholar]
- Simanshu, D.K.; Zhai, X.; Munch, D.; Hofius, D.; Markham, J.E.; Bielawski, J.; Bielawska, A.; Malinina, L.; Molotkovsky, J.G.; Mundy, J.W.; et al. Arabidopsis Accelerated Cell Death 11, ACD11, Is a Ceramide-1-Phosphate Transfer Protein and Intermediary Regulator of Phytoceramide Levels. Cell Rep. 2014, 6, 388–399. [Google Scholar] [CrossRef] [Green Version]
- Zarogoulidis, P.; Tsakiridis, K.; Karapantzou, C.; Lampaki, S.; Kioumis, I.; Pitsiou, G.; Papaiwannou, A.; Hohenforst-Schmidt, W.; Huang, H.; Kesisis, G.; et al. Use of Proteins as Biomarkers and Their Role in Carcinogenesis. J. Cancer 2015, 6, 9–18. [Google Scholar] [CrossRef]
- Mishra, S.K.; Gao, Y.-G.; Deng, Y.; Chalfant, C.E.; Hinchcliffe, E.H.; Brown, R.E. CPTP: A sphingolipid transfer protein that regulates autophagy and inflammasome activation. Autophagy 2018, 14, 862–879. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Duo, Y.; Zhai, P.; He, L.; Zhong, K.; Zhang, Y.; Huang, K.; Luo, J.; Zhang, H.; Yu, X. Dual targeting delivery of miR-328 by functionalized mesoporous silica nanoparticles for colorectal cancer therapy. Nanomedicine (London) 2018, 13. [Google Scholar] [CrossRef]
- Mishra, S.K.; Brown, R.E. Abstract 1123: Sphingolipids transfer proteins (GLTP and CPTP) regulate the neoplastic progression of colon and breast cancer cells. Exp. Mol. Ther. 2017, 77, 1123. [Google Scholar]
- Ogretmen, B. Sphingolipid metabolism in cancer signalling and therapy. Nat. Rev. Cancer 2018, 18, 33–50. [Google Scholar] [CrossRef]
- Newton, J.; Lima, S.; Maceyka, M.; Spiegel, S. Revisiting the sphingolipid rheostat: Evolving concepts in cancer therapy. Exp. Cell Res. 2015, 333, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Hisano, Y.; Kobayashi, N.; Kawahara, A.; Yamaguchi, A.; Nishi, T. The Sphingosine 1-Phosphate Transporter, SPNS2, Functions as a Transporter of the Phosphorylated Form of the Immunomodulating Agent FTY720. J. Biol. Chem. 2011, 286, 1758–1766. [Google Scholar] [CrossRef] [Green Version]
- Vu, T.M.; Ishizu, A.-N.; Foo, J.C.; Toh, X.R.; Zhang, F.; Whee, D.M.; Torta, F.; Cazenave-Gassiot, A.; Matsumura, T.; Kim, S.; et al. Mfsd2b is essential for the sphingosine-1-phosphate export in erythrocytes and platelets. Nature 2017, 550, 524–528. [Google Scholar] [CrossRef]
- Kawahara, A.; Nishi, T.; Hisano, Y.; Fukui, H.; Yamaguchi, A.; Mochizuki, N. The Sphingolipid Transporter Spns2 Functions in Migration of Zebrafish Myocardial Precursors. Science 2009, 323, 524–527. [Google Scholar] [CrossRef] [Green Version]
- Fukuhara, S.; Simmons, S.; Kawamura, S.; Inoue, A.; Orba, Y.; Tokudome, T.; Sunden, Y.; Arai, Y.; Moriwaki, K.; Ishida, J.; et al. The sphingosine-1-phosphate transporter Spns2 expressed on endothelial cells regulates lymphocyte trafficking in mice. J. Clin. Investig. 2012, 122, 1416–1426. [Google Scholar] [CrossRef] [Green Version]
- Nagahashi, M.; Kim, E.Y.; Yamada, A.; Ramachandran, S.; Allegood, J.C.; Hait, N.C.; Maceyka, M.; Milstien, S.; Takabe, K.; Spiegel, S. Spns2, a transporter of phosphorylated sphingoid bases, regulates their blood and lymph levels, and the lymphatic network. FASEB J. 2013, 27, 1001–1011. [Google Scholar] [CrossRef]
- Spiegel, S.; Maczis, M.A.; Maceyka, M.; Milstien, S. New insights into functions of the sphingosine-1-phosphate transporter SPNS2. J. Lipid Res. 2019, 60, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Bradley, E.; Dasgupta, S.; Jiang, X.; Zhao, X.; Zhu, G.; He, Q.; Dinkins, M.; Bieberich, E.; Wang, G. Critical Role of Spns2, a Sphingosine-1-Phosphate Transporter, in Lung Cancer Cell Survival and Migration. PLoS ONE 2014, 9, e110119. [Google Scholar] [CrossRef]
- Canals, D.; Jeong, N.; Hernandez-Corbacho, M.; Adada, M.M.; Kelkar, A.D.; Pulkoski-Gross, M.J.; Donaldson, J.C.; A Hannun, Y.; Obeid, L.M. Intracellular sphingosine kinase 2-derived sphingosine-1-phosphate mediates epidermal growth factor-induced ezrin-radixin-moesin phosphorylation and cancer cell invasion. FASEB J. 2015, 29, 4654–4669. [Google Scholar]
- Van der Weyden, L.; Arends, M.J.; Campbell, A.D.; Bald, T.; Wardle-Jones, H.; Griggs, N.; Velasco-Herrera, M.D.; Tüting, T.; Sansom, O.J.; Karp, N.A.; et al. Genome-wide in vivo screen identifies novel host regulators of metastatic colonization. Nature 2017, 541, 233. [Google Scholar] [CrossRef]
- Kobayashi, N.; Kawasaki-Nishi, S.; Otsuka, M.; Hisano, Y.; Yamaguchi, A.; Nishi, T. MFSD2B is a sphingosine 1-phosphate transporter in erythroid cells. Sci. Rep. 2018, 8, 4969. [Google Scholar] [CrossRef]
- van Meer, G.; Halter, D.; Sprong, H.; Somerharju, P.; Egmond, M.R. ABC lipid transporters: Extruders, flippases, or flopless activators? FEBS Lett. 2006, 580, 1171–1177. [Google Scholar] [CrossRef]
- Yamaji, T.; Hanada, K. Sphingolipid metabolism and interorganellar transport: Localization of sphingolipid enzymes and lipid transfer proteins. Traffic 2015, 16, 101–122. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samaha, D.; Hamdo, H.H.; Wilde, M.; Prause, K.; Arenz, C. Sphingolipid-Transporting Proteins as Cancer Therapeutic Targets. Int. J. Mol. Sci. 2019, 20, 3554. https://doi.org/10.3390/ijms20143554
Samaha D, Hamdo HH, Wilde M, Prause K, Arenz C. Sphingolipid-Transporting Proteins as Cancer Therapeutic Targets. International Journal of Molecular Sciences. 2019; 20(14):3554. https://doi.org/10.3390/ijms20143554
Chicago/Turabian StyleSamaha, Doaa, Housam H. Hamdo, Max Wilde, Kevin Prause, and Christoph Arenz. 2019. "Sphingolipid-Transporting Proteins as Cancer Therapeutic Targets" International Journal of Molecular Sciences 20, no. 14: 3554. https://doi.org/10.3390/ijms20143554
APA StyleSamaha, D., Hamdo, H. H., Wilde, M., Prause, K., & Arenz, C. (2019). Sphingolipid-Transporting Proteins as Cancer Therapeutic Targets. International Journal of Molecular Sciences, 20(14), 3554. https://doi.org/10.3390/ijms20143554