RcAP1, a Homolog of APETALA1, is Associated with Flower Bud Differentiation and Floral Organ Morphogenesis in Rosa chinensis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
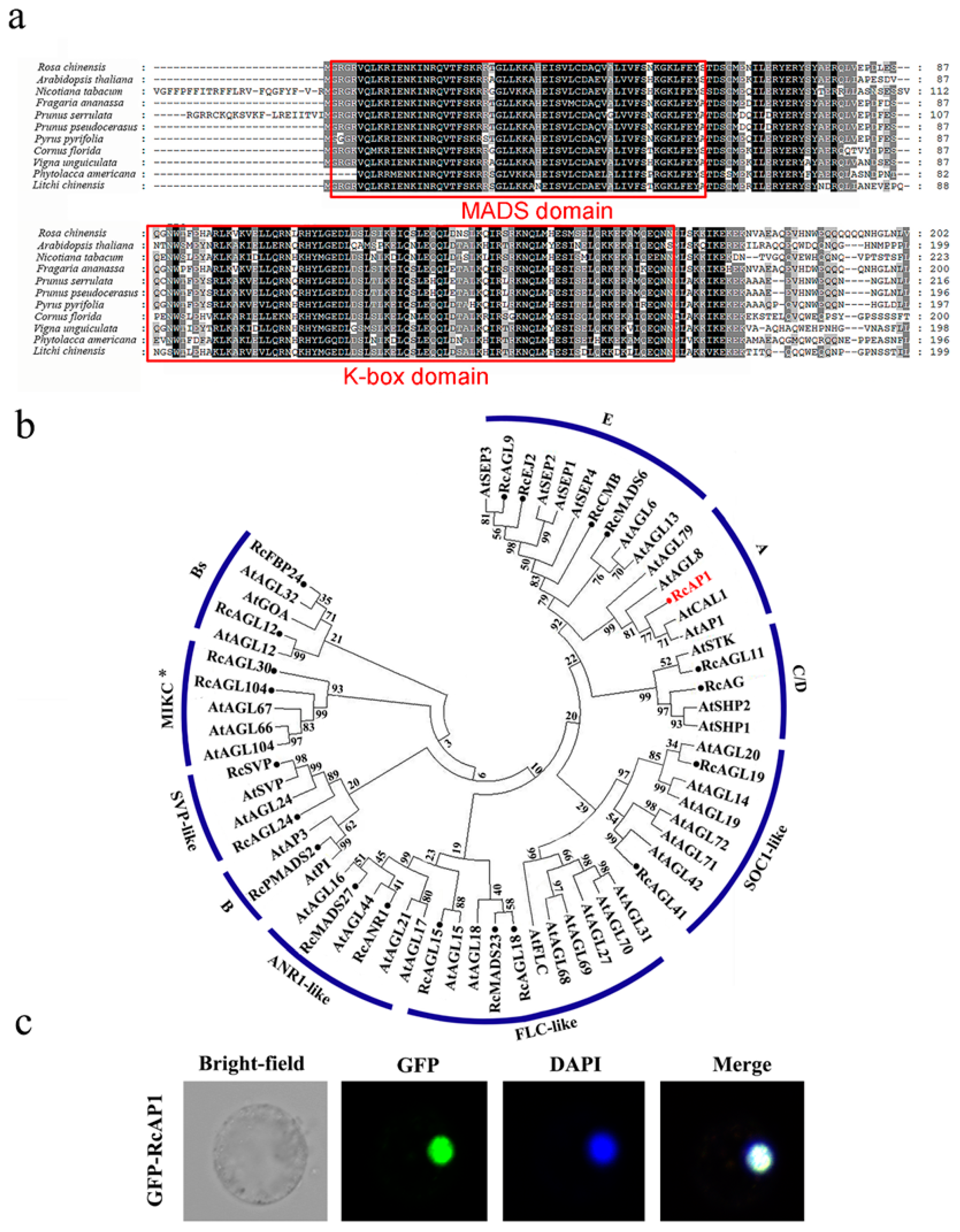
2.1. Bioinformatics Analysis of APETALA1
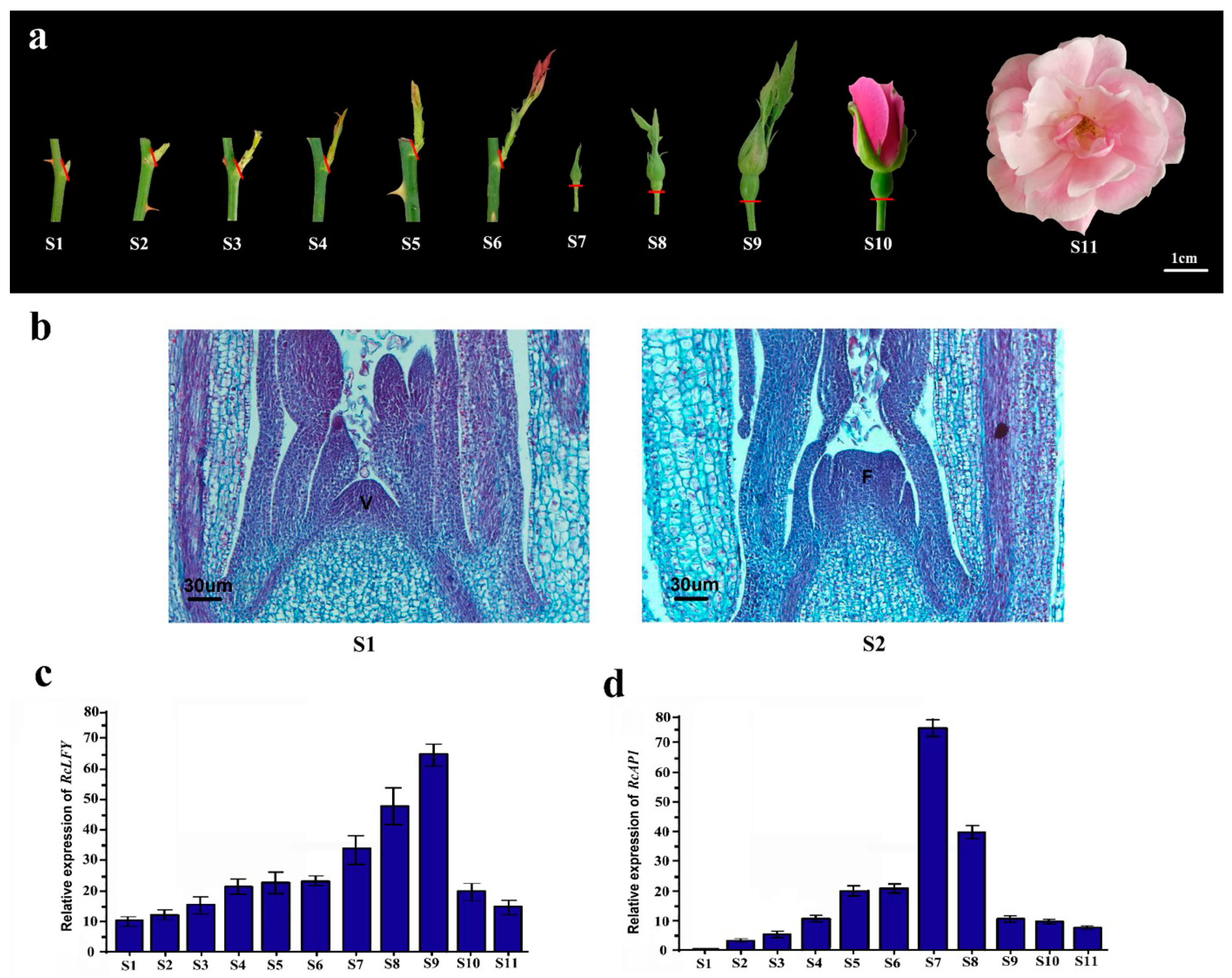
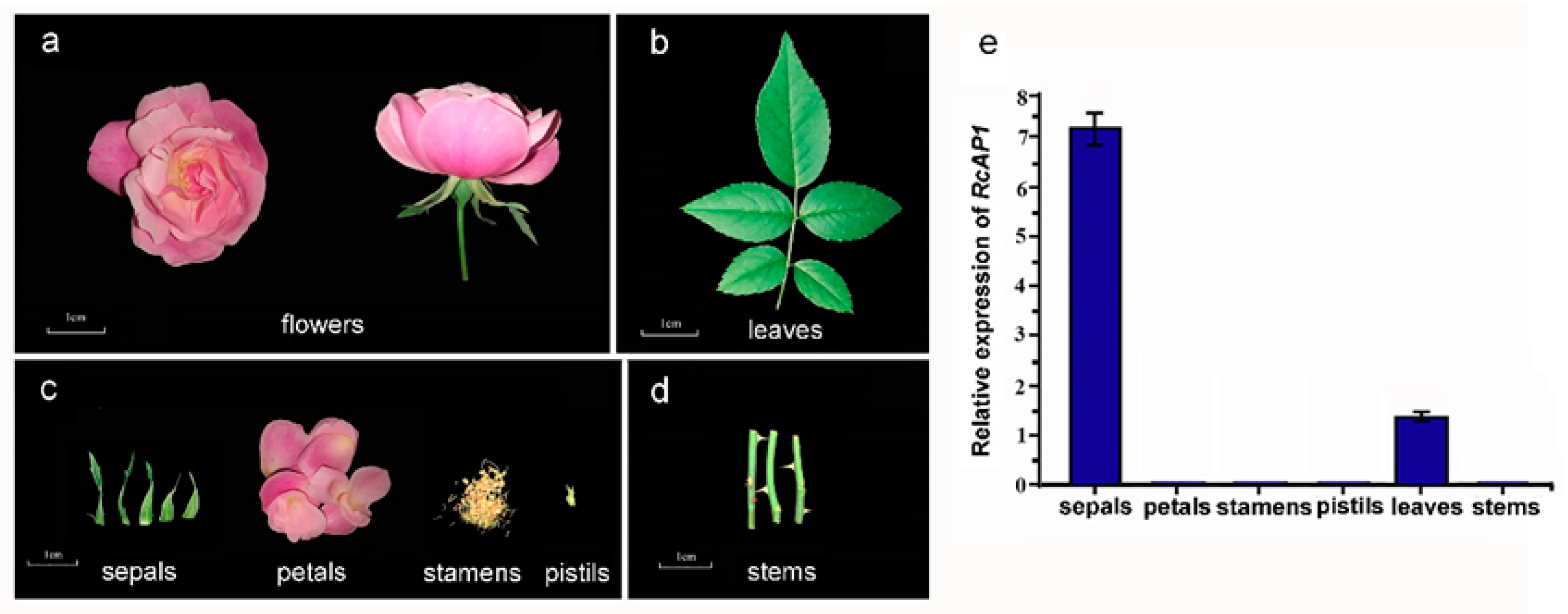
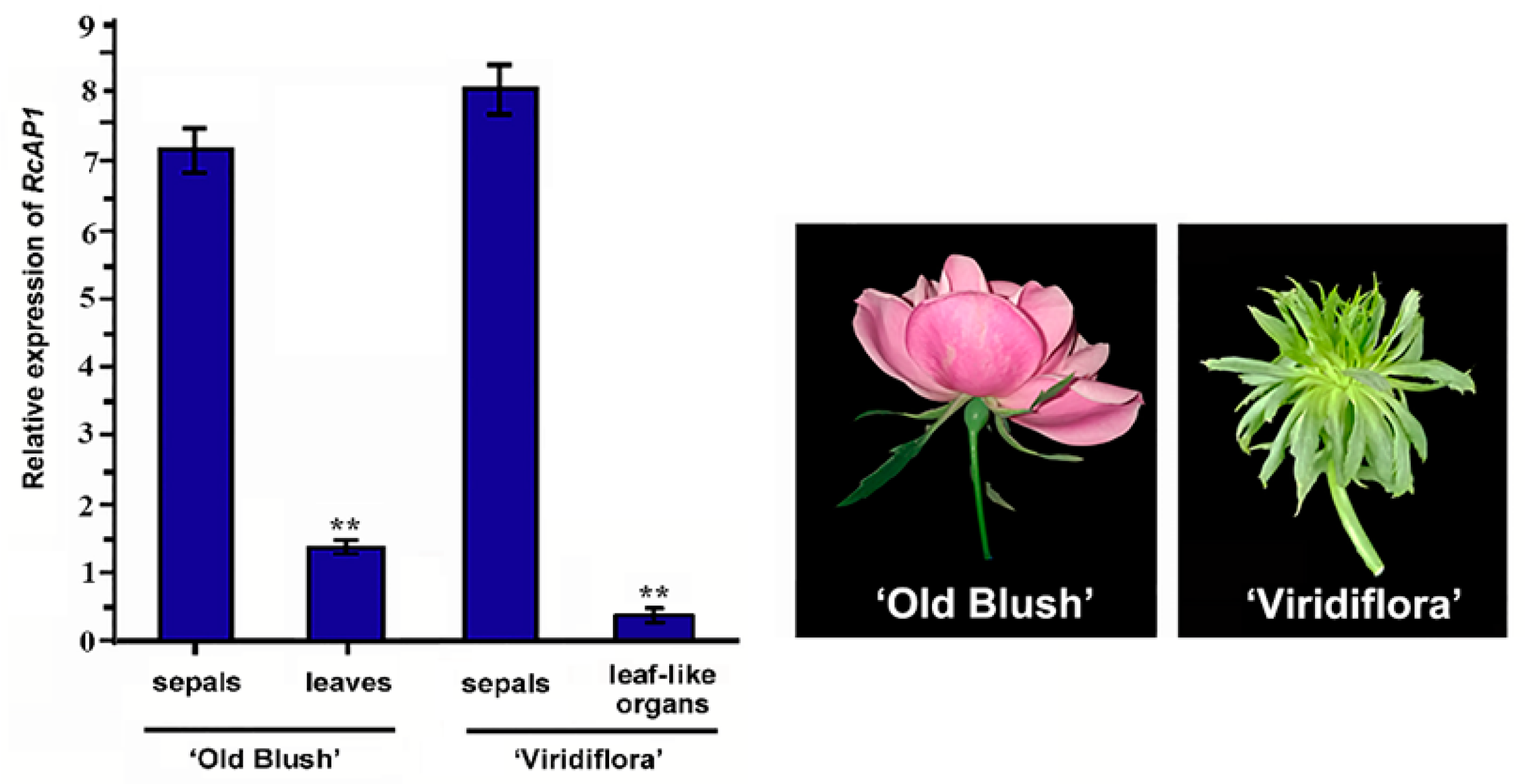
2.2. RcAP1 is Highly Expressed during the Bud Appearance Stage and is Mainly Expressed in Sepals
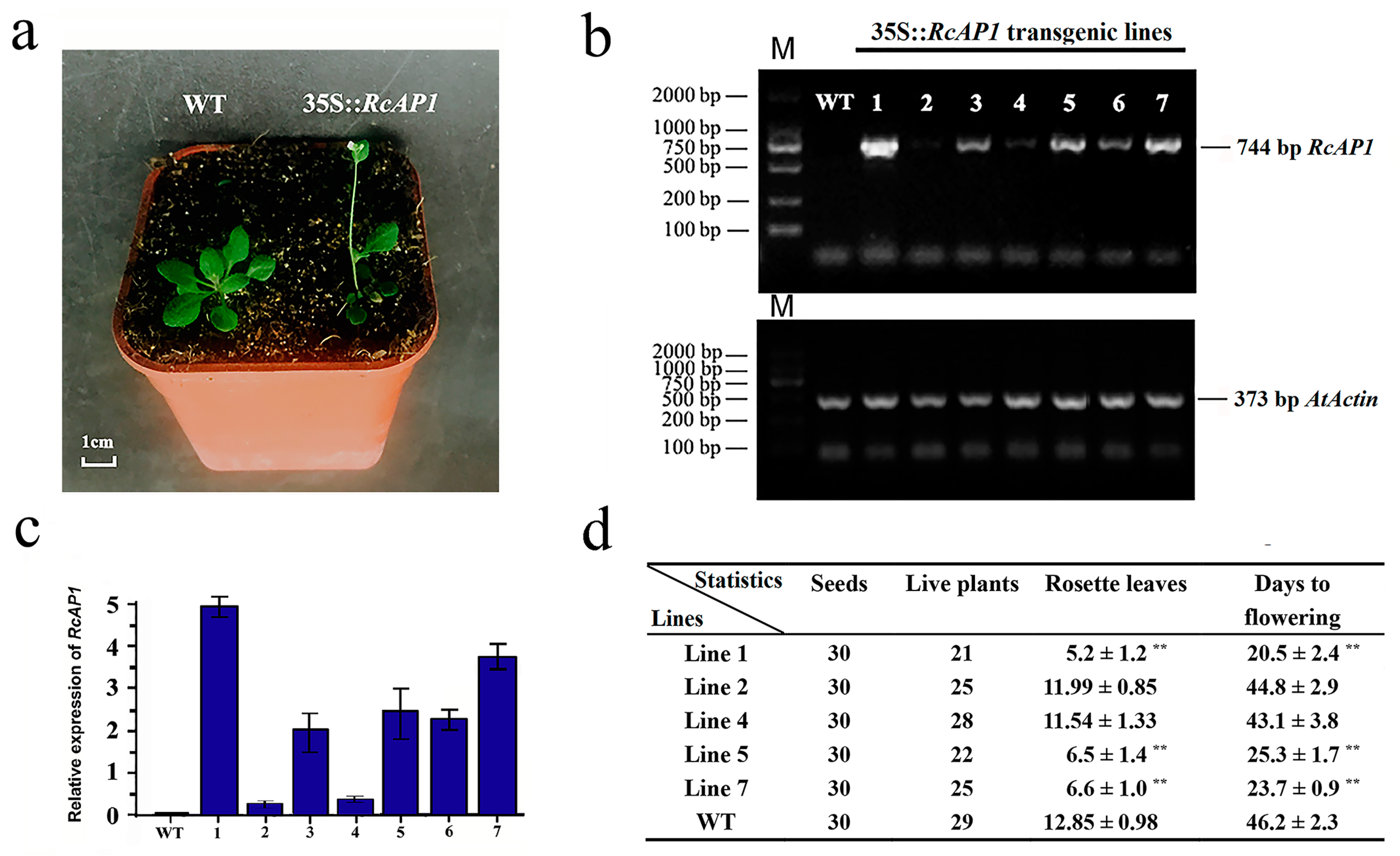
2.3. Overexpression of RcAP1 in Arabidopsis Thaliana Induces Early Flowering
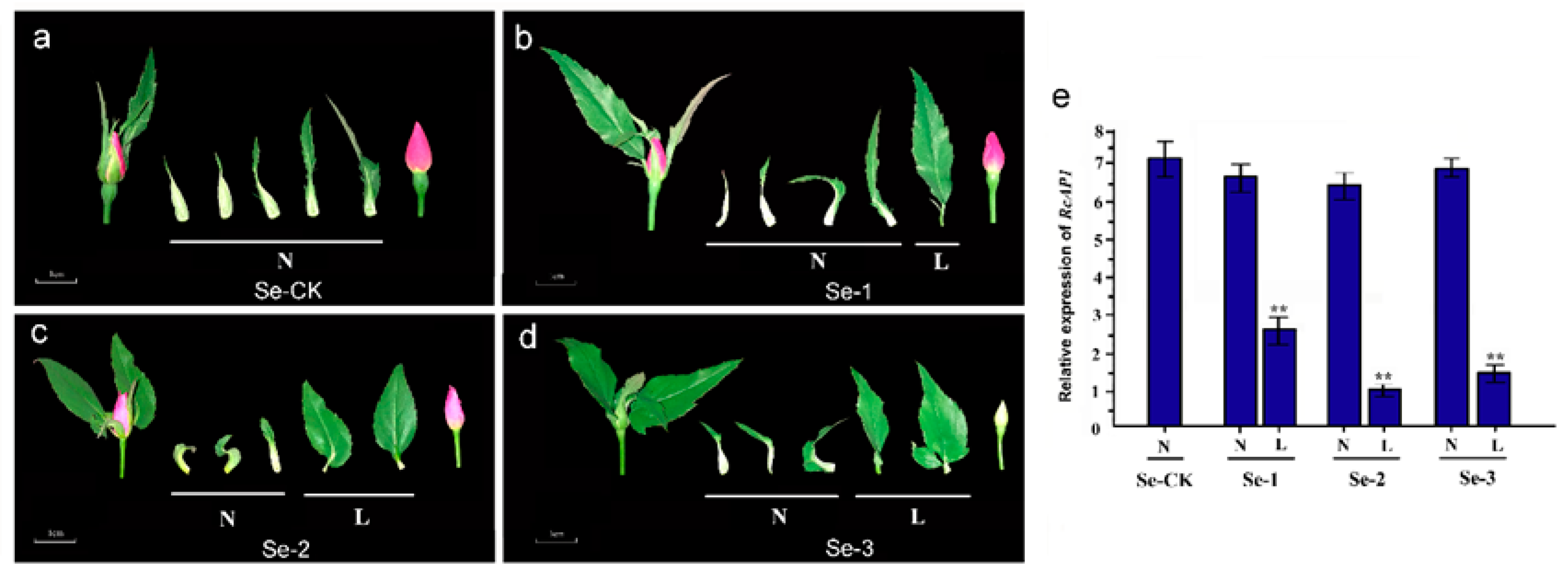
2.4. Knockdown of RcAP1 Expression in ‘Old Blush’ Inhibits Flower Bud Formation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Paraffin Sections and Staining
4.3. RNA and DNA Extraction
4.4. ORF and Promoter Cloning
4.5. Sequence and Phylogenetic Analyses
4.6. qRT-PCR Validation
4.7. Subcellular Localization of RcAP1
4.8. Arabidopsis Thaliana Transformation
4.9. RcAP1 Knockdown in ‘Old Blush’ Plants by the VIGS Method
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| aa | Amino acids |
| VIGS | Virus-induced gene silencing |
| ORF | Open reading frame |
| NCBI | National Center for Biotechnology Information |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| WT | Wild-type |
| RACE | Rapid-amplification of cDNA ends |
| GFP | Green fluorescent protein |
| DAPI | 4′,6-diamino-2-phenylindole |
References
- Thomas, C. Modern Roses XI: The World Encyclopedia of Roses; American Rose Society: Shreveport, LA, USA, 2000; Volume 638. [Google Scholar]
- Cháb, D.; Kolář, J.; Olson, M.S.; Štorchová, H. Two Flowering Locus T (FT) homologs in Chenopodium rubrum differ in expression patterns. Planta 2008, 228, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Yang, H.; Mockler, T.C.; Lin, C. Regulation of Flowering Time by Arabidopsis Photoreceptors. Science 1998, 279, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Ai, X.Y.; Zhang, J.Z. Genetic regulation of flowering time in annual and perennial plants. Wiley Interdiscip. Rev. RNA 2014, 5, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Becker, A. The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Mol. Phylogenet. Evol. 2003, 29, 464–489. [Google Scholar] [CrossRef]
- Smaczniak, C.; Immink, R.G.H.; Angenent, G.C.; Kaufmann, K. Developmental and evolutionary diversity of plant MADS-domain factors: Insights from recent studies. Development 2012, 139, 3081–3098. [Google Scholar] [CrossRef] [PubMed]
- Theissen, G. Development of floral organ identity: Stories from the MADS house. Curr. Opin. Plant Boil. 2001, 4, 75–85. [Google Scholar] [CrossRef]
- Liljegren, S.J.; Gustafson-Brown, C.; Pinyopich, A.; Ditta, G.S.; Yanofsky, M.F. Interactions among APETALA1, LEAFY, and TERMINAL FLOWER1 Specify Meristem Fate. Plant Cell 1999, 11, 1007–1018. [Google Scholar] [CrossRef]
- McCarthy, E.W.; Mohamed, A.; Litt, A. Functional Divergence of APETALA1 and FRUITFULL is due to Changes in both Regulation and Coding Sequence. Front. Plant Sci. 2015, 6, 3099. [Google Scholar] [CrossRef]
- Liu, Z.; Mara, C. Regulatory mechanisms for floral homeotic gene expression. Semin. Cell Dev. Boil. 2010, 21, 80–86. [Google Scholar] [CrossRef]
- Theissen, G.; Becker, A.; Di Rosa, A.; Kanno, A.; Kim, J.T.; Münster, T.; Winter, K.-U.; Saedler, H.; Rosa, A. A short history of MADS-box genes in plants. Plant Mol. Evol. 2000, 42, 115–149. [Google Scholar]
- Theissen, G.; Saedler, H. The golden decade of molecular floral development (1990–1999): A cheerful obituary. Dev. Genet. 1999, 25, 181–193. [Google Scholar]
- Hibino, Y.; Kitahara, K.; Hirai, S.; Matsumoto, S. Structural and functional analysis of rose class B MADS-box genes ‘MASAKO BP, euB3, and B3’: Paleo-type AP3 homologue ‘MASAKO B3’ association with petal development. Plant Sci. 2006, 170, 778–785. [Google Scholar] [CrossRef]
- Kitahara, K.; Matsumoto, S. Rose MADS-box genes ‘MASAKO C1 and D1’ homologous to class C floral identity genes. Plant Sci. 2000, 151, 121–134. [Google Scholar] [CrossRef]
- Mandel, M.A.; Gustafson-Brown, C.; Savidge, B.; Yanofsky, M.F. Molecular characterization of the Arabidopsis floral homeotic gene APETALA1. Nature 1992, 360, 273–277. [Google Scholar] [CrossRef]
- Ferrándiz, C.; Gu, Q.; Martienssen, R.; Yanofsky, M.F. Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1 and CAULIFLOWER. Development 2000, 127, 725. [Google Scholar]
- Liu, C.; Xi, W.; Shen, L.; Tan, C.; Yu, H. Regulation of Floral Patterning by Flowering Time Genes. Dev. Cell 2009, 16, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Samach, A.; Caldeira, K.; Duffy, P.B. Distinct Roles of CONSTANS Target Genes in Reproductive Development of Arabidopsis. Science 2000, 288, 1613–1616. [Google Scholar] [CrossRef]
- Wagner, D. Transcriptional Activation of APETALA1 by LEAFY. Science 1999, 285, 582–584. [Google Scholar] [CrossRef]
- Blázquez, M.A.; Ferrándiz, C.; Madueño, F.; Parcy, F. How Floral Meristems are Built. Plant Mol. Boil. 2006, 60, 855–870. [Google Scholar] [CrossRef]
- Irish, V.F.; Sussex, I.M. Function of the apetala-1 gene during Arabidopsis floral development. Plant Cell 1990, 2, 741–753. [Google Scholar]
- Shchennikova, A.V.; Shulga, O.A.; Immink, R.; Skryabin, K.G.; Angenent, G.C. Identification and Characterization of Four Chrysanthemum MADS-Box Genes, Belonging to the APETALA1/FRUITFULL and SEPALLATA3 Subfamilies1. Plant Physiol. 2004, 134, 1632–1641. [Google Scholar] [CrossRef]
- Chen, M.-K.; Lin, I.-C.; Yang, C.-H. Functional Analysis of Three Lily (Lilium longiflorum) APETALA1-like MADS Box Genes in Regulating Floral Transition and Formation. Plant Cell Physiol. 2008, 49, 704–717. [Google Scholar] [CrossRef]
- Pabón-Mora, N.; Ambrose, B.A.; Litt, A. Poppy APETALA1/FRUITFULL Orthologs Control Flowering Time, Branching, Perianth Identity, and Fruit Development. Plant Physiol. 2012, 158, 1685–1704. [Google Scholar] [CrossRef]
- Sun, Y.K.; Fan, Z.Q.; Li, X.L.; Li, J.Y.; Yin, H.F. The APETALA1 and FRUITFUL homologs in Camellia japonica and their roles in double flower domestication. Mol. Breed. 2014, 33, 821–834. [Google Scholar] [CrossRef]
- Chuang, T.-H.; Li, K.-H.; Li, P.-F.; Yang, C.-H. Functional analysis of an APETALA1-like MADS box gene from Eustoma grandiflorum in regulating floral transition and formation. Plant Biotechnol. Rep. 2018, 12, 115–125. [Google Scholar] [CrossRef]
- Fernando, D.D.; Zhang, S.L. Constitutive expression of the SAP1 gene from willow (Salix discolor) causes early flowering in Arabidopsis thaliana. Dev. Genes Evol. 2006, 216, 19–28. [Google Scholar] [CrossRef]
- Tang, M.; Tao, Y.-B.; Xu, Z.-F.; McCormick, S. Ectopic expression of Jatropha curcas APETALA1 (JcAP1) caused early flowering in Arabidopsis, but not in Jatropha. PeerJ 2016, 4, 1969. [Google Scholar] [CrossRef]
- Bowman, J.L.; Alvarez, J.; Weigel, D.; Meyerowitz, E.M.; Smyth, D.R. Control of flower development in Arabidopsis thaliana by APETALA1 and interacting genes. Development 1993, 119, 721. [Google Scholar]
- Litt, A.; Irish, V.F. Duplication and diversification in the APETALA1/FRUITFULL floral homeotic gene lineage: Implications for the evolution of floral development. Genetics 2003, 165, 821–833. [Google Scholar]
- Smyth, D. A reverse trend—MADS functions revealed. Trends Plant Sci. 2000, 5, 315–317. [Google Scholar] [CrossRef]
- Wang, L.N.; Liu, Y.F.; Zhang, Y.M.; Fang, R.X.; Liu, Q.L. The expression level of Rosa Terminal Flower 1 (RTFL1) is related with recurrent flowering in roses. Mol. Biol. Rep. 2012, 39, 3737–3746. [Google Scholar] [CrossRef]
- Parcy, F.; Nilsson, O.; Busch, M.A.; Lee, I.; Weigel, D. A genetic framework for floral patterning. Nature 1998, 395, 561–566. [Google Scholar] [CrossRef]
- William, D.A.; Su, Y.; Smith, M.R.; Lu, M.; Baldwin, D.A.; Wagner, D. Genomic identification of direct target genes of LEAFY. Proc. Natl. Acad. Sci. USA 2004, 101, 1775. [Google Scholar] [CrossRef]
- Remay, A.; Lalanne, D.; Thouroude, T.; Le Couviour, F.; Oyant, L.H.-S.; Foucher, F.; Couviour, F. A survey of flowering genes reveals the role of gibberellins in floral control in rose. Theor. Appl. Genet. 2009, 119, 767–781. [Google Scholar] [CrossRef]
- Liu, C.; Zhou, J.; Bracha-Drori, K.; Yalovsky, S.; Ito, T.; Yu, H. Specification of Arabidopsis floral meristem identity by repression of flowering time genes. Development 2007, 134, 1901–1910. [Google Scholar] [CrossRef]
- Mandel, M.A.; Yanofsky, M.F. The Arabidopsis AGL8 MADS box gene is expressed in inflorescence meristems and is negatively regulated by APETALA1. Plant Cell 1995, 7, 1763–1771. [Google Scholar]
- Melzer, R.; Wang, Y.-Q.; Theißen, G. The naked and the dead: The ABCs of gymnosperm reproduction and the origin of the angiosperm flower. Semin. Cell Dev. Boil. 2010, 21, 118–128. [Google Scholar] [CrossRef]
- Pollock, R.; Treisman, R. Human SRF-related proteins: DNA-binding properties and potential regulatory targets. Genes Dev. 1991, 5, 2327–2341. [Google Scholar] [CrossRef]
- Randoux, M.; Jeauffre, J.; Thouroude, T.; Vasseur, F.; Hamama, L.; Juchaux, M.; Sakr, S.; Foucher, F. Gibberellins regulate the transcription of the continuous flowering regulator, RoKSN, a rose TFL1 homologue. J. Exp. Bot. 2012, 63, 6543–6554. [Google Scholar] [CrossRef]
- Liu, Y.X.; Song, H.W.; Liu, Z.L.; Hu, G.B.; Lin, S.Q. Molecular characterization of loquat EjAP1 gene in relation to flowering. Plant Growth Regul. 2013, 70, 287–296. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, X.; Yan, G.; Zhou, Y.; Zhang, K. Over-expression of the PaAP1 gene from sweet cherry (Prunus avium L.) causes early flowering in Arabidopsis thaliana. J. Plant Physiol. 2013, 170, 315–320. [Google Scholar] [CrossRef]
- Liu, Y.X.; Kong, J.; Li, T.Z.; Wang, Y.; Wang, A.D.; Han, Z.H. Isolation and Characterization of an APETALA1-Like Gene from Pear (Pyrus pyrifolia). Plant Mol. Biol. Report. 2013, 31, 1031–1039. [Google Scholar] [CrossRef]
- Mibus, H.; Heckl, D.; Serek, M. Cloning and Characterization of Three APETALA1/FRUITFULL-like Genes in Different Flower Types of Rosa × hybrida L. J. Plant Growth Regul. 2011, 30, 272–285. [Google Scholar] [CrossRef]
- Yan, H.J.; Zhang, H.; Wang, Q.G.; Jian, H.Y.; Qiu, X.Q.; Baudino, S.; Just, J.; Raymond, O.; Gu, L.F.; Wang, J.H.; et al. The Rosa chinensis cv. Viridiflora Phyllody Phenotype Is Associated with Misexpression of Flower Organ Identity Genes. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.-M.; Zhang, J.-Z.; Mei, L.; Hu, C.-G. Molecular cloning, promoter analysis and functional characterization of APETALA 1-like gene from precocious trifoliate orange (Poncirus trifoliata L. Raf.). Sci. Hortic. 2014, 178, 95–105. [Google Scholar] [CrossRef]
- Sawettalake, N.; Bunnag, S.; Wang, Y.W.; Shen, L.S.; Yu, H. DOAP1 Promotes Flowering in the Orchid Dendrobium Chao Praya Smile. Front. Plant Sci. 2016, 8. [Google Scholar] [CrossRef]
- Yan, H.J.; Shi, S.C.; Ma, N.; Cao, X.Q.; Zhang, H.; Qiu, X.Q.; Wang, Q.; Jian, H.Y.; Zhou, N.N.; Zhang, Z.; et al. Graft-accelerated virus-induced gene silencing facilitates functional genomics in rose flowers. J. Integr. Plant Biol. 2018, 60, 34–44. [Google Scholar] [CrossRef]
- García-Ponce, B.; Garay-Arroyo, A.; Alvarez-Buylla, E.R. Unique and redundant functional domains of APETALA1 and CAULIFLOWER, two recently duplicated Arabidopsis thaliana floral MADS-box genes. J. Exp. Bot. 2006, 57, 3099–3107. [Google Scholar] [Green Version]
- Sakai, W.S. Simple Method for Differential Staining of Paraffin Embedded Plant Material Using Toluidine Blue O. Stain. Technol. 1973, 48, 247–249. [Google Scholar] [CrossRef]
- Lescot, M. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Solovyev, V.V.; Shahmuradov, I.A.; Salamov, A.A. Identification of Promoter Regions and Regulatory Sites. Evol. Genom. 2010, 674, 57–83. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sheen, J.; Hwang, S.; Niwa, Y.; Kobayashi, H.; Galbraith, D.W. Green-fluorescent protein as a new vital marker in plant cells. Plant J. 1995, 8, 777–784. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; Tang, A.; Yu, J.; Cheng, T.; Wang, J.; Yang, W.; Pan, H.; Zhang, Q. RcAP1, a Homolog of APETALA1, is Associated with Flower Bud Differentiation and Floral Organ Morphogenesis in Rosa chinensis. Int. J. Mol. Sci. 2019, 20, 3557. https://doi.org/10.3390/ijms20143557
Han Y, Tang A, Yu J, Cheng T, Wang J, Yang W, Pan H, Zhang Q. RcAP1, a Homolog of APETALA1, is Associated with Flower Bud Differentiation and Floral Organ Morphogenesis in Rosa chinensis. International Journal of Molecular Sciences. 2019; 20(14):3557. https://doi.org/10.3390/ijms20143557
Chicago/Turabian StyleHan, Yu, Aoying Tang, Jiayao Yu, Tangren Cheng, Jia Wang, Weiru Yang, Huitang Pan, and Qixiang Zhang. 2019. "RcAP1, a Homolog of APETALA1, is Associated with Flower Bud Differentiation and Floral Organ Morphogenesis in Rosa chinensis" International Journal of Molecular Sciences 20, no. 14: 3557. https://doi.org/10.3390/ijms20143557
APA StyleHan, Y., Tang, A., Yu, J., Cheng, T., Wang, J., Yang, W., Pan, H., & Zhang, Q. (2019). RcAP1, a Homolog of APETALA1, is Associated with Flower Bud Differentiation and Floral Organ Morphogenesis in Rosa chinensis. International Journal of Molecular Sciences, 20(14), 3557. https://doi.org/10.3390/ijms20143557