The Role of Leukotrienes as Potential Therapeutic Targets in Allergic Disorders


Abstract
:1. Introduction
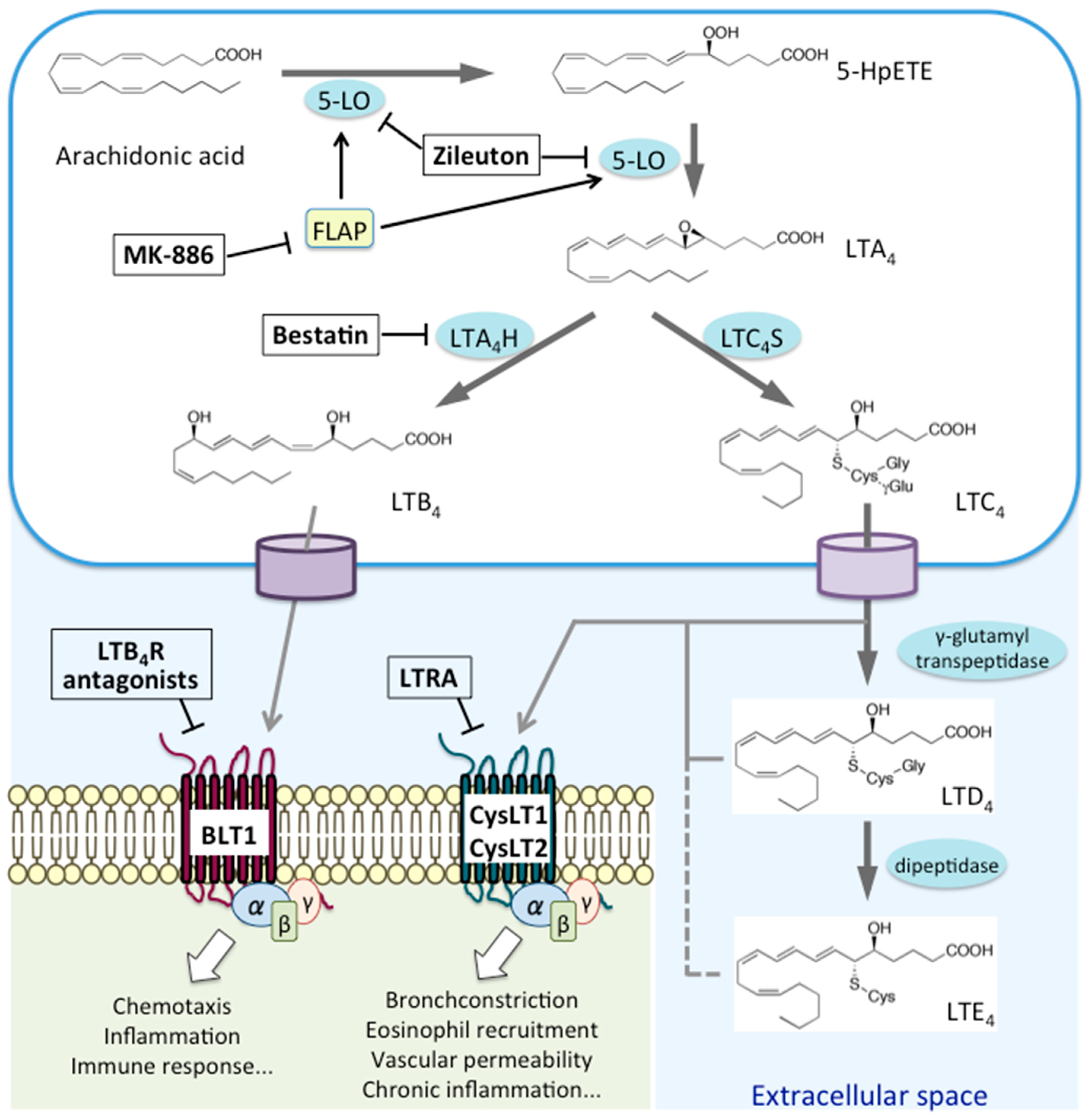
2. Biosynthesis and Metabolism of LTs
3. Expression of LT Receptors and Their Associated Signaling Pathways
3.1. LTB4 Receptors: BLT1 and BLT2
3.2. CysLTs Receptors: CysLT1 and CysLT2
3.3. GPR99
4. LTs and Allergic Diseases
4.1. Asthma
4.1.1. Pathology
4.1.2. The LTB4–BLT1 Pathway in Asthma
4.1.3. The CysLT Pathway in Asthma
4.2. Exercise-Induced Asthma (EIA)
4.3. Aspirin-Sensitive Asthma (ASA)
4.4. Allergic Rhinitis (AR)
4.5. Atopic Dermatitis (AD)
4.6. Allergic Conjunctivitis
4.7. Anaphylaxis
5. Urinary LTE4 as a Biomarker of Allergic Disease
6. Other Diseases
7. Conclusions
Conflicts of Interest
Abbreviations
| PG | prostaglandin |
| LT | leukotriene |
| AA | arachidonic acid |
| GPCR | G-protein-coupled receptor |
| LTB4 | leukotriene B4 |
| PLA2 | phospholipase A2 |
| 5-LO | 5-lipoxygenase |
| COX | cyclooxygenase |
| 5-HpETE | 5-hydroxyperoxyeicosatetraenoic acid |
| LTA4 | leukotriene A4 |
| LTA4H | leukotriene A4 hydrolase |
| LTC4S | leukotriene C4 synthase |
| COPD | chronic obstructive pulmonary disease |
| AMD | age-related macular degeneration |
| BLT1 | LTB4 receptor 1 |
| BLT2 | LTB4 receptor 2 |
| 12-HHT | 12(S)-hydroxyheptadeca-5Z,8E,10E-trienoic acid |
| OXGR1 | oxoglutarate receptor |
| ICS | inhaled corticosteroid |
| AHR | hyper-responsiveness |
| BAL | bronchoalveolar lavage |
| EIA | Exercise-induced asthma |
| LTRA | leukotriene receptor antagonist |
| FLAP | five-lipoxygenase-activating protein |
| EBC | exhaled breath condensate |
| ASA | aspirin-sensitive asthma |
| AERD | aspirin-exacerbated respiratory disease |
| IL | interleukin |
| AR | allergic rhinitis |
| SAH | H1-antihistamines |
| AD | atopic dermatitis |
| MC | mast cell |
| MCAS | mast cell activation syndrome |
| SM | systemic mastocytosis |
| uLTE4 | urinary LTE4 |
References
- Shimizu, T. Lipid mediators in health and disease: Enzymes and receptors as therapeutic targets for the regulation of immunity and inflammation. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 123–150. [Google Scholar] [CrossRef] [PubMed]
- Radmark, O.; Werz, O. 5-Lipoxygenase, a key enzyme for leukotriene biosynthesis in health and disease. Biochim. Biophys. Acta 2015, 1851, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Back, M.; Powell, W.S. Update on leukotriene, lipoxin and oxoeicosanoid receptors: IUPHAR Review 7. Br. J. Pharmacol. 2014, 171, 3551–3574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Yokomizo, T. The role of leukotrienes in allergic diseases. Allergol. Int. 2015, 64, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Haeggstrom, J.Z.; Funk, C.D. Lipoxygenase and leukotriene pathways: Biochemistry, biology, and roles in disease. Chem. Rev. 2011, 111, 5866–5898. [Google Scholar] [CrossRef] [PubMed]
- White, A.A.; Stevenson, D.D. Aspirin-Exacerbated Respiratory Disease. N. Engl. J. Med. 2018, 379, 1060–1070. [Google Scholar] [CrossRef]
- Cuzzo, B.; Lappin, S.L. Physiology, Leukotrienes; StatPearls: Treasure Island, FL, USA, 2019. [Google Scholar]
- Satpathy, S.R.; Jala, V.R. Crystalline silica-induced leukotriene B4-dependent inflammation promotes lung tumour growth. Nat. Commun. 2015, 6, 7064. [Google Scholar] [CrossRef]
- Jala, V.R.; Bodduluri, S.R. The yin and yang of leukotriene B4 mediated inflammation in cancer. Semin. Immunol. 2017, 33, 58–64. [Google Scholar] [CrossRef]
- Park, J.; Jang, J.H. BLT2, a leukotriene B4 receptor 2, as a novel prognostic biomarker of triple-negative breast cancer. BMB Rep. 2018, 51, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Houthuijzen, J.M.; Daenen, L.G. Lysophospholipids secreted by splenic macrophages induce chemotherapy resistance via interference with the DNA damage response. Nat. Commun. 2014, 5, 5275. [Google Scholar] [CrossRef] [Green Version]
- Mathis, S.P.; Jala, V.R. Nonredundant roles for leukotriene B4 receptors BLT1 and BLT2 in inflammatory arthritis. J. Immunol. 2010, 185, 3049–3056. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, Y.; Okuno, T. Protective role of the leukotriene B4 receptor BLT2 in murine inflammatory colitis. FASEB J. 2010, 24, 4678–4690. [Google Scholar] [CrossRef] [PubMed]
- Paruchuri, S.; Tashimo, H. Leukotriene E4-induced pulmonary inflammation is mediated by the P2Y12 receptor. J. Exp. Med. 2009, 206, 2543–2555. [Google Scholar] [CrossRef] [PubMed]
- Foster, H.R.; Fuerst, E. Characterisation of P2Y(12) receptor responsiveness to cysteinyl leukotrienes. PLoS ONE 2013, 8, e58305. [Google Scholar] [CrossRef] [PubMed]
- Bankova, L.G.; Lai, J. Leukotriene E4 elicits respiratory epithelial cell mucin release through the G-protein-coupled receptor, GPR99. Proc. Natl. Acad. Sci. USA 2016, 113, 6242–6247. [Google Scholar] [CrossRef] [PubMed]
- Ciana, P.; Fumagalli, M. The orphan receptor GPR17 identified as a new dual uracil nucleotides/cysteinyl-leukotrienes receptor. EMBO J. 2006, 25, 4615–4627. [Google Scholar] [CrossRef]
- Davenport, A.P.; Alexander, S.P. International Union of Basic and Clinical Pharmacology. LXXXVIII. G protein-coupled receptor list: Recommendations for new pairings with cognate ligands. Pharmacol. Rev. 2013, 65, 967–986. [Google Scholar] [CrossRef]
- Krishnamoorthy, S.; Recchiuti, A. Resolvin D1 binds human phagocytes with evidence for proresolving receptors. Proc. Natl. Acad. Sci. USA 2010, 107, 1660–1665. [Google Scholar] [CrossRef] [Green Version]
- Chiang, N.; Fredman, G. Infection regulates pro-resolving mediators that lower antibiotic requirements. Nature 2012, 484, 524–528. [Google Scholar] [CrossRef] [Green Version]
- Dalli, J.; Winkler, J.W. Resolvin D3 and aspirin-triggered resolvin D3 are potent immunoresolvents. Chem. Biol. 2013, 20, 188–201. [Google Scholar] [CrossRef]
- Wittamer, V.; Gregoire, F. The C-terminal nonapeptide of mature chemerin activates the chemerin receptor with low nanomolar potency. J. Biol. Chem. 2004, 279, 9956–9962. [Google Scholar] [CrossRef] [PubMed]
- Wittamer, V.; Franssen, J.D. Specific recruitment of antigen-presenting cells by chemerin, a novel processed ligand from human inflammatory fluids. J. Exp. Med. 2003, 198, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Arita, M.; Bianchini, F. Stereochemical assignment, antiinflammatory properties, and receptor for the omega-3 lipid mediator resolvin E1. J. Exp. Med. 2005, 201, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, T. Two distinct leukotriene B4 receptors, BLT1 and BLT2. J. BioChem. 2015, 157, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, T.; Izumi, T. A G-protein-coupled receptor for leukotriene B4 that mediates chemotaxis. Nature 1997, 387, 620–624. [Google Scholar] [CrossRef]
- Qiu, H.; Johansson, A.S. Differential induction of BLT receptor expression on human endothelial cells by lipopolysaccharide, cytokines, and leukotriene B4. Proc. Natl. Acad. Sci. USA 2006, 103, 6913–6918. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Arita, M. Leukotriene B4 and lipoxin A4 are regulatory signals for neural stem cell proliferation and differentiation. FASEB J. 2006, 20, 1785–1792. [Google Scholar] [CrossRef]
- Sun, R.; Ba, X. Leukotriene B4 regulates proliferation and differentiation of cultured rat myoblasts via the BLT1 pathway. Mol. Cells 2009, 27, 403–408. [Google Scholar] [CrossRef]
- Yokomizo, T.; Kato, K. A second leukotriene B(4) receptor, BLT2. A new therapeutic target in inflammation and immunological disorders. J. Exp. Med. 2000, 192, 421–432. [Google Scholar] [CrossRef]
- Yokomizo, T.; Ogawa, Y. cDNA cloning, expression, and mutagenesis study of leukotriene B4 12-hydroxydehydrogenase. J. Biol. Chem. 1996, 271, 2844–2850. [Google Scholar] [CrossRef]
- Okuno, T.; Iizuka, Y. 12(S)-Hydroxyheptadeca-5Z, 8E, 10E-trienoic acid is a natural ligand for leukotriene B4 receptor 2. J. Exp. Med. 2008, 205, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Saeki, K. 12-Hydroxyheptadecatrienoic acid promotes epidermal wound healing by accelerating keratinocyte migration via the BLT2 receptor. J. Exp. Med. 2014, 211, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, S.; Koga, T. Non-steroidal anti-inflammatory drug delays corneal wound healing by reducing production of 12-hydroxyheptadecatrienoic acid, a ligand for leukotriene B4 receptor 2. Sci. Rep. 2017, 7, 13267. [Google Scholar] [CrossRef] [PubMed]
- Shigematsu, M.; Koga, T. Leukotriene B4 receptor type 2 protects against pneumolysin-dependent acute lung injury. Sci. Rep. 2016, 6, 34560. [Google Scholar] [CrossRef] [PubMed]
- Saeki, K.; Yokomizo, T. Identification, signaling, and functions of LTB4 receptors. Semin. Immunol. 2017, 33, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Thompson-Souza, G.A.; Gropillo, I. Cysteinyl Leukotrienes in Eosinophil Biology: Functional Roles and Therapeutic Perspectives in Eosinophilic Disorders. Front. Med. (Lausanne) 2017, 4, 106. [Google Scholar] [CrossRef]
- Wunder, F.; Tinel, H. Pharmacological characterization of the first potent and selective antagonist at the cysteinyl leukotriene 2 (CysLT(2)) receptor. Br. J. Pharmacol. 2010, 160, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.R.; O’Neill, G.P. Characterization of the human cysteinyl leukotriene CysLT1 receptor. Nature 1999, 399, 789–793. [Google Scholar] [CrossRef]
- Heise, C.E.; O’Dowd, B.F. Characterization of the human cysteinyl leukotriene 2 receptor. J. Biol. Chem. 2000, 275, 30531–30536. [Google Scholar] [CrossRef]
- Mechiche, H.; Naline, E. Effects of cysteinyl leukotrienes in small human bronchus and antagonist activity of montelukast and its metabolites. Clin. Exp. Allergy 2003, 33, 887–894. [Google Scholar] [CrossRef]
- Figueroa, D.J.; Breyer, R.M. Expression of the cysteinyl leukotriene 1 receptor in normal human lung and peripheral blood leukocytes. Am. J. Respir. Crit. Care Med. 2001, 163, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Hui, Y.; Cheng, Y. Directed vascular expression of human cysteinyl leukotriene 2 receptor modulates endothelial permeability and systemic blood pressure. Circulation 2004, 110, 3360–3366. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, A.; Kanaoka, Y. Functional recognition of a distinct receptor preferential for leukotriene E4 in mice lacking the cysteinyl leukotriene 1 and 2 receptors. Proc. Natl. Acad. Sci. USA 2008, 105, 16695–16700. [Google Scholar] [CrossRef] [PubMed]
- Kanaoka, Y.; Maekawa, A. Identification of GPR99 protein as a potential third cysteinyl leukotriene receptor with a preference for leukotriene E4 ligand. J. Biol. Chem. 2013, 288, 10967–10972. [Google Scholar] [CrossRef] [PubMed]
- Shirasaki, H.; Kanaizumi, E. Expression and localization of GPR99 in human nasal mucosa. Auris Nasus Larynx 2017, 44, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, B.C.; Rabinovitch, N. Urinary Leukotriene E4 as a Biomarker of Exposure, Susceptibility, and Risk in Asthma: An Update. Immunol. Allergy Clin. N. Am. 2018, 38, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Peebles, R.S., Jr.; Aronica, M.A. Proinflammatory Pathways in the Pathogenesis of Asthma. Clin. Chest Med. 2019, 40, 29–50. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, E.W. Importance of the leukotriene B4-BLT1 and LTB4-BLT2 pathways in asthma. Semin. Immunol. 2017, 33, 44–51. [Google Scholar] [CrossRef]
- Miligkos, M.; Bannuru, R.R. Leukotriene-receptor antagonists versus placebo in the treatment of asthma in adults and adolescents: A systematic review and meta-analysis. Ann. Intern. Med. 2015, 163, 756–767. [Google Scholar] [CrossRef]
- Bruno, F.; Spaziano, G. Recent advances in the search for novel 5-lipoxygenase inhibitors for the treatment of asthma. Eur. J. Med. Chem. 2018, 153, 65–72. [Google Scholar] [CrossRef]
- Matsunaga, Y.; Fukuyama, S. Leukotriene B4 receptor BLT2 negatively regulates allergic airway eosinophilia. FASEB J. 2013, 27, 3306–3314. [Google Scholar] [CrossRef] [PubMed]
- Kandhare, A.D.; Liu, Z. Therapeutic Potential of Morin in Ovalbumin-induced Allergic Asthma Via Modulation of SUMF2/IL-13 and BLT2/NF-kB Signaling Pathway. Curr. Mol. Pharmacol. 2019, 12, 122–138. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Shen, J. Leukotriene B4 receptor 2 regulates the proliferation, migration, and barrier integrity of bronchial epithelial cells. J. Cell. Physiol. 2018, 233, 6117–6124. [Google Scholar] [CrossRef] [PubMed]
- Seymour, M.L.; Rak, S. Leukotriene and prostanoid pathway enzymes in bronchial biopsies of seasonal allergic asthmatics. Am. J. Respir. Crit. Care Med. 2001, 164, 2051–2056. [Google Scholar] [CrossRef] [PubMed]
- Zaitsu, M.; Hamasaki, Y. Leukotriene synthesis is increased by transcriptional up-regulation of 5-lipoxygenase, leukotriene A4 hydrolase, and leukotriene C4 synthase in asthmatic children. J. Asthma 2003, 40, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Pal, K.; Feng, X. Leukotriene A4 Hydrolase Activation and Leukotriene B4 Production by Eosinophils in Severe Asthma. Am. J. Respir. Cell Mol. Biol. 2019, 60, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Kazani, S.; Planaguma, A. Exhaled breath condensate eicosanoid levels associate with asthma and its severity. J. Allergy Clin. Immunol. 2013, 132, 547–553. [Google Scholar] [CrossRef] [Green Version]
- Trischler, J.; Muller, C.M. Elevated exhaled leukotriene B(4) in the small airway compartment in children with asthma. Ann. Allergy Asthma Immunol. 2015, 114, 111–116. [Google Scholar] [CrossRef]
- Ohnishi, H.; Miyahara, N. The role of leukotriene B(4) in allergic diseases. Allergol. Int. 2008, 57, 291–298. [Google Scholar] [CrossRef]
- Loutsios, C.; Farahi, N. Biomarkers of eosinophilic inflammation in asthma. Expert Rev. Respir. Med. 2014, 8, 143–150. [Google Scholar] [CrossRef]
- Ito, K.; Herbert, C. Steroid-resistant neutrophilic inflammation in a mouse model of an acute exacerbation of asthma. Am. J. Respir. Cell Mol. Biol. 2008, 39, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; He, X.Y. Different inflammatory phenotypes in adults and children with acute asthma. Eur. Respir. J. 2011, 38, 567–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panettieri, R.A., Jr. Neutrophilic and Pauci-immune Phenotypes in Severe Asthma. Immunol. Allergy Clin. N. Am. 2016, 36, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.S.; Lee, T.H. Neutrophilic inflammation in asthma: Mechanisms and therapeutic considerations. Expert Rev. Respir. Med. 2017, 11, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Kolls, J.K. Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol. 2017, 38, 942–954. [Google Scholar] [CrossRef]
- Moore, W.C.; Hastie, A.T. Sputum neutrophil counts are associated with more severe asthma phenotypes using cluster analysis. J. Allergy Clin. Immunol. 2014, 133, 1557–1563.e5. [Google Scholar] [CrossRef]
- Simpson, J.L.; Guest, M. Occupational exposures, smoking and airway inflammation in refractory asthma. BMC Pulm. Med. 2014, 14, 207. [Google Scholar] [CrossRef]
- Carroll, N.; Carello, S. Airway structure and inflammatory cells in fatal attacks of asthma. Eur. Respir. J. 1996, 9, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Sur, S.; Crotty, T.B. Sudden-onset fatal asthma. A distinct entity with few eosinophils and relatively more neutrophils in the airway submucosa? Am. Rev. Respir. Dis. 1993, 148, 713–719. [Google Scholar] [CrossRef]
- Xiong, Y.; Cui, X. BLT1 signaling in epithelial cells mediates allergic sensitization via promotion of IL-33 production. Allergy 2019, 74, 495–506. [Google Scholar] [CrossRef]
- Gelfand, E.W.; Alam, R. The other side of asthma: Steroid-refractory disease in the absence of TH2-mediated inflammation. J. Allergy Clin. Immunol. 2015, 135, 1196–1198. [Google Scholar] [CrossRef] [PubMed]
- Medoff, B.D.; Tager, A.M. Antibody-antigen interaction in the airway drives early granulocyte recruitment through BLT1. Am. J. Physiol Lung Cell Mol. Physiol. 2006, 290, L170–L178. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, N.; Takeda, K. Leukotriene B4 receptor-1 is essential for allergen-mediated recruitment of CD8+ T cells and airway hyperresponsiveness. J. Immunol. 2005, 174, 4979–4984. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, N.; Ohnishi, H. Leukotriene B4 receptor 1 expression on dendritic cells is required for the development of Th2 responses and allergen-induced airway hyperresponsiveness. J. Immunol. 2008, 181, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, H.; Miyahara, N. Corticosteroids enhance CD8+ T cell-mediated airway hyperresponsiveness and allergic inflammation by upregulating leukotriene B4 receptor 1. J. Allergy Clin. Immunol. 2008, 121, 864–871.e4. [Google Scholar] [CrossRef]
- Gelfand, E.W.; Dakhama, A. CD8+ T lymphocytes and leukotriene B4: Novel interactions in the persistence and progression of asthma. J. Allergy Clin. Immunol. 2006, 117, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.J.; Barnes, P.J. Effect of a leukotriene B4 receptor antagonist, LY293111, on allergen induced responses in asthma. Thorax 1996, 51, 1178–1184. [Google Scholar] [CrossRef]
- Asanuma, F.; Kuwabara, K. Effects of leukotriene B4 receptor antagonist, LY293111Na, on antigen-induced bronchial hyperresponsiveness and leukocyte infiltration in sensitized guinea pigs. Inflamm. Res. 2001, 50, 136–141. [Google Scholar] [CrossRef]
- Turner, C.R.; Breslow, R. In vitro and in vivo effects of leukotriene B4 antagonism in a primate model of asthma. J. Clin. Investig. 1996, 97, 381–387. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Bjermer, L. Mast Cell-Mediated Orchestration of the Immune Responses in Human Allergic Asthma: Current Insights. Clin. Rev. Allergy Immunol. 2019, 56, 234–247. [Google Scholar] [CrossRef]
- Kouyama, S.; Otomo-Abe, A. A contraction assay system using primary cultured mouse bronchial smooth muscle cells. Int. Arch. Allergy Immunol. 2013, 161 (Suppl. 2), 93–97. [Google Scholar] [CrossRef]
- Yokomizo, T.; Nakamura, M. Leukotriene receptors as potential therapeutic targets. J. Clin. Investig. 2018, 128, 2691–2701. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.M.; Phaneuf, D.J. Short-term impact of PM2.5 on contemporaneous asthma medication use: Behavior and the value of pollution reductions. Proc. Natl. Acad. Sci. USA 2019, 116, 5246–5253. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, N.; Jones, M.J. Cysteinyl Leukotriene Receptor 1 and Health Effects of Particulate Exposure in Asthma. Ann. Am. Thorac. Soc. 2018, 15, S129. [Google Scholar] [CrossRef] [PubMed]
- Fregonese, L.; Silvestri, M. Cysteinyl leukotrienes induce human eosinophil locomotion and adhesion molecule expression via a CysLT1 receptor-mediated mechanism. Clin. Exp. Allergy 2002, 32, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Shirasaki, H.; Kanaizumi, E. Leukotriene D4 induces chemotaxis in human eosinophilc cell line, EoL-1 cells via CysLT1 receptor activation. Heliyon 2017, 3, e00464. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.C.; McKee, K. Eosinophil-eicosanoid interactions: Inhibition of eosinophil chemotaxis in vivo by a LTD4-receptor antagonist. Eur. J. Pharmacol. 1990, 191, 273–280. [Google Scholar]
- Laitinen, L.A.; Laitinen, A. Leukotriene E4 and granulocytic infiltration into asthmatic airways. Lancet 1993, 341, 989–990. [Google Scholar] [CrossRef]
- Wang, H.B.; Akuthota, P. Airway eosinophil migration into lymph nodes in mice depends on leukotriene C4. Allergy 2017, 72, 927–936. [Google Scholar] [CrossRef]
- Liu, T.; Garofalo, D. Platelet-driven leukotriene C4-mediated airway inflammation in mice is aspirin-sensitive and depends on T prostanoid receptors. J. Immunol. 2015, 194, 5061–5068. [Google Scholar] [CrossRef]
- Ilmarinen, P.; Kankaanranta, H. Eosinophil apoptosis as a therapeutic target in allergic asthma. Basic Clin. Pharmacol. Toxicol. 2014, 114, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Duah, E.; Adapala, R.K. Cysteinyl leukotrienes regulate endothelial cell inflammatory and proliferative signals through CysLT(2) and CysLT(1) receptors. Sci. Rep. 2013, 3, 3274. [Google Scholar] [CrossRef] [PubMed]
- Mauser, P.J.; House, A. Pharmacological characterization of the late phase reduction in lung functions and correlations with microvascular leakage and lung edema in allergen-challenged Brown Norway rats. Pulm. Pharmacol. Ther. 2013, 26, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, K.; Radford, K. Modulation of human airway smooth muscle migration by lipid mediators and Th-2 cytokines. Am. J. Respir. Cell Mol. Biol. 2007, 37, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Dholia, N.; Yadav, U.C.S. Lipid mediator Leukotriene D4-induces airway epithelial cells proliferation through EGFR/ERK1/2 pathway. Prostaglandins Other Lipid Mediat. 2018, 136, 55–63. [Google Scholar] [CrossRef]
- Holgate, S.T.; Peters-Golden, M. Roles of cysteinyl leukotrienes in airway inflammation, smooth muscle function, and remodeling. J. Allergy Clin. Immunol. 2003, 111, S18–S34; discussion S34–S16. [Google Scholar] [CrossRef]
- Mehrotra, A.K.; Henderson, W.R., Jr. The role of leukotrienes in airway remodeling. Curr. Mol. Med. 2009, 9, 383–391. [Google Scholar] [CrossRef]
- Matsuda, M.; Tabuchi, Y. Increased expression of CysLT2 receptors in the lung of asthmatic mice and role in allergic responses. Prostaglandins Leukot. Essent. Fatty Acids 2018, 131, 24–31. [Google Scholar] [CrossRef]
- Worrell, K.; Shaw, M.R. A systematic review of the literature on screening for exercise-induced asthma: Considerations for school nurses. J. Sch. Nurs. 2015, 31, 70–76. [Google Scholar] [CrossRef]
- Roche, A.; Ahmareen, O. The role of leukotriene receptor antagonists in exercise induced bronchoconstriction in children. Diagnosis 2014, 1, 213–222. [Google Scholar] [CrossRef]
- Tamada, T.; Ichinose, M. Leukotriene Receptor Antagonists and Antiallergy Drugs. Handb. Exp. Pharmacol. 2017, 237, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Hilberg, T.; Deigner, H.P. Transcription in response to physical stress--clues to the molecular mechanisms of exercise-induced asthma. FASEB J. 2005, 19, 1492–1494. [Google Scholar] [CrossRef] [PubMed]
- Bikov, A.; Gajdocsi, R. Exercise increases exhaled breath condensate cysteinyl leukotriene concentration in asthmatic patients. J. Asthma 2010, 47, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Arm, J.P.; Horton, C.E. Enhanced generation of leukotriene B4 by neutrophils stimulated by unopsonized zymosan and by calcium ionophore after exercise-induced asthma. Am. Rev. Respir. Dis. 1988, 138, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Li, K.L.; Lee, A.Y. Aspirin Exacerbated Respiratory Disease: Epidemiology, Pathophysiology, and Management. Med. Sci. 2019, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Rajan, J.P.; Wineinger, N.E. Prevalence of aspirin-exacerbated respiratory disease among asthmatic patients: A meta-analysis of the literature. J. Allergy Clin. Immunol. 2015, 135, 676–681.e1. [Google Scholar] [CrossRef] [PubMed]
- Sakalar, E.G.; Muluk, N.B. Aspirin-exacerbated respiratory disease and current treatment modalities. Eur. Arch. Otorhinolaryngol. 2017, 274, 1291–1300. [Google Scholar] [CrossRef]
- Kim, S.D.; Cho, K.S. Samter’s Triad: State of the Art. Clin. Exp. Otorhinolaryngol. 2018, 11, 71–80. [Google Scholar] [CrossRef]
- Morales, D.R.; Lipworth, B.J. Safety risks for patients with aspirin-exacerbated respiratory disease after acute exposure to selective nonsteroidal anti-inflammatory drugs and COX-2 inhibitors: Meta-analysis of controlled clinical trials. J. Allergy Clin. Immunol. 2014, 134, 40–45. [Google Scholar] [CrossRef]
- Steinke, J.W.; Borish, L. Factors driving the aspirin exacerbated respiratory disease phenotype. Am. J. Rhinol. Allergy 2015, 29, 35–40. [Google Scholar] [CrossRef]
- Peters-Golden, M.; Gleason, M.M. Cysteinyl leukotrienes: Multi-functional mediators in allergic rhinitis. Clin. Exp. Allergy 2006, 36, 689–703. [Google Scholar] [CrossRef] [PubMed]
- Milanovic, M.; Terszowski, G. IFN consensus sequence binding protein (Icsbp) is critical for eosinophil development. J. Immunol. 2008, 181, 5045–5053. [Google Scholar] [CrossRef] [PubMed]
- Steinke, J.W.; Liu, L. Prominent role of IFN-gamma in patients with aspirin-exacerbated respiratory disease. J. Allergy Clin. Immunol. 2013, 132, 856–865.e3. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Kanaoka, Y. Aspirin-Exacerbated Respiratory Disease Involves a Cysteinyl Leukotriene-Driven IL-33-Mediated Mast Cell Activation Pathway. J. Immunol. 2015, 195, 3537–3545. [Google Scholar] [CrossRef] [PubMed]
- Laidlaw, T.M.; Kidder, M.S. Cysteinyl leukotriene overproduction in aspirin-exacerbated respiratory disease is driven by platelet-adherent leukocytes. Blood 2012, 119, 3790–3798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayanankutty, A.; Resendiz-Hernandez, J.M. Biochemical pathogenesis of aspirin exacerbated respiratory disease (AERD). Clin. BioChem. 2013, 46, 566–578. [Google Scholar] [CrossRef]
- Laidlaw, T.M.; Boyce, J.A. Pathogenesis of aspirin-exacerbated respiratory disease and reactions. Immunol. Allergy Clin. N. Am. 2013, 33, 195–210. [Google Scholar] [CrossRef]
- Liu, T.; Barrett, N.A. Type 2 Cysteinyl Leukotriene Receptors Drive IL-33-Dependent Type 2 Immunopathology and Aspirin Sensitivity. J. Immunol. 2018, 200, 915–927. [Google Scholar] [CrossRef]
- Cingi, C.; Muluk, N.B. Antileukotrienes in upper airway inflammatory diseases. Curr. Allergy Asthma Rep. 2015, 15, 64. [Google Scholar] [CrossRef]
- Hoyte, F.C.L.; Nelson, H.S. Recent advances in allergic rhinitis. F1000Research 2018, 7. [Google Scholar] [CrossRef]
- Figueroa, D.J.; Borish, L. Expression of cysteinyl leukotriene synthetic and signalling proteins in inflammatory cells in active seasonal allergic rhinitis. Clin. Exp. Allergy 2003, 33, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Shirasaki, H.; Kanaizumi, E. Expression and localization of the cysteinyl leukotriene 1 receptor in human nasal mucosa. Clin. Exp. Allergy 2002, 32, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Suojalehto, H.; Kinaret, P. Level of Fatty Acid Binding Protein 5 (FABP5) Is Increased in Sputum of Allergic Asthmatics and Links to Airway Remodeling and Inflammation. PLoS ONE 2015, 10, e0127003. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, J. The efficacy and safety of selective H1-antihistamine versus leukotriene receptor antagonist for seasonal allergic rhinitis: A meta-analysis. PLoS ONE 2014, 9, e112815. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhou, X. Oral Antihistamines Alone vs in Combination with Leukotriene Receptor Antagonists for Allergic Rhinitis: A Meta-analysis. Otolaryngol. Head Neck Surg. 2018, 158, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Seresirikachorn, K.; Chitsuthipakorn, W. Leukotriene Receptor Antagonist Addition to H1-Antihistamine Is Effective for Treating Allergic Rhinitis: A Systematic Review and Meta-analysis. Am. J. Rhinol. Allergy 2019. [Google Scholar] [CrossRef]
- Cap, P.; Maly, M. Exhaled leukotrienes and bronchial responsiveness to methacholine in patients with seasonal allergic rhinitis. Ann. Allergy Asthma Immunol. 2009, 102, 103–109. [Google Scholar] [CrossRef]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Vakharia, P.P.; Silverberg, J.I. New and emerging therapies for paediatric atopic dermatitis. Lancet Child. Adolesc. Health 2019, 3, 343–353. [Google Scholar] [CrossRef]
- Nygaard, U.; Vestergaard, C. Emerging Treatment Options in Atopic Dermatitis: Systemic Therapies. Dermatology 2017, 233, 344–357. [Google Scholar] [CrossRef]
- Jin, H.; He, R. Animal models of atopic dermatitis. J. Investig. Dermatol. 2009, 129, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Fogh, K.; Herlin, T. Eicosanoids in skin of patients with atopic dermatitis: Prostaglandin E2 and leukotriene B4 are present in biologically active concentrations. J. Allergy Clin. Immunol. 1989, 83, 450–455. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, G. Serum metabolomics study and eicosanoid analysis of childhood atopic dermatitis based on liquid chromatography-mass spectrometry. J. Proteome Res. 2014, 13, 5715–5723. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Yasutomo, K. Treatment with DHA/EPA ameliorates atopic dermatitis-like skin disease by blocking LTB4 production. J. Med. Investig. 2016, 63, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Sadik, C.D.; Sezin, T. Leukotrienes orchestrating allergic skin inflammation. Exp. Dermatol. 2013, 22, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Oyoshi, M.K.; He, R. Leukotriene B4-driven neutrophil recruitment to the skin is essential for allergic skin inflammation. Immunity 2012, 37, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Oyoshi, M.K.; He, R. Eosinophil-derived leukotriene C4 signals via type 2 cysteinyl leukotriene receptor to promote skin fibrosis in a mouse model of atopic dermatitis. Proc. Natl. Acad. Sci. USA 2012, 109, 4992–4997. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.H.; Min, T.K. A Double-Blind, Randomized, Crossover Study to Compare the Effectiveness of Montelukast on Atopic Dermatitis in Korean Children. Allergy Asthma Immunol. Res. 2016, 8, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.Y. Is Montelukast Benefical in Children With Atopic Dermatitis? Allergy Asthma Immunol. Res. 2016, 8, 279–281. [Google Scholar] [CrossRef]
- Roekevisch, E.; Spuls, P.I. Efficacy and safety of systemic treatments for moderate-to-severe atopic dermatitis: A systematic review. J. Allergy Clin. Immunol. 2014, 133, 429–438. [Google Scholar] [CrossRef]
- Crow, D.W.; Marsella, R. Double-blinded, placebo-controlled, cross-over pilot study on the efficacy of zileuton for canine atopic dermatitis. Vet. Dermatol. 2001, 12, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Woodmansee, D.P.; Simon, R.A. A pilot study examining the role of zileuton in atopic dermatitis. Ann. Allergy Asthma Immunol. 1999, 83, 548–552. [Google Scholar] [CrossRef]
- Kupczyk, M.; Kuna, P. Targeting the PGD2/CRTH2/DP1 Signaling Pathway in Asthma and Allergic Disease: Current Status and Future Perspectives. Drugs 2017, 77, 1281–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewson, C.A.; Patel, S. Preclinical evaluation of an inhibitor of cytosolic phospholipase A2alpha for the treatment of asthma. J. Pharmacol. Exp. Ther. 2012, 340, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Yanes, D.A.; Mosser-Goldfarb, J.L. Emerging therapies for atopic dermatitis: The prostaglandin/leukotriene pathway. J. Am. Acad. Dermatol. 2018, 78, S71–S75. [Google Scholar] [CrossRef]
- Bielory, L.; Schoenberg, D. Emerging Therapeutics for Ocular Surface Disease. Curr. Allergy Asthma Rep. 2019, 19, 16. [Google Scholar] [CrossRef]
- Spada, C.S.; Woodward, D.F. Leukotrienes cause eosinophil emigration into conjunctival tissue. Prostaglandins 1986, 31, 795–809. [Google Scholar] [CrossRef]
- Andoh, T.; Sakai, K. Involvement of leukotriene B4 in itching in a mouse model of ocular allergy. Exp. Eye Res. 2012, 98, 97–103. [Google Scholar] [CrossRef]
- Pelikan, Z. Mediator profiles in tears during the conjunctival response induced by allergic reaction in the nasal mucosa. Mol. Vis. 2013, 19, 1453–1470. [Google Scholar]
- Dartt, D.A.; Hodges, R.R. Conjunctival goblet cell secretion stimulated by leukotrienes is reduced by resolvins D1 and E1 to promote resolution of inflammation. J. Immunol. 2011, 186, 4455–4466. [Google Scholar] [CrossRef]
- Lambiase, A.; Bonini, S. Montelukast, a leukotriene receptor antagonist, in vernal keratoconjunctivitis associated with asthma. Arch. Ophthalmol. 2003, 121, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Simons, F.E.; Frew, A.J. Risk assessment in anaphylaxis: Current and future approaches. J. Allergy Clin. Immunol. 2007, 120, S2–S24. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, J.H. Increased leukotriene E4 excretion in systemic mastocytosis. Prostaglandins Other Lipid Mediat. 2010, 92, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, A.; Austen, K.F. Targeted gene disruption reveals the role of cysteinyl leukotriene 1 receptor in the enhanced vascular permeability of mice undergoing acute inflammatory responses. J. Biol. Chem. 2002, 277, 20820–20824. [Google Scholar] [CrossRef] [PubMed]
- Castells, M.; Butterfield, J. Mast Cell Activation Syndrome and Mastocytosis: Initial Treatment Options and Long-Term Management. J. Allergy Clin. Immunol. Pract. 2019, 7, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Valent, P.; Akin, C. Mast cell activation syndrome: Importance of consensus criteria and call for research. J. Allergy Clin. Immunol. 2018, 142, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afrin, L.B.; Self, S. Characterization of Mast Cell Activation Syndrome. Am. J. Med. Sci. 2017, 353, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castells, M.; Austen, K.F. Mastocytosis: Mediator-related signs and symptoms. Int. Arch. Allergy Immunol. 2002, 127, 147–152. [Google Scholar] [CrossRef]
- Sala, A.; Folco, G. Transcellular biosynthesis of eicosanoids. Pharmacol. Rep. 2010, 62, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Denzlinger, C.; Haberl, C. Cysteinyl leukotriene production in anaphylactic reactions. Int. Arch. Allergy Immunol. 1995, 108, 158–164. [Google Scholar] [CrossRef]
- Tagari, P.; Rasmussen, J.B. Comparison of urinary leukotriene E4 and 16-carboxytetranordihydro leukotriene E4 excretion in allergic asthmatics after inhaled antigen. Eicosanoids 1990, 3, 75–80. [Google Scholar] [PubMed]
- Kumlin, M.; Dahlen, B. Urinary excretion of leukotriene E4 and 11-dehydro-thromboxane B2 in response to bronchial provocations with allergen, aspirin, leukotriene D4, and histamine in asthmatics. Am. Rev. Respir. Dis. 1992, 146, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.; Liu, A.H. Leukotriene-E4 in human urine: Comparison of on-line purification and liquid chromatography-tandem mass spectrometry to affinity purification followed by enzyme immunoassay. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2009, 877, 3169–3174. [Google Scholar] [CrossRef] [PubMed]
- Lueke, A.J.; Meeusen, J.W. Analytical and clinical validation of an LC-MS/MS method for urine leukotriene E4: A marker of systemic mastocytosis. Clin. BioChem. 2016, 49, 979–982. [Google Scholar] [CrossRef] [PubMed]
- Hagan, J.B.; Laidlaw, T.M. Urinary Leukotriene E4 to Determine Aspirin Intolerance in Asthma: A Systematic Review and Meta-Analysis. J. Allergy Clin. Immunol. Pract. 2017, 5, 990–997.e1. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, N.; Silveira, L. The response of children with asthma to ambient particulate is modified by tobacco smoke exposure. Am. J. Respir. Crit. Care Med. 2011, 184, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Vachier, I.; Kumlin, M. High levels of urinary leukotriene E4 excretion in steroid treated patients with severe asthma. Respir. Med. 2003, 97, 1225–1229. [Google Scholar] [CrossRef] [Green Version]
- Divekar, R.; Hagan, J. Diagnostic Utility of Urinary LTE4 in Asthma, Allergic Rhinitis, Chronic Rhinosinusitis, Nasal Polyps, and Aspirin Sensitivity. J. Allergy Clin. Immunol. Pract. 2016, 4, 665–670. [Google Scholar] [CrossRef] [Green Version]
- Marmarinos, A.; Saxoni-Papageorgiou, P. Urinary leukotriene E4 levels in atopic and non-atopic preschool children with recurrent episodic (viral) wheezing: A potential marker? J. Asthma 2015, 52, 554–559. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Tsai, M.H. Urinary LTE4 levels as a diagnostic marker for IgE-mediated asthma in preschool children: A birth cohort study. PLoS ONE 2014, 9, e115216. [Google Scholar] [CrossRef]
- Kaditis, A.G.; Alexopoulos, E. Urine concentrations of cysteinyl leukotrienes in children with obstructive sleep-disordered breathing. Chest 2009, 135, 1496–1501. [Google Scholar] [CrossRef] [PubMed]
- Satdhabudha, A.; Sritipsukho, P. Urine cysteinyl leukotriene levels in children with sleep disordered breathing before and after adenotonsillectomy. Int. J. Pediatr. Otorhinolaryngol. 2017, 94, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, N.; Graber, N.J. Urinary leukotriene E4/exhaled nitric oxide ratio and montelukast response in childhood asthma. J. Allergy Clin. Immunol. 2010, 126, 545–551.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabinovitch, N.; Mauger, D.T. Predictors of asthma control and lung function responsiveness to step 3 therapy in children with uncontrolled asthma. J. Allergy Clin. Immunol. 2014, 133, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Sunkonkit, K.; Sritippayawan, S. Urinary cysteinyl leukotriene E4 level and therapeutic response to montelukast in children with mild obstructive sleep apnea. Asian Pac. J. Allergy Immunol. 2017, 35, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.Y.; Ro, M. Mediatory roles of leukotriene B4 receptors in LPS-induced endotoxic shock. Sci. Rep. 2019, 9, 5936. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.X.; Li, K.X. Pain and bone damage in rheumatoid arthritis: Role of leukotriene B4. Clin. Exp. Rheumatol. 2019, in press. [Google Scholar]
- Zhou, J.; Lai, W. BLT1 in dendritic cells promotes Th1/Th17 differentiation and its deficiency ameliorates TNBS-induced colitis. Cell Mol. Immunol. 2018, 15, 1047–1056. [Google Scholar] [CrossRef]
- Madeira, M.F.M.; Queiroz-Junior, C.M. The role of 5-lipoxygenase in Aggregatibacter actinomycetemcomitans-induced alveolar bone loss. J. Clin. Periodontol. 2017, 44, 793–802. [Google Scholar] [CrossRef]
- Hikiji, H.; Ishii, S. A distinctive role of the leukotriene B4 receptor BLT1 in osteoclastic activity during bone loss. Proc. Natl. Acad. Sci. USA 2009, 106, 21294–21299. [Google Scholar] [CrossRef]
- Sezin, T.; Krajewski, M. The Leukotriene B4 and its Receptor BLT1 Act as Critical Drivers of Neutrophil Recruitment in Murine Bullous Pemphigoid-Like Epidermolysis Bullosa Acquisita. J. Investig. Dermatol. 2017, 137, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Toda, A.; Terawaki, K. Attenuated Th1 induction by dendritic cells from mice deficient in the leukotriene B4 receptor 1. Biochimie 2010, 92, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Zou, L. Leukotriene B(4)-leukotriene B(4) receptor axis promotes oxazolone-induced contact dermatitis by directing skin homing of neutrophils and CD8(+) T cells. Immunology 2015, 146, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Sumida, H.; Yanagida, K. Interplay between CXCR2 and BLT1 facilitates neutrophil infiltration and resultant keratinocyte activation in a murine model of imiquimod-induced psoriasis. J. Immunol. 2014, 192, 4361–4369. [Google Scholar] [CrossRef] [PubMed]
- Hegde, B.; Bodduluri, S.R. Inflammasome-Independent Leukotriene B4 Production Drives Crystalline Silica-Induced Sterile Inflammation. J. Immunol. 2018, 200, 3556–3567. [Google Scholar] [CrossRef] [PubMed]
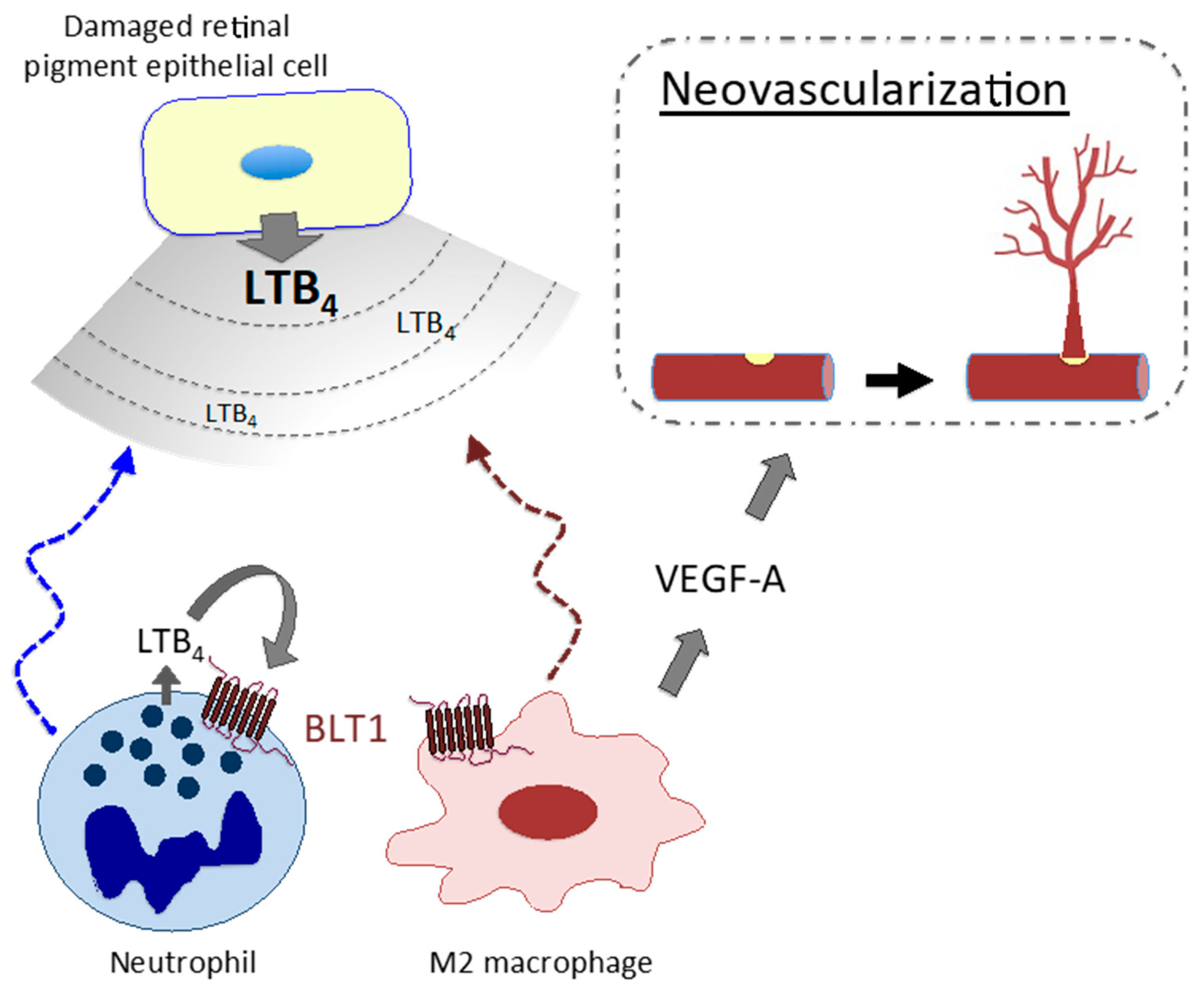
- Sasaki, F.; Koga, T. Leukotriene B4 promotes neovascularization and macrophage recruitment in murine wet-type AMD models. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Disease | Roles of the LTB4-BLT1 Pathway | Roles of CysLT Pathways | Anti-leukotriene Standard Therapy in a Clinical Trial Experimentally Effective |
|---|---|---|---|
| Asthma |
|
| Leukotriene receptor antagonist (LTRA) 5-LO inhibitor (zileuton) |
| Neutrophilic asthma |
| BLT1 antagonist (LY293111, CP-105,696) | |
| Aspirin-exacerbated respiratory disease; aspirin-sensitive asthma |
| ||
| Allergic rhinitis |
| LTRA | |
| Atopic dermatitis |
|
| Q301 (zileuton cream)? ZPL-521 (cPLA2 inhibitor ointment)? 5-LO inhibitor (zileuton) |
| Allergic conjunctivitis |
|
| LTRA (montelukast) BLT1 antagonist (ONO-4057) 5-LO inhibitor (zileuton) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo-Watanabe, A.; Okuno, T.; Yokomizo, T. The Role of Leukotrienes as Potential Therapeutic Targets in Allergic Disorders. Int. J. Mol. Sci. 2019, 20, 3580. https://doi.org/10.3390/ijms20143580
Jo-Watanabe A, Okuno T, Yokomizo T. The Role of Leukotrienes as Potential Therapeutic Targets in Allergic Disorders. International Journal of Molecular Sciences. 2019; 20(14):3580. https://doi.org/10.3390/ijms20143580
Chicago/Turabian StyleJo-Watanabe, Airi, Toshiaki Okuno, and Takehiko Yokomizo. 2019. "The Role of Leukotrienes as Potential Therapeutic Targets in Allergic Disorders" International Journal of Molecular Sciences 20, no. 14: 3580. https://doi.org/10.3390/ijms20143580
APA StyleJo-Watanabe, A., Okuno, T., & Yokomizo, T. (2019). The Role of Leukotrienes as Potential Therapeutic Targets in Allergic Disorders. International Journal of Molecular Sciences, 20(14), 3580. https://doi.org/10.3390/ijms20143580