The Antitumor Activity of TCR-Mimic Antibody-Drug Conjugates (TCRm-ADCs) Targeting the Intracellular Wilms Tumor 1 (WT1) Oncoprotein

 , and
, and
Abstract
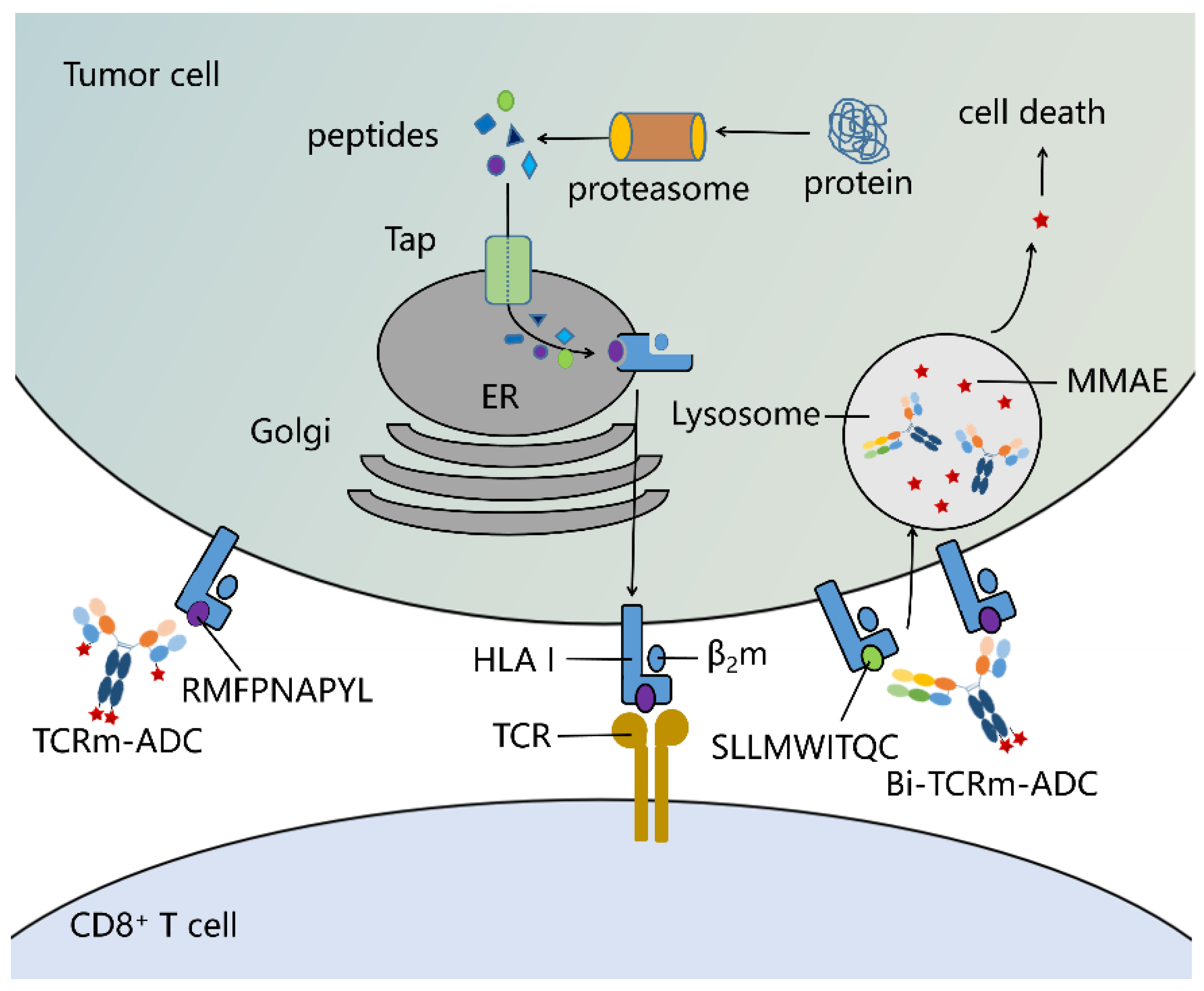
:1. Introduction
2. Results
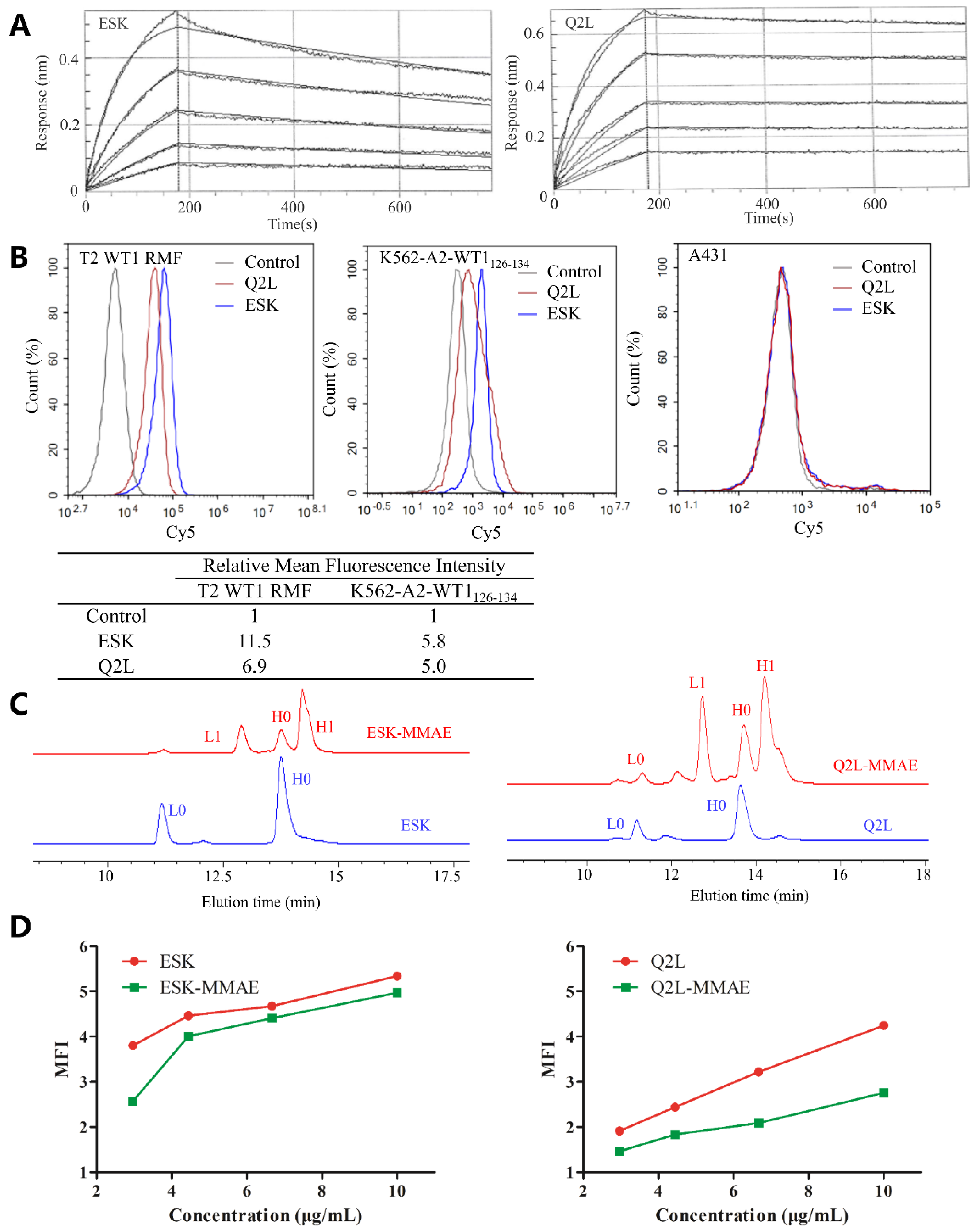
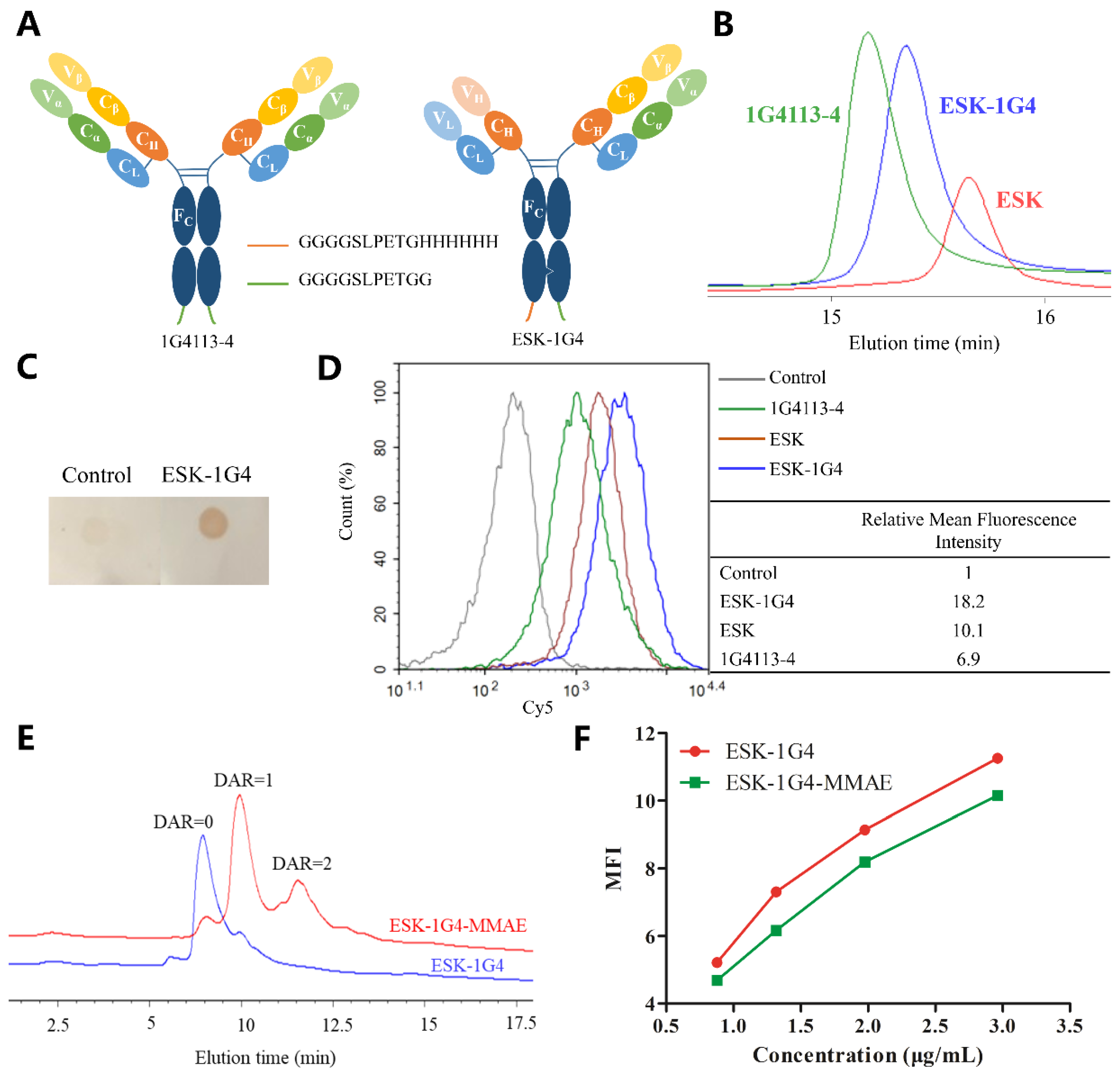
2.1. Preparation and Characterization of TCRm Antibodies
2.2. Sortase A Generated Site-Specific Conjugated TCRm-ADCs and Their Characterizations
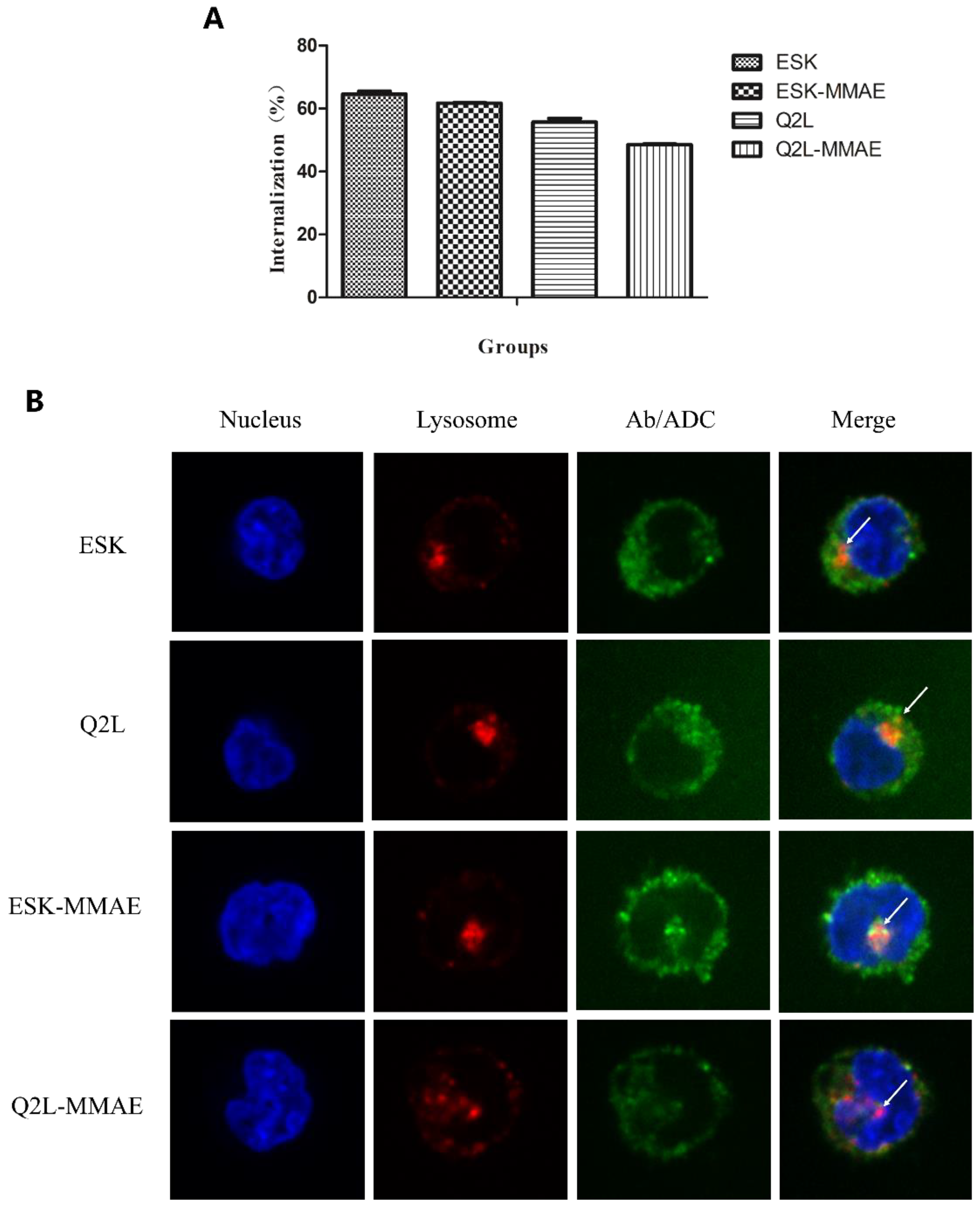
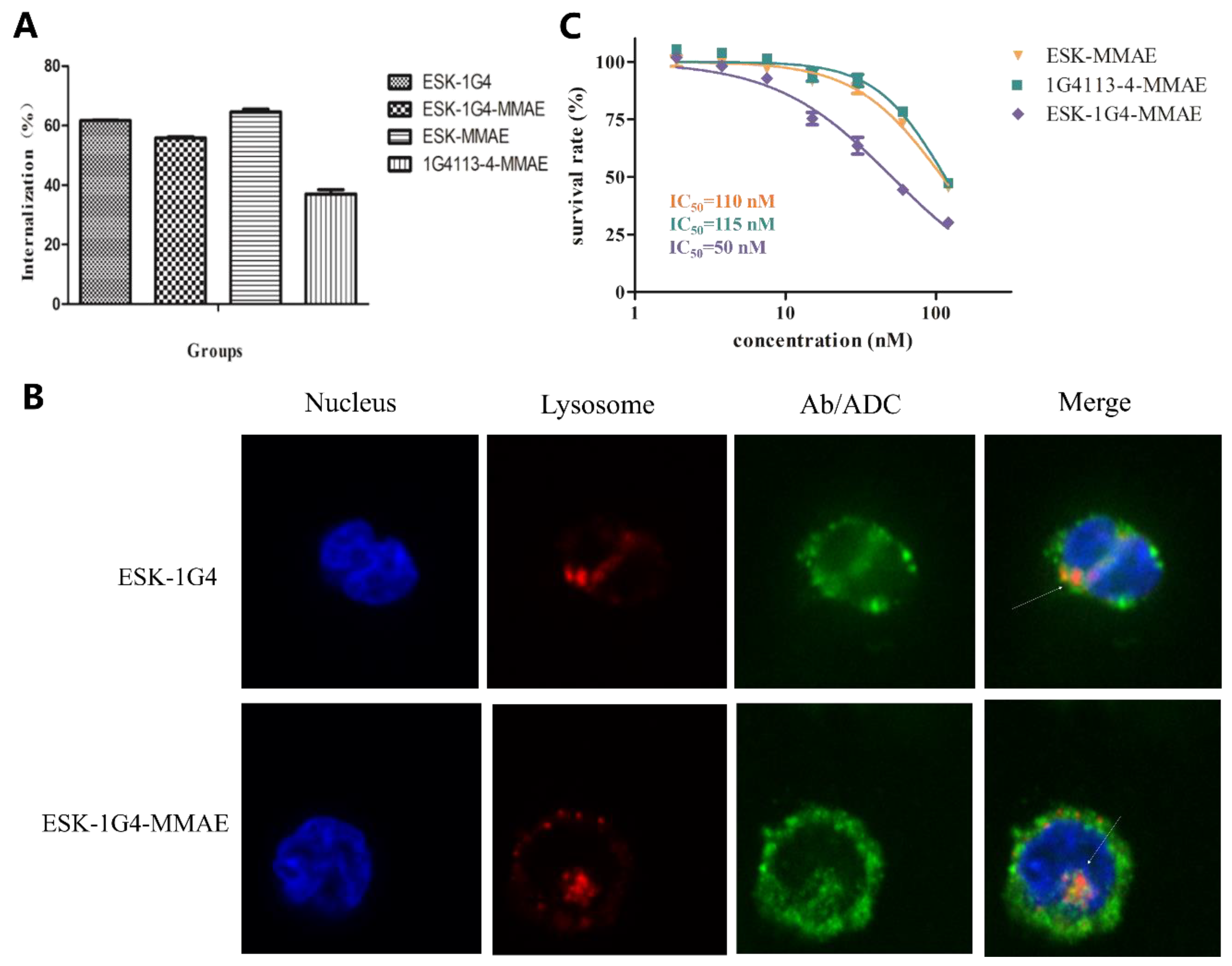
2.3. Internalization of TCRm Antibodies and Their Conjugates
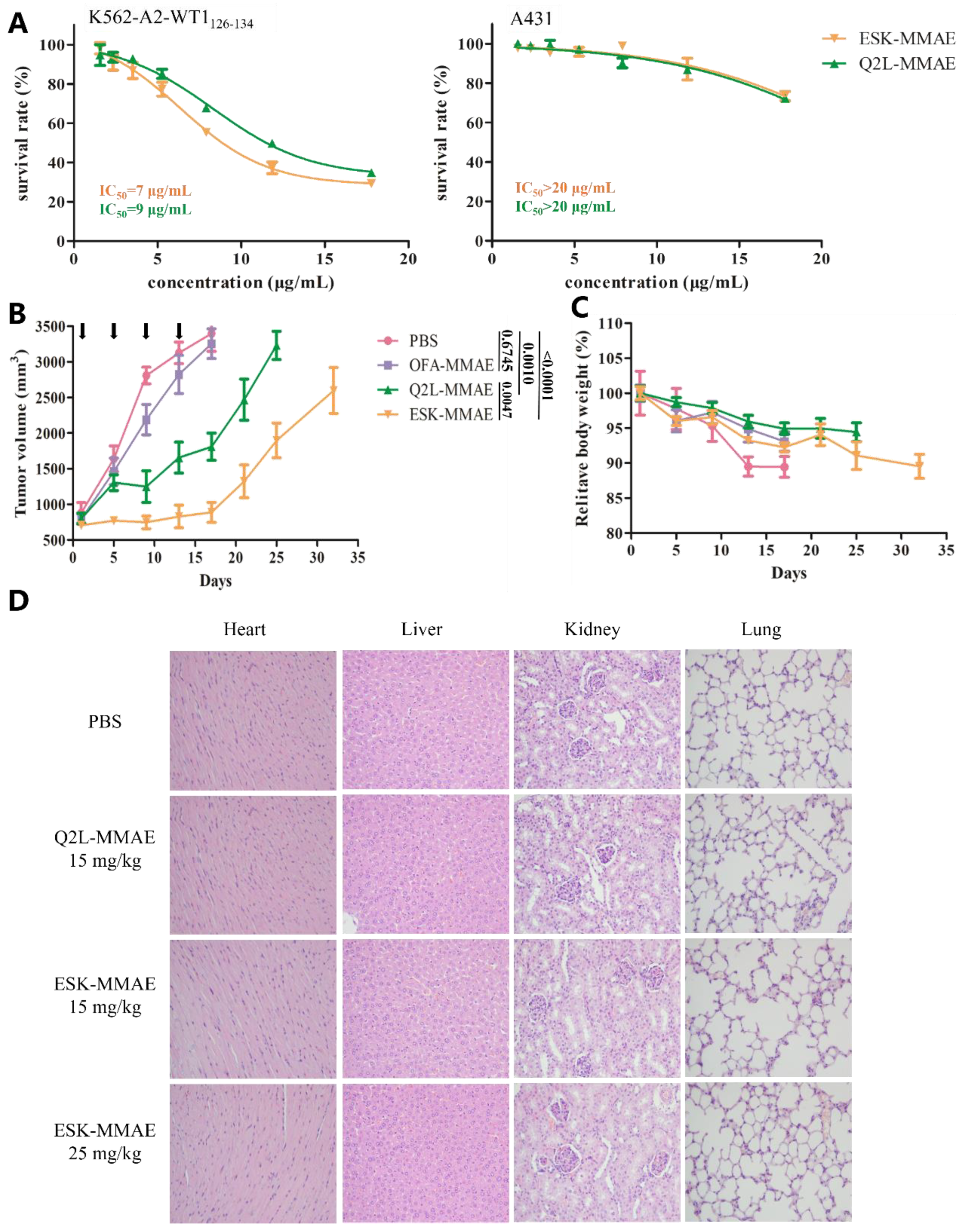
2.4. In Vitro and In Vivo Antitumor Activity of TCRm-ADCs
2.5. Preparation and Characterization of Bi-TCRm-ADC
2.6. Internalization and In Vitro Cytotoxicity of Bi-TCRm-ADC
2.7. Prediction of WT1 Protein Epitope Peptides
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Antibodies and Sortase A Enzyme
4.3. Surface Plasmon Resonance (SPR) Analysis
4.4. Dot-ELISA for Verifying the Expression of ESK-1G4
4.5. Sortase A-Mediated Conjugation of Antibodies with Gly3-Val-Cit-PAB-MMAE
4.6. Characterization of ADCs
4.7. Flow Cytometry Analysis for Cell-Binding Assays
4.7.1. Binding of TCRm Antibodies on Different Cell Lines
4.7.2. Influence of Binding Affinity of ESK, Q2L, and ESK-1G4 after Sortase A-Mediated Conjugation on K562-A2-WT1126–134 Cell Line
4.8. Cellular Internalization
4.8.1. Cellular Internalization Ratio of TCRm Antibodies and Their Conjugates
4.8.2. Microscopy for TCRm Antibodies and Conjugates Trafficking
4.9. In Vitro Efficacy
4.10. Mouse Xenograft Study
4.11. Prediction of WT1 Protein Epitope Peptides
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- de Goeij, B.E.; Lambert, J.M. New developments for antibody-drug conjugate-based therapeutic approaches. Curr. Opin. Immunol. 2016, 40, 14–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner, L.M.; Murray, J.C.; Shuptrine, C.W. Antibody-based immunotherapy of cancer: New insights, new targets. Cell 2012, 148, 1081–1084. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.M.; Wolchok, J.D.; Old, L.J. Antibody therapy of cancer. Nat. Rev. Cancer 2012, 12, 278–287. [Google Scholar] [CrossRef] [PubMed]
- York, I.A.; Rock, K.L. Antigen processing and presentation by the class I major histocompatibility complex. Annu. Rev. Immunol. 1996, 14, 369–396. [Google Scholar] [CrossRef] [PubMed]
- Vyas, J.M.; Van Der Veen, A.G.; Ploegh, H.L. The known unknowns of antigen processing and presentation. Nat. Rev. Immunol. 2008, 8, 607–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epel, M.; Carmi, I.; Soueid-Baumgarten, S.; Oh, S.K.; Bera, T.; Pastan, I.; Berzofsky, J.; Reiter, Y. Targeting TARP, a novel breast and prostate tumor-associated antigen, with T cell receptor-like human recombinant antibodies. Eur. J. Immunol. 2008, 38, 1706–1720. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Rawat, A.; Verma, B.; Markiewski, M.M.; Weidanz, J.A. Antitumor activity of a monoclonal antibody targeting major histocompatibility complex class I-Her2 peptide complexes. J. Natl. Cancer Inst. 2013, 105, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.; Yan, S.; Veomett, N.; Pankov, D.; Zhou, L.; Korontsvit, T.; Scott, A.; Whitten, J.; Maslak, P.; Casey, E.; et al. Targeting the intracellular WT1 oncogene product with a therapeutic human antibody. Sci. Transl. Med. 2013, 5, 176ra33. [Google Scholar] [CrossRef]
- Veomett, N.; Dao, T.; Liu, H.; Xiang, J.; Pankov, D.; Dubrovsky, L.; Whitten, J.A.; Park, S.M.; Korontsvit, T.; Zakhaleva, V.; et al. Therapeutic efficacy of an Fc-enhanced TCR-like antibody to the intracellular WT1 oncoprotein. Clin. Cancer Res. 2014, 20, 4036–4046. [Google Scholar] [CrossRef]
- Chang, A.Y.; Dao, T.; Gejman, R.S.; Jarvis, C.A.; Scott, A.; Dubrovsky, L.; Mathias, M.D.; Korontsvit, T.; Zakhaleva, V.; Curcio, M.; et al. A therapeutic T cell receptor mimic antibody targets tumor-associated PRAME peptide/HLA-I antigens. J. Clin. Invest. 2017, 127, 2705–2718. [Google Scholar] [CrossRef]
- Lai, J.; Wang, Y.; Wu, S.S.; Ding, D.; Sun, Z.Y.; Zhang, Y.; Zhou, J.; Zhou, Z.; Xu, Y.C.; Pan, L.Q.; et al. Elimination of melanoma by sortase A-generated TCR-like antibody-drug conjugates (TL-ADCs) targeting intracellular melanoma antigen MART-1. Biomaterials 2018, 178, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Lowe, D.B.; Bivens, C.K.; Mobley, A.S.; Herrera, C.E.; McCormick, A.L.; Wichner, T.; Sabnani, M.K.; Wood, L.M.; Weidanz, J.A. TCR-like antibody drug conjugates mediate killing of tumor cells with low peptide/HLA targets. MAbs 2017, 9, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keilholz, U.; Menssen, H.D.; Gaiger, A.; Menke, A.; Oji, Y.; Oka, Y.; Scheibenbogen, C.; Stauss, H.; Thiel, E.; Sugiyama, H. Wilms’ tumour gene 1 (WT1) in human neoplasia. Leukemia 2005, 19, 1318–1323. [Google Scholar] [CrossRef] [PubMed]
- Cheever, M.A.; Allison, J.P.; Ferris, A.S.; Finn, O.J.; Hastings, B.M.; Hecht, T.T.; Mellman, I.; Prindiville, S.A.; Viner, J.L.; Weiner, L.M.; et al. The prioritization of cancer antigens: A National Cancer Institute pilot project for the acceleration of translational research. Clin. Cancer Res. 2009, 15, 5323–5337. [Google Scholar] [CrossRef] [PubMed]
- Keilholz, U.; Letsch, A.; Busse, A.; Asemissen, A.M.; Bauer, S.; Blau, I.W.; Hofmann, W.-K.; Uharek, L.; Thiel, E.; Scheibenbogen, C. A clinical and immunologic phase 2 trial of Wilms tumor gene product 1 (WT1) peptide vaccination in patients with AML and MDS. Blood 2009, 113, 6541–6548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezvani, K.; Yong, A.S.M.; Mielke, S.; Savani, B.N.; Musse, L.; Superata, J.; Jafarpour, B.; Boss, C.; Barrett, A.J. Leukemia-associated antigen-specific T-cell responses following combined PR1 and WT1 peptide vaccination in patients with myeloid malignancies. Blood 2008, 111, 236–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krug, L.M.; Dao, T.; Brown, A.B.; Maslak, P.; Travis, W.; Bekele, S.; Korontsvit, T.; Zakhaleva, V.; Wolchok, J.; Yuan, J.; et al. WT1 peptide vaccinations induce CD4 and CD8 T cell immune responses in patients with mesothelioma and non-small cell lung cancer. Cancer Immunol. Immunother. 2010, 59, 1467–1479. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Ahmed, M.; Tassev, D.V.; Hasan, A.; Kuo, T.-Y.; Guo, H.-F.; O’Reilly, R.J.; Cheung, N.-K.V. Affinity maturation of T-cell receptor-like antibodies for Wilms tumor 1 peptide greatly enhances therapeutic potential. Leukemia 2015, 29, 2238–2247. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, S.; Purdon, T.J.; Daniyan, A.F.; Koneru, M.; Dao, T.; Liu, C.; Scheinberg, D.A.; Brentjens, R.J. Optimized T-cell receptor-mimic chimeric antigen receptor T cells directed toward the intracellular Wilms Tumor 1 antigen. Leukemia 2017, 31, 1788–1797. [Google Scholar] [CrossRef]
- Akcakanat, A.; Kanda, T.; Koyama, Y.; Watanabe, M.; Kimura, E.; Yoshida, Y.; Komukai, S.; Nakagawa, S.; Odani, S.; Fujii, H.; et al. NY-ESO-1 expression and its serum immunoreactivity in esophageal cancer. Cancer Chemother. Pharm. 2004, 54, 95–100. [Google Scholar] [CrossRef]
- Prasad, M.L.; Jungbluth, A.A.; Patel, S.G.; Iversen, K.; Hoshaw-Woodard, S.; Busam, K.J. Expression and significance of cancer testis antigens in primary mucosal melanoma of the head and neck. Head Neck 2004, 26, 1053–1057. [Google Scholar] [CrossRef]
- Thomas, R.; Al-Khadairi, G.; Roelands, J.; Hendrickx, W.; Dermime, S.; Bedognetti, D.; Decock, J. NY-ESO-1 based immunotherapy of cancer: Current perspectives. Front. Immunol. 2018, 9, 947. [Google Scholar] [CrossRef]
- McCormack, E.; Adams, K.J.; Hassan, N.J.; Kotian, A.; Lissin, N.M.; Sami, M.; Mujić, M.; Osdal, T.; Gjertsen, B.T.; Baker, D.; et al. Bi-specific TCR-anti CD3 redirected T-cell targeting of NY-ESO-1- and LAGE-1-positive tumors. Cancer Immunol. Immunother. 2013, 62, 773–785. [Google Scholar] [CrossRef]
- Salter, R.D.; Howell, D.N.; Cresswell, P. Genes regulating HLA class I antigen expression in T-B lymphoblast hybrids. Immunogenetics 1985, 21, 235–246. [Google Scholar] [CrossRef]
- Brickner, A.G. The PANE1 gene encodes a novel human minor histocompatibility antigen that is selectively expressed in B-lymphoid cells and B-CLL. Blood 2006, 107, 3779–3786. [Google Scholar] [CrossRef]
- Li, Z.H.; Zhang, Q.; Wang, H.B.; Zhang, Y.N.; Ding, D.; Pan, L.Q.; Miao, D.; Xu, S.; Zhang, C.; Luo, P.H.; et al. Preclinical studies of targeted therapies for CD20-positive B lymphoid malignancies by Ofatumumab conjugated with auristatin. Invest. New Drugs 2014, 32, 75–86. [Google Scholar] [CrossRef]
- Xu, Y.; Jin, S.; Zhao, W.; Liu, W.; Ding, D.; Zhou, J.; Chen, S. A versatile chemo-enzymatic conjugation approach yields homogeneous and highly potent antibody-drug conjugates. Int. J. Mol. Sci. 2017, 18, 2284. [Google Scholar] [CrossRef]
- Zhao, W. Study on the Factors of Safety and Efficacy of Novel Antibody-Drug Conjugates. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2019. [Google Scholar]
- Pan, L.; Zhao, W.; Lai, J.; Ding, D.; Zhang, Q.; Yang, X.; Huang, M.; Jin, S.; Xu, Y.; Zeng, S.; et al. Sortase A-generated highly potent anti-CD20-MMAE conjugates for efficient elimination of B-lineage lymphomas. Small 2017, 13, 1–12. [Google Scholar] [CrossRef]
- Wakankar, A.; Chen, Y.; Gokarn, Y.; Jacobson, F.S. Analytical methods for physicochemical characterization of antibody drug conjugates. MAbs 2011, 3, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Zuckier, L.S.; Berkowitz, E.Z.; Sattenberg, R.J.; Zhao, Q.H.; Deng, H.F.; Scharff, M.D. Influence of affinity and antigen density on antibody localization in a modifiable tumor targeting model. Cancer Res. 2000, 60, 7008–7013. [Google Scholar]
- Zhou, F. Molecular mechanisms of IFN-γ to up-regulate MHC class I antigen processing and presentation. Int. Rev. Immunol. 2009, 28, 239–260. [Google Scholar] [CrossRef]
- Yang, Y.M.; Shang, D.S.; Zhao, W.D.; Fang, W.G.; Chen, Y.H. Microglial TNF-α-dependent elevation of MHC class I expression on brain endothelium induced by amyloid-beta promotes T cell transendothelial migration. Neurochem. Res. 2013, 38, 2295–2304. [Google Scholar] [CrossRef]
- Jäger, E.; Chen, Y.-T.; Drijfhout, J.W.; Karbach, J.; Ringhoffer, M.; Jäger, D.; Arand, M.; Wada, H.; Noguchi, Y.; Stockert, E.; et al. Simultaneous humoral and cellular immune response against cancer–testis antigen NY-ESO-1: Definition of human histocompatibility leukocyte antigen (HLA)-A2–binding peptide epitopes. J. Exp. Med. 2002, 187, 265–270. [Google Scholar] [CrossRef]
- Ledsgaard, L.; Kilstrup, M.; Karatt-Vellatt, A.; McCafferty, J.; Laustsen, A.H. Basics of antibody phage display technology. Toxins 2018, 10, 236. [Google Scholar] [CrossRef]
- Tillotson, B.J.; Lajoie, J.M.; Shusta, E.V. Yeast display-based antibody affinity maturation using detergent-solubilized cell lysates. Methods Mol. Biol. 2015, 1319, 65–78. [Google Scholar]
- Brinkmann, U.; Kontermann, R.E. The making of bispecific antibodies. MAbs 2017, 9, 182–212. [Google Scholar] [CrossRef]
- Runcie, K.; Budman, D.R.; John, V.; Seetharamu, N. Bi-specific and tri-specific antibodies- the next big thing in solid tumor therapeutics. Mol. Med. 2018, 24, 50. [Google Scholar] [CrossRef]
- Steinhardt, J.J.; Guenaga, J.; Turner, H.L.; McKee, K.; Louder, M.K.; O’Dell, S.; Chiang, C.I.; Lei, L.; Galkin, A.; Andrianov, A.K.; et al. Rational design of a trispecific antibody targeting the HIV-1 Env with elevated anti-viral activity. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Skora, A.D.; Douglass, J.; Hwang, M.S.; Tam, A.J.; Blosser, R.L.; Gabelli, S.B.; Cao, J.; Diaz, L.A.; Papadopoulos, N.; Kinzler, K.W.; et al. Generation of MANAbodies specific to HLA-restricted epitopes encoded by somatically mutated genes. Proc. Natl. Acad. Sci. 2015, 112, 9967–9972. [Google Scholar] [CrossRef] [Green Version]
- Beerli, R.R.; Hell, T.; Merkel, A.S.; Grawunder, U. Sortase enzyme-mediated generation of site-specifically conjugated antibody drug conjugates with high in vitro and in vivo potency. PloS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Li, Y.; Moysey, R.; Molloy, P.E.; Vuidepot, A.L.; Mahon, T.; Baston, E.; Dunn, S.; Liddy, N.; Jacob, J.; Jakobsen, B.K.; et al. Directed evolution of human T-cell receptors with picomolar affinities by phage display. Nat. Biotechnol. 2005, 23, 349–354. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Y.; Zhang, Y.; Li, J.; Tsao, S.W.; Zhang, M.-Y. Superior antitumor activity of a novel bispecific antibody cotargeting human epidermal growth factor receptor 2 and type I insulin-like growth factor receptor. Mol. Cancer 2013, 13, 90–100. [Google Scholar] [CrossRef]
- Yang, D.; Singh, A.; Wu, H.; Kroe-Barrett, R. Dataset of the binding kinetic rate constants of anti-PCSK9 antibodies obtained using the Biacore T100, ProteOn XPR36, Octet RED384, and IBIS MX96 biosensor platforms. Data Br. 2016, 8, 1173–1183. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TCRm Antibodies | IgG Isotype | Ka (1/Ms) | Kd (1/s) | KD (nM) |
|---|---|---|---|---|
| ESK | Hu IgG1 | 1.64 × 105 | 5.73 × 10−4 | 3.50 |
| Q2L | Hu IgG1 | 9.96 × 103 | 9.03 × 10−5 | 9.09 |
| Peptide Length | Position | Peptide Sequence | NetCTL | NetMHC | |
|---|---|---|---|---|---|
| Cleavage | Tap | Affinity(nM) | |||
| 8 | 8–15 | LNALLPAV | 0.482568 | NK | 274.80 |
| 9 | 10–18 | ALLPAVPSL | 0.028869 | 1.2770 | 6.01 |
| 126–134 | RMFPNAPYL | 0.244498 | 1.5860 | 7.14 | |
| 187–195 | SLGEQQYSV | 0.143282 | 0.1940 | 21.76 | |
| 191–200 | QQYSVPPPV | 0.044789 | 0.4230 | 29.84 | |
| 225–233 | NLYQMTSQL | 0.090656 | 1.0590 | 236.39 | |
| 280–288 | ILCGAQYRI | 0.308379 | 0.4410 | 128.47 | |
| 10 | 6–15 | RDLNALLPAV | 0.093866 | NK | 33.75 |
| 9–18 | RMFPNAPYLP | 0.802949 | NK | 173.54 | |
| 125–134 | ARMFPNAPYL | 0.297255 | NK | 28.46 | |
| 126–135 | ALLPAVPSLG | 0.035614 | NK | 223.13 | |
| 11 | 5–15 | VRDLNALLPAV | 0.018917 | NK | 285.66 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Li, Y.-M.; Zhou, J.-J.; Zhou, Z.; Xu, Y.-C.; Zhao, W.-B.; Chen, S.-Q. The Antitumor Activity of TCR-Mimic Antibody-Drug Conjugates (TCRm-ADCs) Targeting the Intracellular Wilms Tumor 1 (WT1) Oncoprotein. Int. J. Mol. Sci. 2019, 20, 3912. https://doi.org/10.3390/ijms20163912
Shen Y, Li Y-M, Zhou J-J, Zhou Z, Xu Y-C, Zhao W-B, Chen S-Q. The Antitumor Activity of TCR-Mimic Antibody-Drug Conjugates (TCRm-ADCs) Targeting the Intracellular Wilms Tumor 1 (WT1) Oncoprotein. International Journal of Molecular Sciences. 2019; 20(16):3912. https://doi.org/10.3390/ijms20163912
Chicago/Turabian StyleShen, Ying, Yi-Ming Li, Jing-Jing Zhou, Zhan Zhou, Ying-Chun Xu, Wen-Bin Zhao, and Shu-Qing Chen. 2019. "The Antitumor Activity of TCR-Mimic Antibody-Drug Conjugates (TCRm-ADCs) Targeting the Intracellular Wilms Tumor 1 (WT1) Oncoprotein" International Journal of Molecular Sciences 20, no. 16: 3912. https://doi.org/10.3390/ijms20163912
APA StyleShen, Y., Li, Y. -M., Zhou, J. -J., Zhou, Z., Xu, Y. -C., Zhao, W. -B., & Chen, S. -Q. (2019). The Antitumor Activity of TCR-Mimic Antibody-Drug Conjugates (TCRm-ADCs) Targeting the Intracellular Wilms Tumor 1 (WT1) Oncoprotein. International Journal of Molecular Sciences, 20(16), 3912. https://doi.org/10.3390/ijms20163912