YaeB, Expressed in Response to the Acidic pH in Macrophages, Promotes Intracellular Replication and Virulence of Salmonella Typhimurium


Abstract
:1. Introduction
2. Results
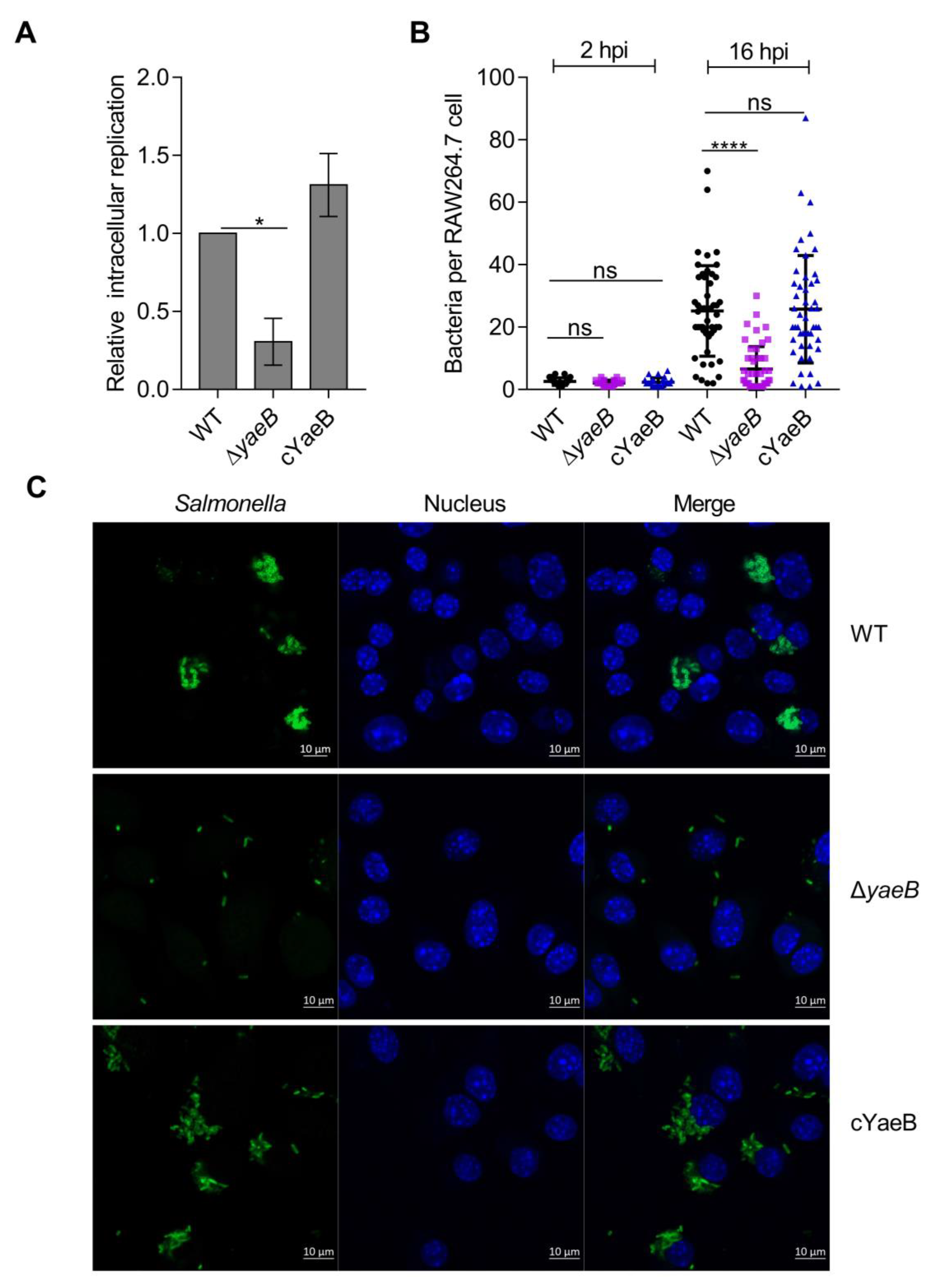
2.1. Deletion of YaeB Impaired S. Typhimurium Growth in Macrophages
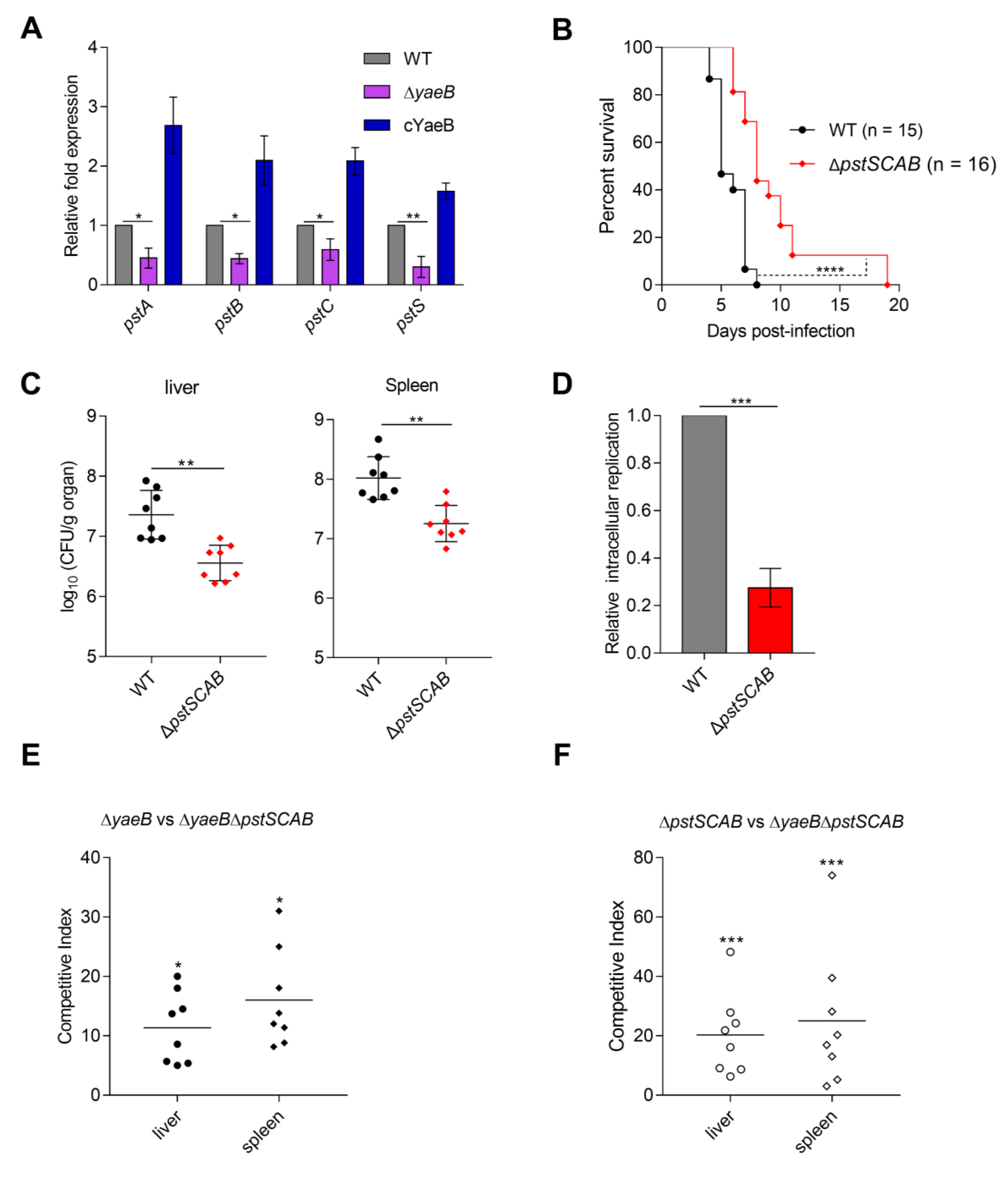
2.2. Deletion of YaeB Impaired S. Typhimurium Virulence in Mice
2.3. Deletion of YaeB Reduced the Expression of the pstSCAB Gene
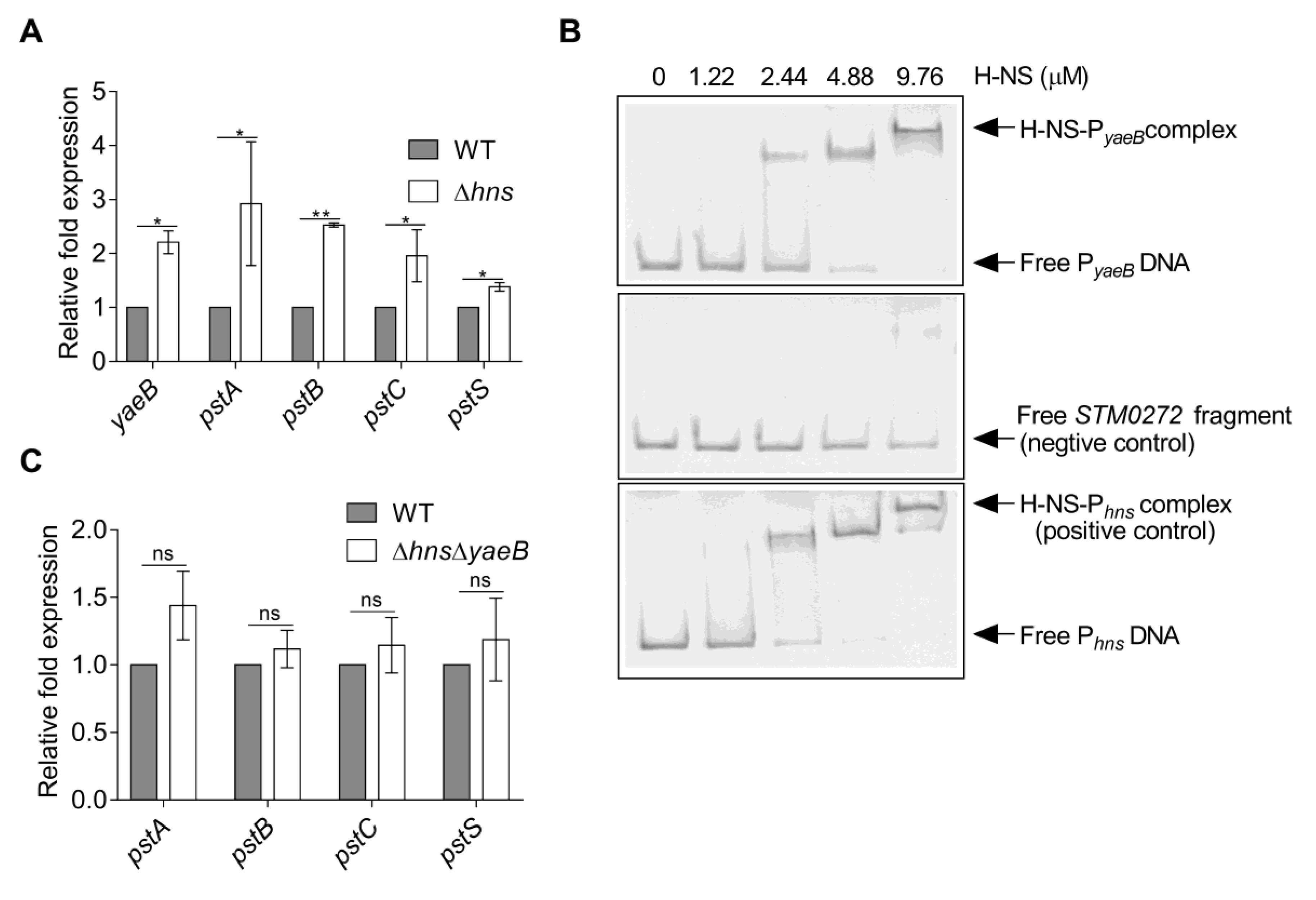
2.4. YaeB was Directly Repressed by H-NS
2.5. YaeB Expression was Activated by an Acidic pH
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Bacterial Strains and Plasmids
4.3. Bacterial Growth Culture and Cell Culture
4.4. Macrophage Replication Assays
4.5. Immunofluorescence Microscopy
4.6. Mouse Infections
4.7. RNA-Seq
4.8. qRT-PCR Analysis
4.9. EMSAs
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SCV | Salmonella-containing vacuole |
| qRT-PCR | quantitative real-time PCR |
| CFU | Colony-Forming Units |
| FPKM | fragments per kilobase of transcript per million fragments mapped |
References
- Gerlach, R.G.; Claudio, N.; Rohde, M.; Jackel, D.; Wagner, C.; Hensel, M. Cooperation of Salmonella pathogenicity islands 1 and 4 is required to breach epithelial barriers. Cell Microbiol. 2008, 11, 2364–2376. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Torres, A.; Fang, F.C. Cellular routes of invasion by enteropathogens. Curr. Opin. Microbiol. 2000, 1, 54–59. [Google Scholar] [CrossRef]
- Dandekar, T.; Astrid, F.; Jasmin, P.; Hensel, M. Salmonella enterica: A surprisingly well-adapted intracellular lifestyle. Front. Microbiol. 2012, 3, 164. [Google Scholar] [CrossRef] [PubMed]
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 1, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Beuzon, C.R.; Banks, G.; Deiwick, J.; Hensel, M.; Holden, D.W. PH-dependent secretion of SseB, a product of the SPI-2 type III secretion system of Salmonella typhimurium. Mol. Microbiol. 1999, 4, 806–816. [Google Scholar] [CrossRef]
- Jennings, E.; Thurston, T.L.M.; Holden, D.W. Salmonella SPI-2 type III secretion system effectors: Molecular mechanisms and physiological consequences. Cell Host Microbe 2017, 2, 217–231. [Google Scholar] [CrossRef]
- Deiwick, J.; Nikolaus, T.; Erdogan, S.; Hensel, M. Environmental regulation of Salmonella pathogenicity island 2 gene expression. Mol. Microbiol. 1999, 6, 1759–1773. [Google Scholar] [CrossRef] [PubMed]
- Garmendia, J.; Beuzon, C.R.; Ruiz-Albert, J.; Holden, D.W. The roles of SsrA-SsrB and OmpR-EnvZ in the regulation of genes encoding the Salmonella typhimurium SPI-2 type III secretion system. Microbiology 2003, 149, 2385–2396. [Google Scholar] [CrossRef]
- Holtkötter, P.; Hensel, M. Metabolism of Intracellular Salmonella enterica. In Host-pathogen interaction: Microbial metabolism, pathogenicity and antiinfectives; Unden, A., Thines, E., Eds.; Wiley: Hoboken, NY, USA, 2016; pp. 37–55. [Google Scholar]
- Parkinson, J.S. Signal transduction schemes of bacteria. Cell 1993, 5, 857–871. [Google Scholar] [CrossRef]
- Chekabab, S.M.; Harel, J.; Dozois, C.M. Interplay between genetic regulation of phosphate homeostasis and bacterial virulence. Virulence 2014, 8, 786–793. [Google Scholar] [CrossRef]
- Kimura, S.; Miyauchi, K.; Ikeuchi, Y.; Thiaville, P.C.; Crecy-Lagard, V.; Suzuki, T. Discovery of the beta-barrel-type RNA methyltransferase responsible for N6-methylation of N6-threonylcarbamoyladenosine in tRNAs. Nucleic Acids Res. 2014, 14, 9350–9365. [Google Scholar] [CrossRef] [PubMed]
- Srikumar, S.; Kröger, C.; Hébrard, M.; Colgan, A.; Owen, S.V.; Sivasankaran, S.K.; Cameron, A.D.S.; Hokamp, K.; Hinton, J.C.D. RNA-seq brings new insights to the Intra-Macrophage transcriptome of salmonella typhimurium. Plos Pathog. 2015, 11, e1005262. [Google Scholar] [CrossRef] [PubMed]
- Guadarrama, C.; Medrano-Lopez, A.; Oropeza, R.; Hernandez-Lucas, I.; Calva, E. The Salmonella enterica serovar Typhi LeuO global regulator forms tetramers: Residues involved in oligomerization, DNA binding, and transcriptional regulation. J. Bacteriol. 2014, 12, 2143–2154. [Google Scholar] [CrossRef] [PubMed]
- Saier, M.J.; Wentzel, D.L.; Feucht, B.U.; Judice, J.J. A transport system for phosphoenolpyruvate, 2-phosphoglycerate, and 3-phosphoglycerate in Salmonella typhimurium. J. Biol. Chem. 1975, 13, 5089–5096. [Google Scholar]
- Niu, S.; Jiang, S.Q.; Hong, J. Salmonella typhimurium pgtB mutants conferring constitutive expression of phosphoglycerate transporter pgtP independent of pgtC. J. Bacteriol. 1995, 15, 4297–4302. [Google Scholar] [CrossRef]
- Steeb, B.; Claudi, B.; Burton, N.A.; Tienz, P.; Schmidt, A.; Farhan, H.; Mazé, A.; Bumann, D. Parallel exploitation of diverse host nutrients enhances Salmonella virulence. Plos Pathog. 2013, 4, e1003301. [Google Scholar] [CrossRef] [PubMed]
- Hengge, R.; Larson, T.J.; Boos, W. Sn-Glycerol-3-phosphate transport in Salmonella typhimurium. J. Bacteriol. 1983, 1, 186–195. [Google Scholar]
- Argast, M.; Ludtke, D.; Silhavy, T.J.; Boos, W. A second transport system for sn-glycerol-3-phosphate in Escherichia coli. J. Bacteriol. 1978, 3, 1070–1083. [Google Scholar]
- Lamarche, M.G.; Wanner, B.L.; Crepin, S.; Harel, J. The phosphate regulon and bacterial virulence: A regulatory network connecting phosphate homeostasis and pathogenesis. FEMS Microbiol. Rev. 2008, 3, 461–473. [Google Scholar] [CrossRef]
- Halsey, T.A.; Vazquez-Torres, A.; Gravdahl, D.J.; Fang, F.C.; Libby, S.J. The ferritin-like Dps protein is required for Salmonella enterica serovar Typhimurium oxidative stress resistance and virulence. Infect. Immun. 2004, 2, 1155–1158. [Google Scholar] [CrossRef]
- Wanner, B.L. Gene regulation by phosphate in enteric bacteria. J. Cell. Biochem. 1993, 1, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, M.G.; Dozois, C.M.; Daigle, F.; Caza, M.; Curtiss, R.R.; Dubreuil, J.D.; Harel, J. Inactivation of the pst system reduces the virulence of an avian pathogenic Escherichia coli O78 strain. Infect. Immun. 2005, 7, 4138–4145. [Google Scholar] [CrossRef] [PubMed]
- Srinivasa, R.P.; Lim, T.M.; Leung, K.Y. Functional genomics approach to the identification of virulence genes involved in Edwardsiella tarda pathogenesis. Infect. Immun. 2003, 3, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, C.P.; Cameron, A.D.S.; Dorman, C.J. H-NS silences gfp, the green fluorescent protein gene: GfpTCD is a genetically remastered gfp gene with reduced susceptibility to H-NS-Mediated transcription silencing and with enhanced translation. J. Bacteriol. 2010, 18, 4790–4793. [Google Scholar] [CrossRef] [PubMed]
- Atlung, T.; Ingmer, H. H-NS: A modulator of environmentally regulated gene expression. Mol. Microbiol. 1997, 1, 7–17. [Google Scholar] [CrossRef]
- Alpuche, A.C.; Swanson, J.A.; Loomis, W.P.; Miller, S.I. Salmonella typhimurium activates virulence gene transcription within acidified macrophage phagosomes. Proc. Natl. Acad. Sci. USA 1992, 21, 10079–10083. [Google Scholar] [CrossRef] [PubMed]
- Karem, K.; Foster, J.W. The influence of DNA topology on the environmental regulation of a pH-regulated locus in Salmonella typhimurium. Mol. Microbiol. 1993, 1, 75–86. [Google Scholar] [CrossRef]
- Rao, N.N.; Torriani, A. Molecular aspects of phosphate transport in Escherichia coli. Mol. Microbiol. 1990, 7, 1083–1090. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Q.; Hu, M.; Yu, K.; Fu, J.; Zhou, F.; Liu, X. Proteomic analyses of intracellular salmonella enterica serovar typhimurium reveal extensive bacterial adaptations to infected host epithelial cells. Infect. Immun. 2015, 7, 2897–2906. [Google Scholar] [CrossRef]
- Stapels, D.; Hill, P.; Westermann, A.J.; Fisher, R.A.; Thurston, T.L.; Saliba, A.E.; Blommestein, I.; Vogel, J.; Helaine, S. Salmonella persisters undermine host immune defenses during antibiotic treatment. Science 2018, 6419, 1156–1160. [Google Scholar] [CrossRef]
- Murphy, F.V., 4th; Ramakrishnan, V.; Malkiewicz, A.; Agris, P.F. The role of modifications in codon discrimination by tRNA(Lys)UUU. Nat. Struct. Mol. Biol. 2004, 12, 1186. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Yoshida, T.; Tanaka, K.; Sasakawa, C.; Mizuno, T. Molecular analysis of the Escherichia coli hns gene encoding a DNA-binding protein, which preferentially recognizes curved DNA sequences. Mol. Gen. Genet. 1991, 230, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Higashi, K.; Tobe, T.; Kanai, A.; Uyar, E.; Ishikawa, S.; Suzuki, Y.; Ogasawara, N.; Kurokawa, K.; Oshima, T. H-NS facilitates sequence diversification of horizontally transferred DNAs during their integration in host chromosomes. Plos Genet. 2016, 1, e1005796. [Google Scholar] [CrossRef] [PubMed]
- Navarre, W.W.; Porwollik, S.; Wang, Y.; McClelland, M.; Rosen, H.; Libby, S.J.; Fang, F.C. Selective silencing of foreign DNA with low GC content by the H-NS protein in Salmonella. Science 2006, 5784, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Goldberg, M.D.; Olsson, T.; Esposito, D.; Hinton, J.C.; Ladbury, J.E. H-NS is a part of a thermally controlled mechanism for bacterial gene regulation. Biochem. J. 2005, 391, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucchini, S.; Rowley, G.; Goldberg, M.D.; Hurd, D.; Harrison, M.; Hinton, J.C. H-NS mediates the silencing of laterally acquired genes in bacteria. Plos Pathog. 2006, 8, e81. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.; Fowler, R.; Kinsella, N.; Howell, G.; Farris, M.; Coote, P.; O’Connor, C.D. Proteomic detection of PhoPQ- and acid-mediated repression of Salmonella motility. Proteomics 2001, 4, 597–607. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 12, 6640–6645. [Google Scholar] [CrossRef]
- Wang, R.F.; Kushner, S.R. Construction of versatile low-copy-number vectors for cloning, sequencing and gene expression in Escherichia coli. Gene 1991, 100, 195–199. [Google Scholar] [CrossRef]
- Coombes, B.K.; Brown, N.F.; Valdez, Y.; Brumell, J.H.; Finlay, B.B. Expression and secretion of Salmonella pathogenicity island-2 virulence genes in response to acidification exhibit differential requirements of a functional type III secretion apparatus and SsaL. J. Biol. Chem. 2004, 48, 49804–49815. [Google Scholar] [CrossRef]
- Bustamante, V.H.; Martinez, L.C.; Santana, F.J.; Knodler, L.A.; Steele-Mortimer, O.; Puente, J.L. HilD-mediated transcriptional cross-talk between SPI-1 and SPI-2. Proc. Natl. Acad. Sci. USA 2008, 38, 14591–14596. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.C.; Yakhnin, H.; Camacho, M.I.; Georgellis, D.; Babitzke, P.; Puente, J.L.; Bustamante, V.H. Integration of a complex regulatory cascade involving the SirA/BarA and Csr global regulatory systems that controls expression of the Salmonella SPI-1 and SPI-2 virulence regulons through HilD. Mol. Microbiol. 2011, 6, 1637–1656. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Feng, L.; Yang, B.; Zhang, W.; Wang, P.; Jiang, X.; Wang, L. Signal transduction pathway mediated by the novel regulator LoiA for low oxygen tension induced Salmonella Typhimurium invasion. Plos Pathog. 2017, 6, e1006429. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 4, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 9, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 4, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Valdivia, R.H.; Falkow, S. Fluorescence-based isolation of bacterial genes expressed within host cells. Science 1997, 5334, 2007–2011. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype or Description | Source | |
|---|---|---|
| S. Typhimurium strains | ||
| WT | Wild-type S. Typhimurium strain 14028s | Our lab |
| ΔyaeB | WT strain yaeB::Cm; CmR | This study |
| ΔpstSCAB | WT strain pstSCAB::Km; KmR | This study |
| Δhns | WT strain hns::Cm; CmR | This study |
| ΔyaeBΔpstSCAB | WT strain yaeB::Cm, pstSCAB::Km; CmR, KmR | This study |
| ΔhnsΔyaeB | WT strain hns::Cm; yaeB::Km; CmR, KmR | This study |
| cYaeB | ΔyaeB harboring plasmid pYaeB; CmR, KmR; complement strains | This study |
| BL21 | For expressing protein | TransGen Biotech |
| Plasmids | ||
| pKD4 | For λRed recombination; KmR | Our lab |
| pKD3 | For λRed recombination; CmR | Our lab |
| pWSK129 | For generating complementation strains; KmR | Our lab |
| pKD46 | For generating mutant strains with λ Red recombinase system under an arabinose-inducible promoter; ApR | Our lab |
| pET28a | Expression vector; KmR | Our lab |
| pYaeB | pWSK129 carrying the WT yaeB gene; KmR | This study |
| pET-H-NS | pET28a carrying the WT hns gene; KmR | This study |
| Target Gene | Primer Sequence (5′–3′) | ||
|---|---|---|---|
| Primers for construction of mutants* | |||
| yaeB | F | ATGAGCAGCTTTCAGTTTGAACAAATCGGCGTTATTCGGTGTAGGCTGGAGCTGCTTCG | |
| R | TTACCGTGGTTCCAGCGCAAACACTTCAAATCCCGAGTCATATGAATATCCTCCTTAG | ||
| pstSCAB | F | ATGAAAGTTATGCGTACCACTGTCGCAACTGTTGTCGCGTGTAGGCTGGAGCTGCTTCG | |
| R | TTAACCGTAACGACCGGTGATATAGTCTTCTGTTTGTTCATATGAATATCCTCCTTAG | ||
| hns | F | ATGAGCGAAGCACTTAAAATTCTGAACAACATCCGTACGTGTAGGCTGGAGCTGCTTCG | |
| R | TTATTCCTTGATCAGGAAATCTTCCAGTTGCTTACCTTCATATGAATATCCTCCTTAG | ||
| Primers for identification of mutants | |||
| yaeB | F | GCCAGGCTGCTATCGTCA | |
| R | TGTTGTTACGGTTCCAGTT | ||
| pstSCAB | F | CCTGGGCACGATTGTCTG | |
| R | GCGGTGATAGCGTCAGAAA | ||
| hns | F | CTCAACAAACCACCCCAATA | |
| R | GGCTTGAAGAAGAAATGGGTA | ||
| rpoS | F | ATAAGGAGTAACCGATGATTTG | |
| R | CTACGCCCATAATGATACG | ||
| phoP | F | TTTGCCCGTTTCATCGTA | |
| R | TGCCGAAGGAATGAAACA | ||
| Primers for construction of clones and complemented strains | |||
| yaeB | F1 | CGGGATCCACGCCAGCGAAAGACGGTAT | |
| R1 | AATATTTCCTCAATGAATGT | ||
| F2 | CAATATCTGGCAATTAGAACATTCATTGAGGAAATATTATGAGCAGCTTTCAGTTTGAA | ||
| R2 | CCCAAGCTTTTACCGTGGTTCCAGCGCAAAC | ||
| hns-his6 | F | GGAATTCCATATGATGAGCGAAGCACTTAAAAT | |
| R | CCGCTCGAGTTATTCCTTGATCAGGAAAT | ||
| qRT-PCR primers | |||
| 16S rRNA | F | GAAAGCGTGGgGAGCAAAC | |
| R | ACATGCTCCACCGCTTGTG | ||
| yaeB | F | GCCTGGAAGCGTTCAGCCATT | |
| R | CGAGCGACATGCCAATCGGATT | ||
| pstA | F | GCTACGCTTGATATGCAGAACA | |
| R | GCCATCGTCGCCATTGAGA | ||
| pstB | F | ACGCCGTTCCCGATGTCCAT | |
| R | ACAGACGCTGCTGCTGACCA | ||
| pstC | F | GCTGATTGTGCTATTGATGT | |
| R | GCGTCCCATTCCTTAGTC | ||
| pstS | F | GCCAAATGGGCGGATACCTACC | |
| R | TCACCGTCGGGAACTGGAACA | ||
| hns | F | GCTGCTGCTGCTGAAGTGGAA | |
| R | GCTGCGCGTTTAGCTTTGGTAC | ||
| Primers for EMSAs | |||
| Phns | F | CGACAGACGGTGAGTATC | |
| R | TGCGTTCTTCCACTTCAG | ||
| PyaeB | F | CAAAGACGCCGAGAATGTG | |
| R | GCCAGATATTGAATCAGGAGC | ||
| STM0272 | F | CACCACGCTATTACTCACCACC | |
| R | GCCGTGCTGTTCACTCATCC | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Song, X.; Wang, P.; Lv, R.; Ma, S.; Jiang, L. YaeB, Expressed in Response to the Acidic pH in Macrophages, Promotes Intracellular Replication and Virulence of Salmonella Typhimurium. Int. J. Mol. Sci. 2019, 20, 4339. https://doi.org/10.3390/ijms20184339
Zhang H, Song X, Wang P, Lv R, Ma S, Jiang L. YaeB, Expressed in Response to the Acidic pH in Macrophages, Promotes Intracellular Replication and Virulence of Salmonella Typhimurium. International Journal of Molecular Sciences. 2019; 20(18):4339. https://doi.org/10.3390/ijms20184339
Chicago/Turabian StyleZhang, Huan, Xiaorui Song, Peisheng Wang, Runxia Lv, Shuangshuang Ma, and Lingyan Jiang. 2019. "YaeB, Expressed in Response to the Acidic pH in Macrophages, Promotes Intracellular Replication and Virulence of Salmonella Typhimurium" International Journal of Molecular Sciences 20, no. 18: 4339. https://doi.org/10.3390/ijms20184339
APA StyleZhang, H., Song, X., Wang, P., Lv, R., Ma, S., & Jiang, L. (2019). YaeB, Expressed in Response to the Acidic pH in Macrophages, Promotes Intracellular Replication and Virulence of Salmonella Typhimurium. International Journal of Molecular Sciences, 20(18), 4339. https://doi.org/10.3390/ijms20184339