Efficacy of Azatyrosine-Phenylbutyric Hydroxamides, a Histone Deacetylase Inhibitor, on Chemotherapy-Induced Gastrointestinal Mucositis


Abstract
:1. Introduction
2. Results
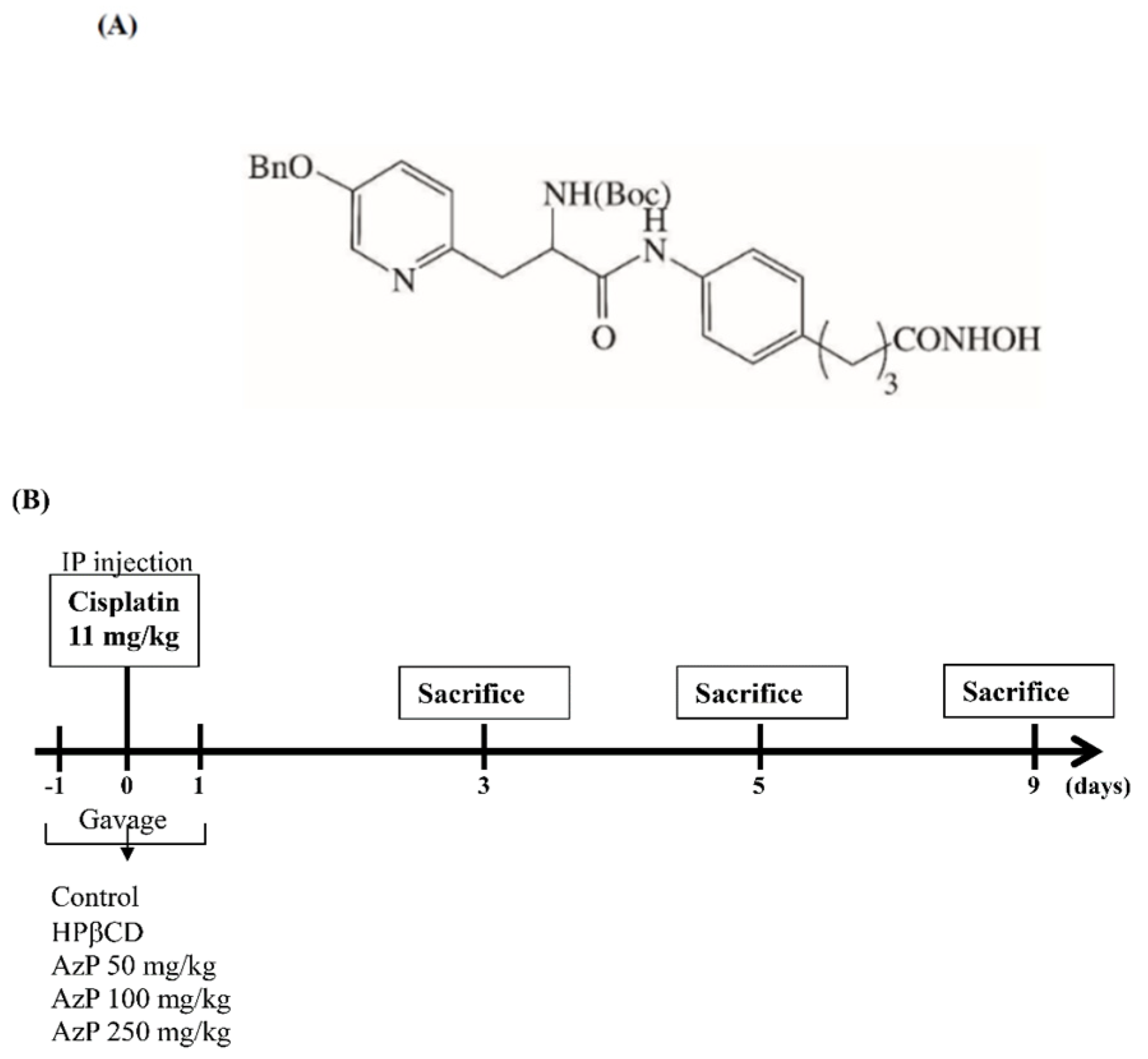
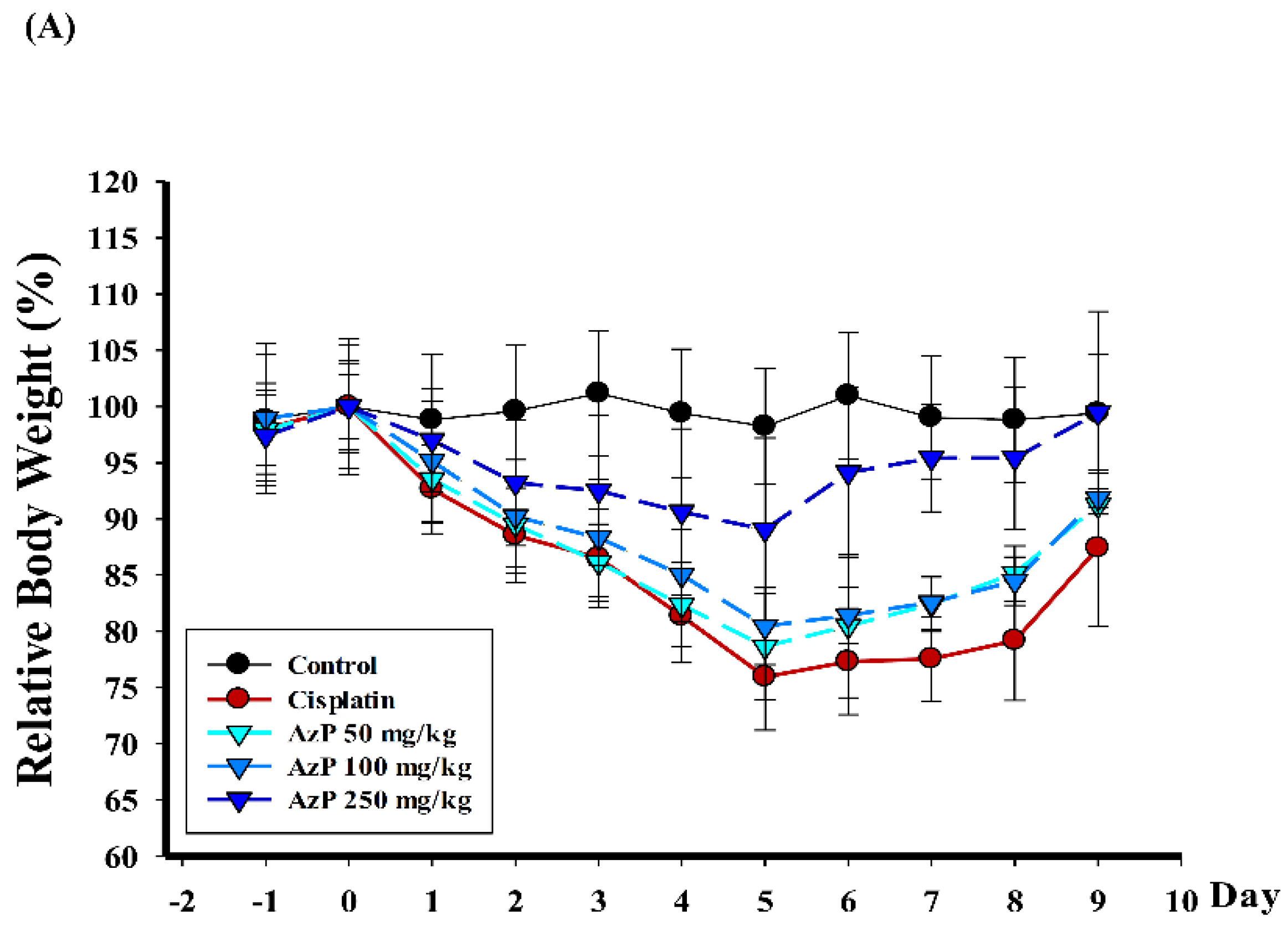
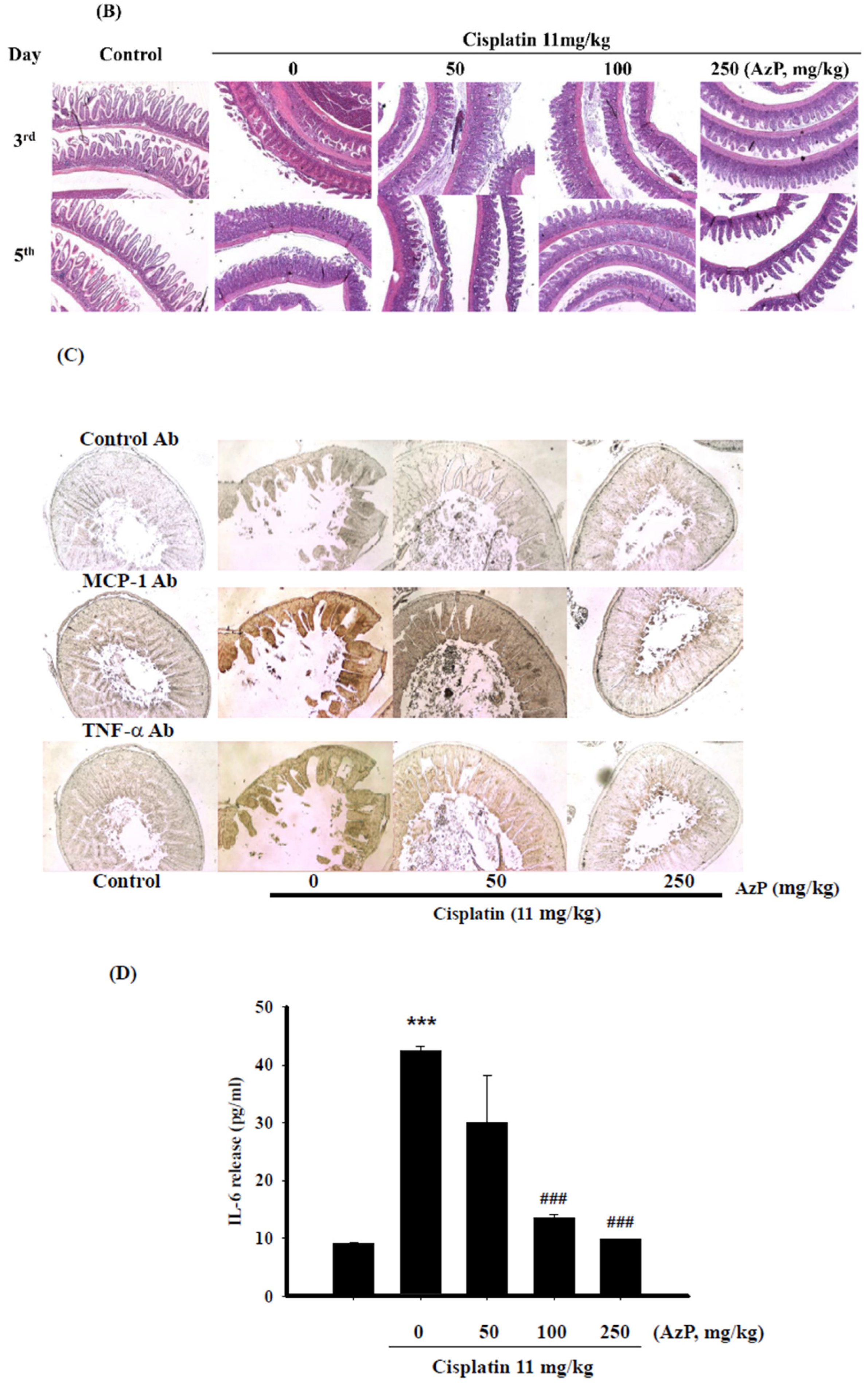
2.1. Protective Effect of Azatyrosine-Phenylbutyric Hydroxamides (Aza-PBHA) in Mucositis Model
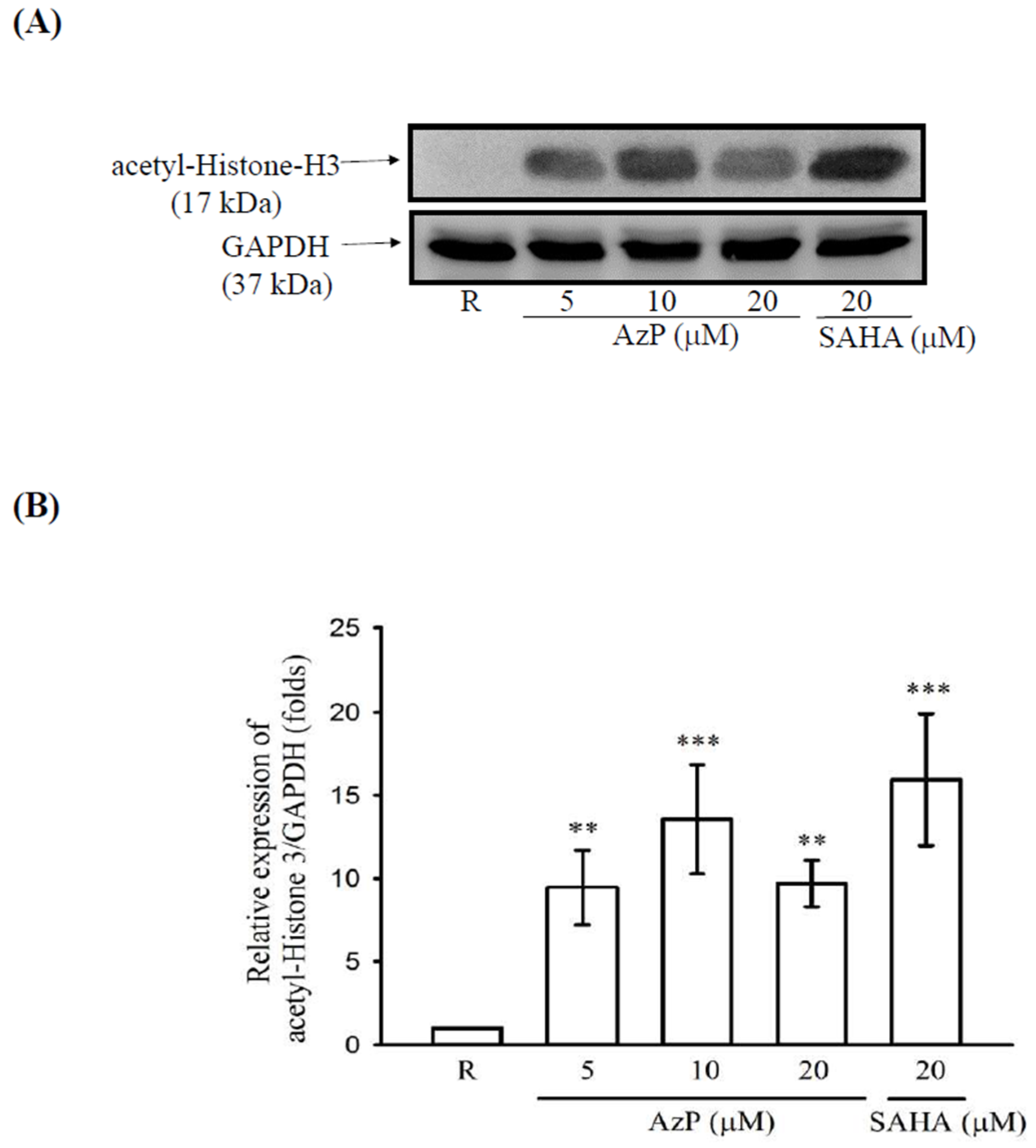
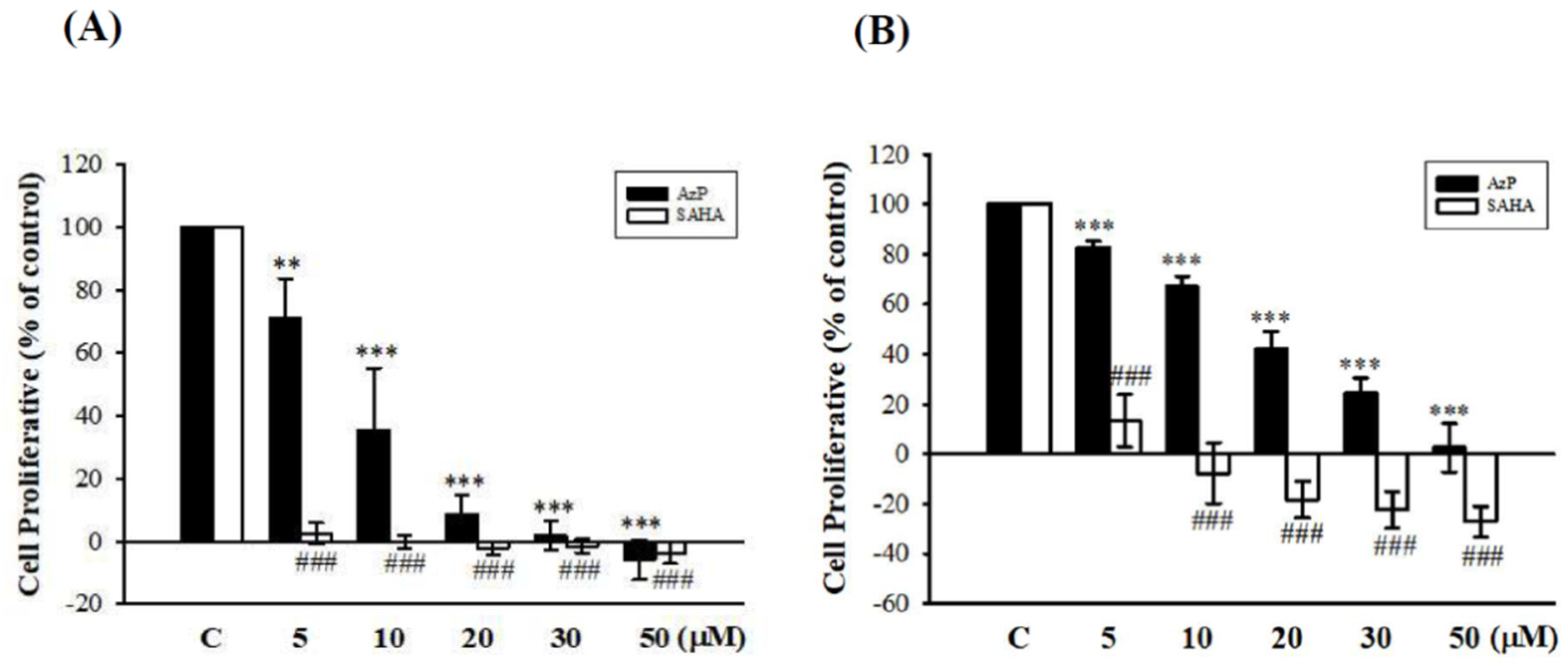
2.2. Aza-PBHA Showed Histone Deacetylase (HDAC) Inhibition Activity
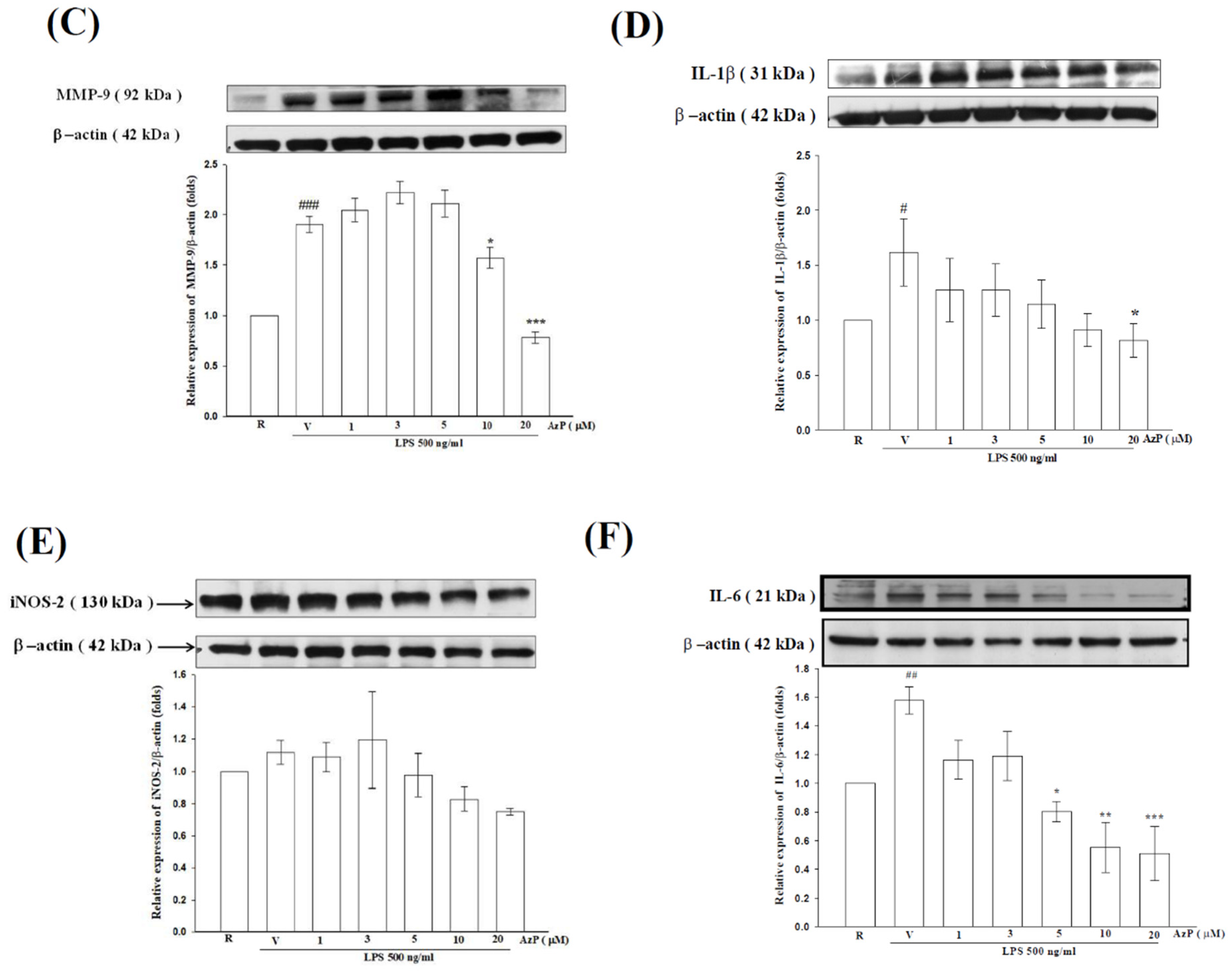
2.3. AzP Exerted In Vitro Anti-Inflammatory Potential
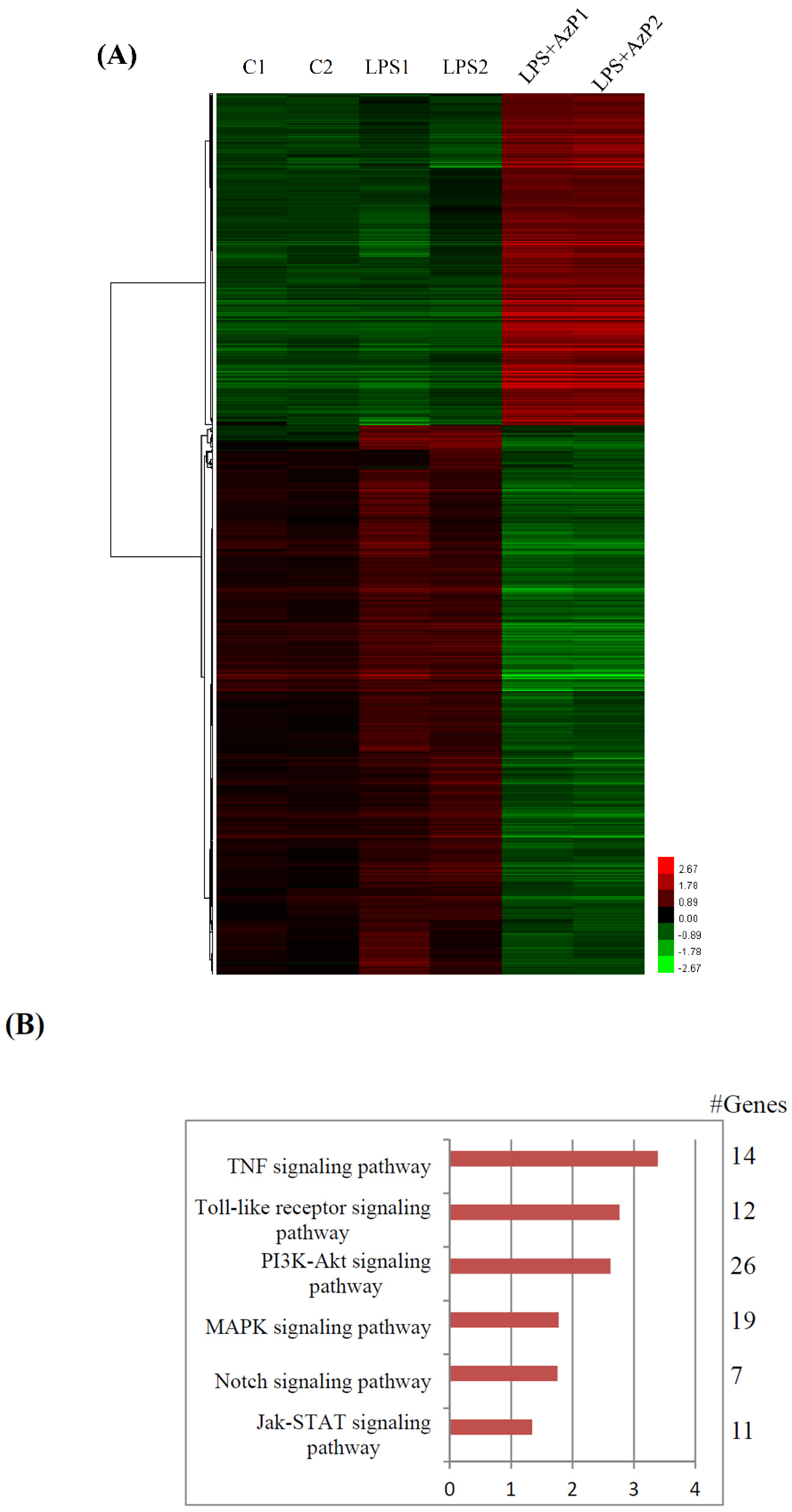
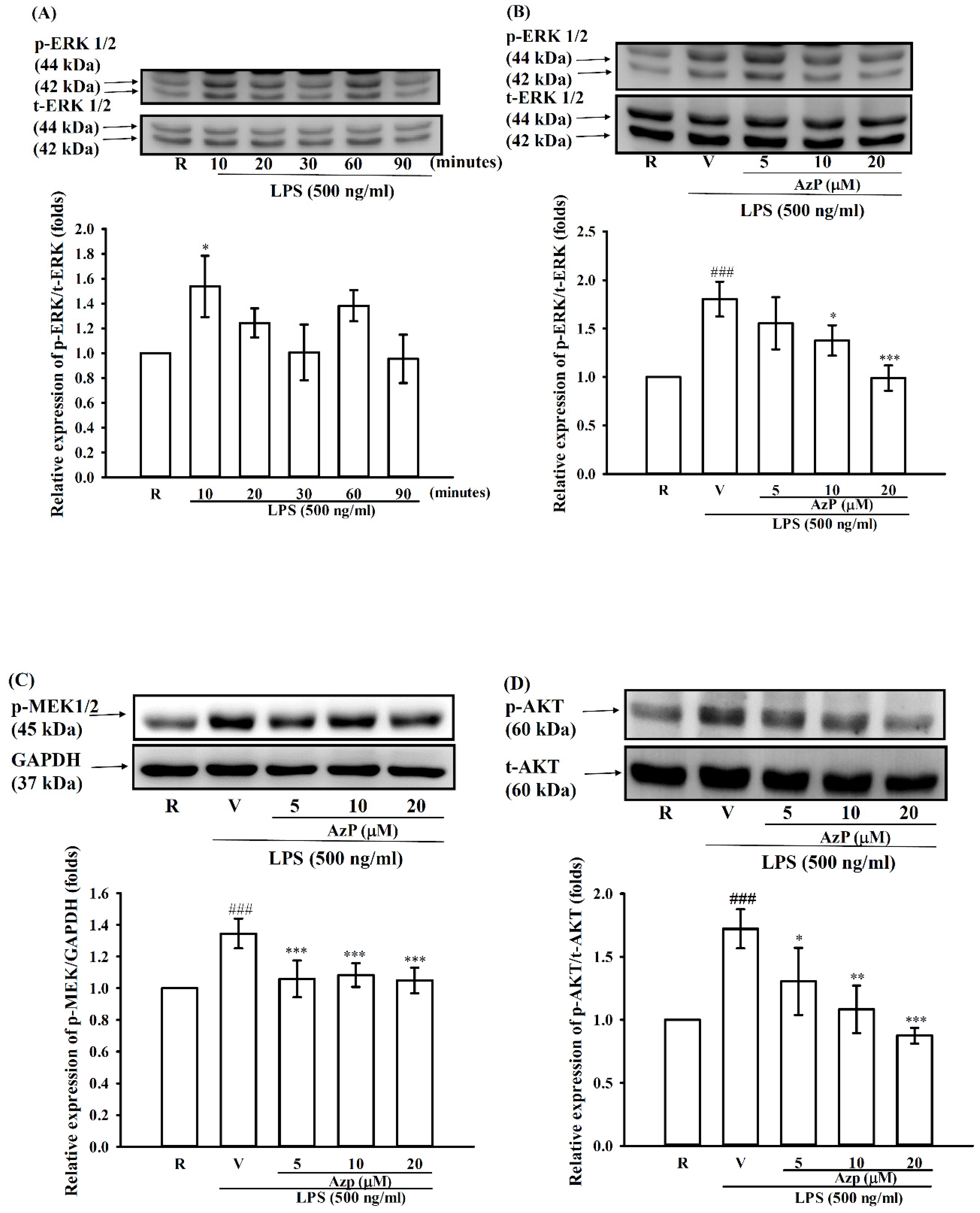
2.4. Possible Molecular Mechanisms of AzP in IEC-6 Cells
2.5. AzP Did Not Cause Mutagenicity or Clastogenicity In Vitro or In Vivo
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. HDAC Enzyme Activity Assay
4.4. Animal Housing
4.5. Mucositis Model
4.6. Histological and Immunohistological Observations of Crypt and Villus
4.7. Western Blot Analysis
4.8. Oligonucleotide DNA Microarray
4.9. Salmonella/Microsome Reversion Assay: Ames Test
4.10. Sulforhodamine B (SRB) Assay
4.11. Chromosomal Aberrations
4.12. Micronucleus Test
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Reyes-Gibby, C.C.; Melkonian, S.C.; Wang, J.; Yu, R.K.; Shelburne, S.A.; Lu, C.; Gunn, G.B.; Chambers, M.S.; Hanna, E.Y.; Yeung, S.J.; et al. Identifying novel genes and biological processes relevant to the development of cancer therapy-induced mucositis: An informative gene network analysis. PLoS ONE 2017, 12, e0180396. [Google Scholar] [CrossRef] [PubMed]
- Stansborough, R.L.; Al-Dasooqi, N.; Bateman, E.H.; Keefe, D.M.; Gibson, R.J. Radiotherapy-induced gut toxicity: Involvement of matrix metalloproteinases and the intestinal microvasculature. Int. J. Radiat. Biol. 2016, 92, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.B., 3rd; Ajani, J.A.; Catalano, R.B.; Engelking, C.; Kornblau, S.M.; Martenson, J.A., Jr.; McCallum, R.; Mitchell, E.P.; O’Dorisio, T.M.; Vokes, E.E.; et al. Recommended guidelines for the treatment of cancer treatment-induced diarrhea. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2004, 22, 2918–2926. [Google Scholar] [CrossRef] [PubMed]
- Maria, O.M.; Eliopoulos, N.; Muanza, T. Radiation-Induced Oral Mucositis. Front. Oncol. 2017, 7, 89. [Google Scholar] [CrossRef] [PubMed]
- Sonis, S.T. The pathobiology of mucositis. Nat. Rev. Cancer 2004, 4, 277–284. [Google Scholar] [CrossRef]
- Shahid, F.; Farooqui, Z.; Khan, F. Cisplatin-induced gastrointestinal toxicity: An update on possible mechanisms and on available gastroprotective strategies. Eur. J. Pharmacol. 2018, 827, 49–57. [Google Scholar] [CrossRef]
- Haberland, M.; Montgomery, R.L.; Olson, E.N. The many roles of histone deacetylases in development and physiology: Implications for disease and therapy. Nat. Rev. Genet. 2009, 10, 32–42. [Google Scholar] [CrossRef]
- Felice, C.; Lewis, A.; Armuzzi, A.; Lindsay, J.O.; Silver, A. Review article: Selective histone deacetylase isoforms as potential therapeutic targets in inflammatory bowel diseases. Aliment. Pharmacol. Ther. 2015, 41, 26–38. [Google Scholar] [CrossRef]
- De Las Heras, J.; Aldamiz-Echevarria, L.; Martinez-Chantar, M.L.; Delgado, T.C. An update on the use of benzoate, phenylacetate and phenylbutyrate ammonia scavengers for interrogating and modifying liver nitrogen metabolism and its implications in urea cycle disorders and liver disease. Expert Opin. Drug Metab. Toxicol. 2017, 13, 439–448. [Google Scholar] [CrossRef]
- Kusaczuk, M.; Bartoszewicz, M.; Cechowska-Pasko, M. Phenylbutyric Acid: Simple structure—Multiple effects. Curr. Pharm. Des. 2015, 21, 2147–2166. [Google Scholar] [CrossRef]
- Kusaczuk, M.; Kretowski, R.; Bartoszewicz, M.; Cechowska-Pasko, M. Phenylbutyrate-a pan-HDAC inhibitor-suppresses proliferation of glioblastoma LN-229 cell line. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.L.; Lee, M.Y.; Pui, N.N. Epigenetic therapy using the histone deacetylase inhibitor for increasing therapeutic gain in oral cancer: Prevention of radiation-induced oral mucositis and inhibition of chemical-induced oral carcinogenesis. Carcinogenesis 2009, 30, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Yen, S.H.; Wang, L.W.; Lin, Y.H.; Jen, Y.M.; Chung, Y.L. Phenylbutyrate mouthwash mitigates oral mucositis during radiotherapy or chemoradiotherapy in patients with head-and-neck cancer. Int. J. Radiat. Oncol. Biol. Phys. 2012, 82, 1463–1470. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.C.; Cohen, S.; Stewart, M.; Rivas, R.; Lison, P. Radioprotection by the histone deacetylase inhibitor phenylbutyrate. Radiat. Environ. Biophys. 2011, 50, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.C.; Rivas, R.; McMahon, R.; Miller, K.; Tesoro, L.; Villa, V.; Yanushkevich, D.; Lison, P. Radiation protection and mitigation potential of phenylbutyrate: Delivered via oral administration. Int. J. Radiat. Biol. 2017, 93, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y. Mechanism-based management for mucositis: Option for treating side effects without compromising the efficacy of cancer therapy. OncoTargets Ther. 2016, 9, 2007–2016. [Google Scholar] [CrossRef]
- Tsirigotis, P.; Triantafyllou, K.; Girkas, K.; Giannopoulou, V.; Ioannidou, E.; Chondropoulos, S.; Kalli, T.; Papaxoinis, G.; Pappa, V.; Papageorgiou, E.; et al. Keratinocyte growth factor is effective in the prevention of intestinal mucositis in patients with hematological malignancies treated with high-dose chemotherapy and autologous hematopoietic SCT: A video-capsule endoscopy study. Bone Marrow Transplant. 2008, 42, 337–343. [Google Scholar] [CrossRef]
- Blijlevens, N.; Sonis, S. Palifermin (recombinant keratinocyte growth factor-1): A pleiotropic growth factor with multiple biological activities in preventing chemotherapy- and radiotherapy-induced mucositis. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2007, 18, 817–826. [Google Scholar] [CrossRef]
- Bian, L.; Han, G.; Zhao, C.W.; Garl, P.J.; Wang, X.J. The role of Smad7 in oral mucositis. Protein Cell 2015, 6, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Abidi, M.H.; Agarwal, R.; Tageja, N.; Ayash, L.; Deol, A.; Al-Kadhimi, Z.; Abrams, J.; Cronin, S.; Ventimiglia, M.; Lum, L.; et al. A phase I dose-escalation trial of high-dose melphalan with palifermin for cytoprotection followed by autologous stem cell transplantation for patients with multiple myeloma with normal renal function. Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 2013, 19, 56–61. [Google Scholar] [CrossRef]
- Lee, C.S.; Ryan, E.J.; Doherty, G.A. Gastro-intestinal toxicity of chemotherapeutics in colorectal cancer: The role of inflammation. World J. Gastroenterol. 2014, 20, 3751–3761. [Google Scholar] [CrossRef] [PubMed]
- Ong, Z.Y.; Gibson, R.J.; Bowen, J.M.; Stringer, A.M.; Darby, J.M.; Logan, R.M.; Yeoh, A.S.; Keefe, D.M. Pro-inflammatory cytokines play a key role in the development of radiotherapy-induced gastrointestinal mucositis. Radiat. Oncol. 2010, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, H.; Ishihara, K.; Takeda, Y.; Koizumi, W.; Ichikawa, T. Changes in the mucus barrier during cisplatin-induced intestinal mucositis in rats. BioMed Res. Int. 2013, 2013, 276186. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, Y.; Gou, W.F.; Zhao, S.; Takano, Y.; Zheng, H.C. The anti-tumor effects and molecular mechanisms of suberoylanilide hydroxamic acid (SAHA) on the aggressive phenotypes of ovarian carcinoma cells. PLoS ONE 2013, 8, e79781. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- Chen, Z.; Gibson, T.B.; Robinson, F.; Silvestro, L.; Pearson, G.; Xu, B.; Wright, A.; Vanderbilt, C.; Cobb, M.H. MAP kinases. Chem. Rev. 2001, 101, 2449–2476. [Google Scholar] [CrossRef]
- Xu, Z.H.; Shun, W.W.; Hang, J.B.; Gao, B.L.; Hu, J.A. Posttranslational modifications of FOXO1 regulate epidermal growth factor receptor tyrosine kinase inhibitor resistance for non-small cell lung cancer cells. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 5485–5495. [Google Scholar] [CrossRef]
- Bialkowska, A.B.; Ghaleb, A.M.; Nandan, M.O.; Yang, V.W. Improved Swiss-rolling Technique for Intestinal Tissue Preparation for Immunohistochemical and Immunofluorescent Analyses. J. Vis. Exp. 2016, 113, e54161. [Google Scholar] [CrossRef]
- Li, C.H.; Liao, J.W.; Liao, P.L.; Huang, W.K.; Tse, L.S.; Lin, C.H.; Kang, J.J.; Cheng, Y.W. Evaluation of Acute 13-Week Subchronic Toxicity and Genotoxicity of the Powdered Root of Tongkat Ali (Eurycoma longifolia Jack). Evid. Based Complement. Altern. Med. 2013, 2013, 102987. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Activity (IC50, nM) | AzP | SAHA | Ration: AzP/SAHA |
|---|---|---|---|
| Pan-HDAC | 1650 ± 103 | 84.5 ± 13.7 | 20:1 |
| HDAC1 | 3440 ± 680 | 566.5 ± 65.6 | 6:1 |
| HDAC3 | 660 ± 99 | 72.5 ± 5.4 | 9.2:1 |
| HDAC4 | 1970 ± 110 | 106.2 ± 27.3 | 106:1 |
| Test Compound | Gene Mutation in Bacteria-Salmonella/Microsome Reversion Assay (Ames Test) of AzP | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AzP | Revertant colonies per plate (-S9) | Revertant colonies per plate (|S9) | ||||||||
| TA98 | TA100 | TA102 | TA1535 | TA1537 | TA98 | TA100 | TA102 | TA1535 | TA1537 | |
| Negative | Negative | |||||||||
| Chromosome aberration assay of AzP in Chinese Hamster Ovary cells | ||||||||||
| 3 h without S9 metabolic activation | 3 h with S9 metabolic activation | 24 h without S9 metabolic activation | ||||||||
| Negative | Negative | Negative | ||||||||
| Micronucleus Test of AzP on male ICR mice | ||||||||||
| AzP treatment after 24 h | AzP treatment after 48 h | AzP treatment after 72 h | ||||||||
| Negative | Negative | Negative | ||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, P.-L.; Huang, S.-H.; Hung, C.-H.; Huang, W.-K.; Tsai, C.-H.; Kang, J.-J.; Wang, H.-P.; Cheng, Y.-W. Efficacy of Azatyrosine-Phenylbutyric Hydroxamides, a Histone Deacetylase Inhibitor, on Chemotherapy-Induced Gastrointestinal Mucositis. Int. J. Mol. Sci. 2019, 20, 249. https://doi.org/10.3390/ijms20020249
Liao P-L, Huang S-H, Hung C-H, Huang W-K, Tsai C-H, Kang J-J, Wang H-P, Cheng Y-W. Efficacy of Azatyrosine-Phenylbutyric Hydroxamides, a Histone Deacetylase Inhibitor, on Chemotherapy-Induced Gastrointestinal Mucositis. International Journal of Molecular Sciences. 2019; 20(2):249. https://doi.org/10.3390/ijms20020249
Chicago/Turabian StyleLiao, Po-Lin, Shih-Hsuan Huang, Chien-Hung Hung, Wei-Kuang Huang, Chi-Hao Tsai, Jaw-Jou Kang, Hui-Po Wang, and Yu-Wen Cheng. 2019. "Efficacy of Azatyrosine-Phenylbutyric Hydroxamides, a Histone Deacetylase Inhibitor, on Chemotherapy-Induced Gastrointestinal Mucositis" International Journal of Molecular Sciences 20, no. 2: 249. https://doi.org/10.3390/ijms20020249
APA StyleLiao, P. -L., Huang, S. -H., Hung, C. -H., Huang, W. -K., Tsai, C. -H., Kang, J. -J., Wang, H. -P., & Cheng, Y. -W. (2019). Efficacy of Azatyrosine-Phenylbutyric Hydroxamides, a Histone Deacetylase Inhibitor, on Chemotherapy-Induced Gastrointestinal Mucositis. International Journal of Molecular Sciences, 20(2), 249. https://doi.org/10.3390/ijms20020249