Activation of Sirtuin 3 and Maintenance of Mitochondrial Integrity by N-Acetylcysteine Protects Against Bisphenol A-Induced Kidney and Liver Toxicity in Rats


Abstract
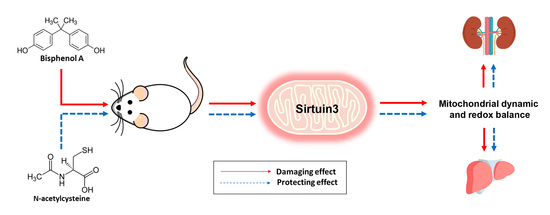
:
1. Introduction
2. Results
2.1. Effects of BPA Exposure and NAC Treatment on Body Weight, Kidney Weight, Liver Weight, Food and Water Intake
2.2. Effects of BPA Exposure and NAC Treatment on Kidney and Liver Functions
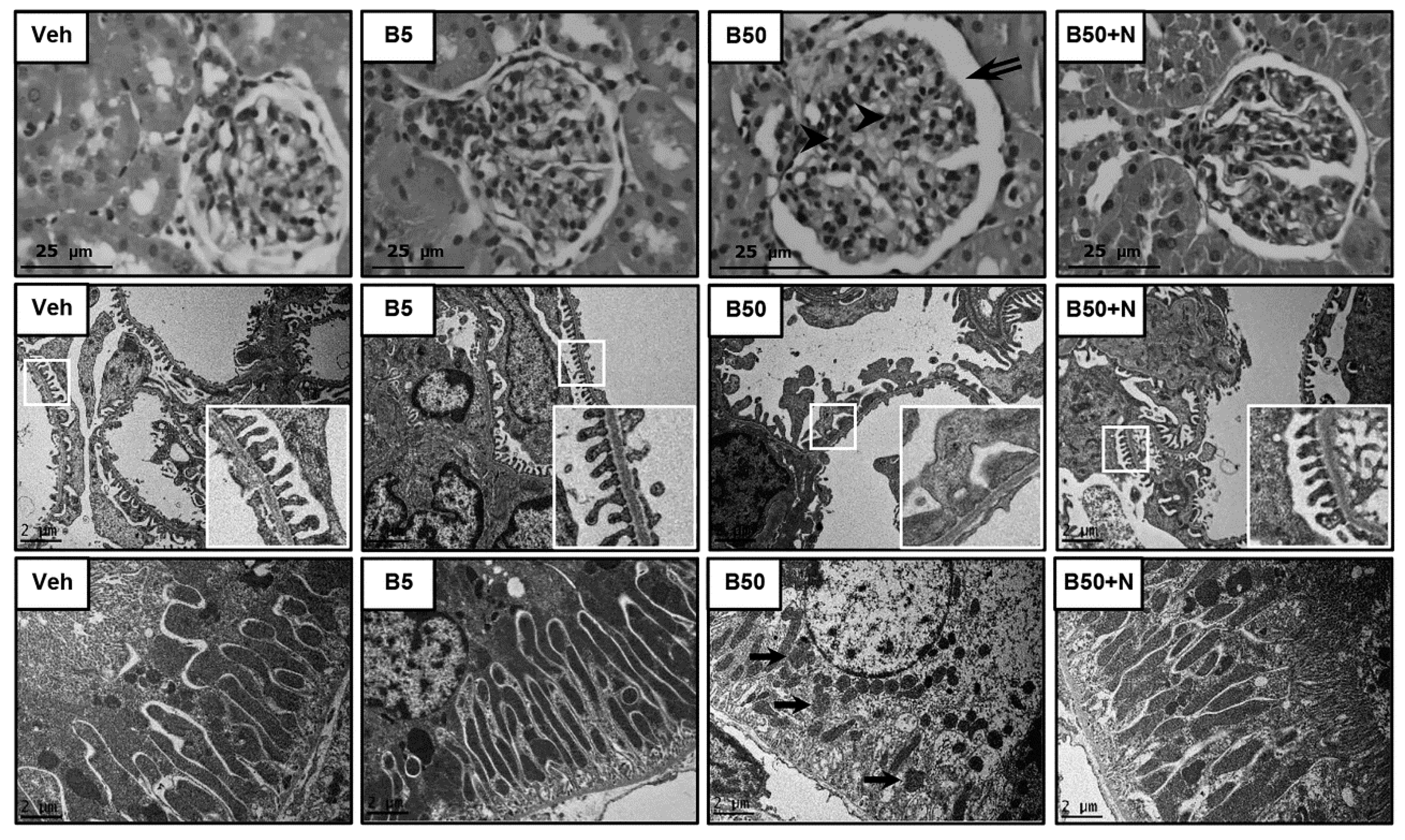
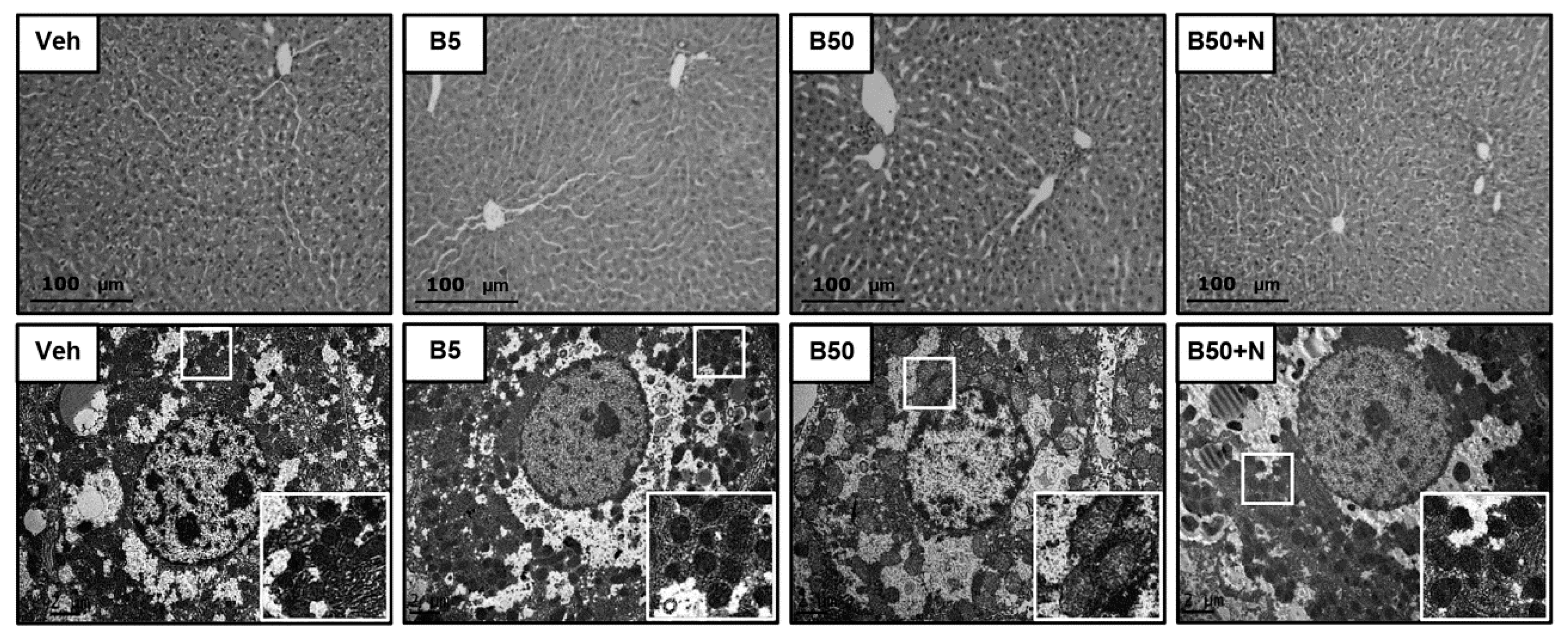
2.3. Effects of BPA Exposure and NAC Treatment on Histopathology of Kidney and Liver
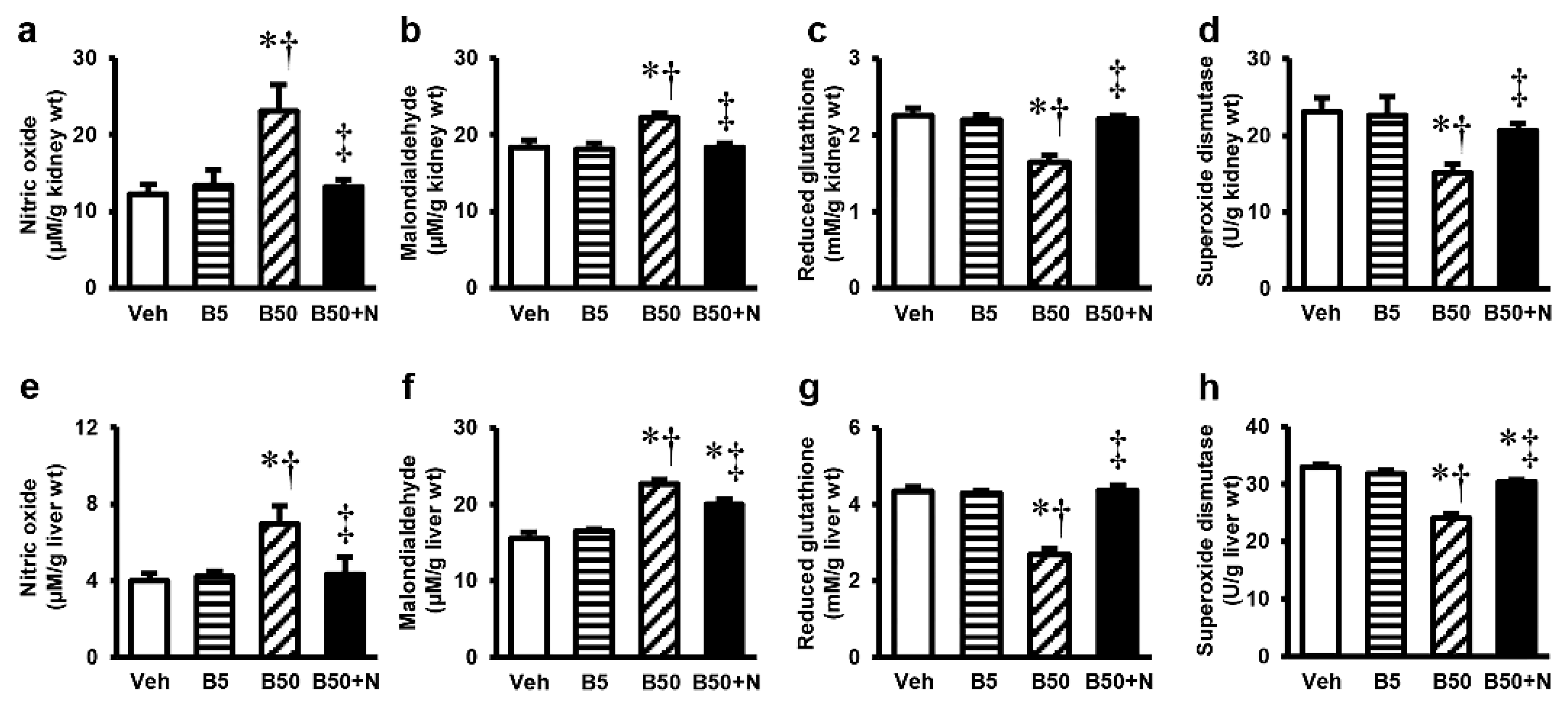
2.4. Effects of BPA Exposure and NAC Treatment on Kidney and Liver Oxidative Stress
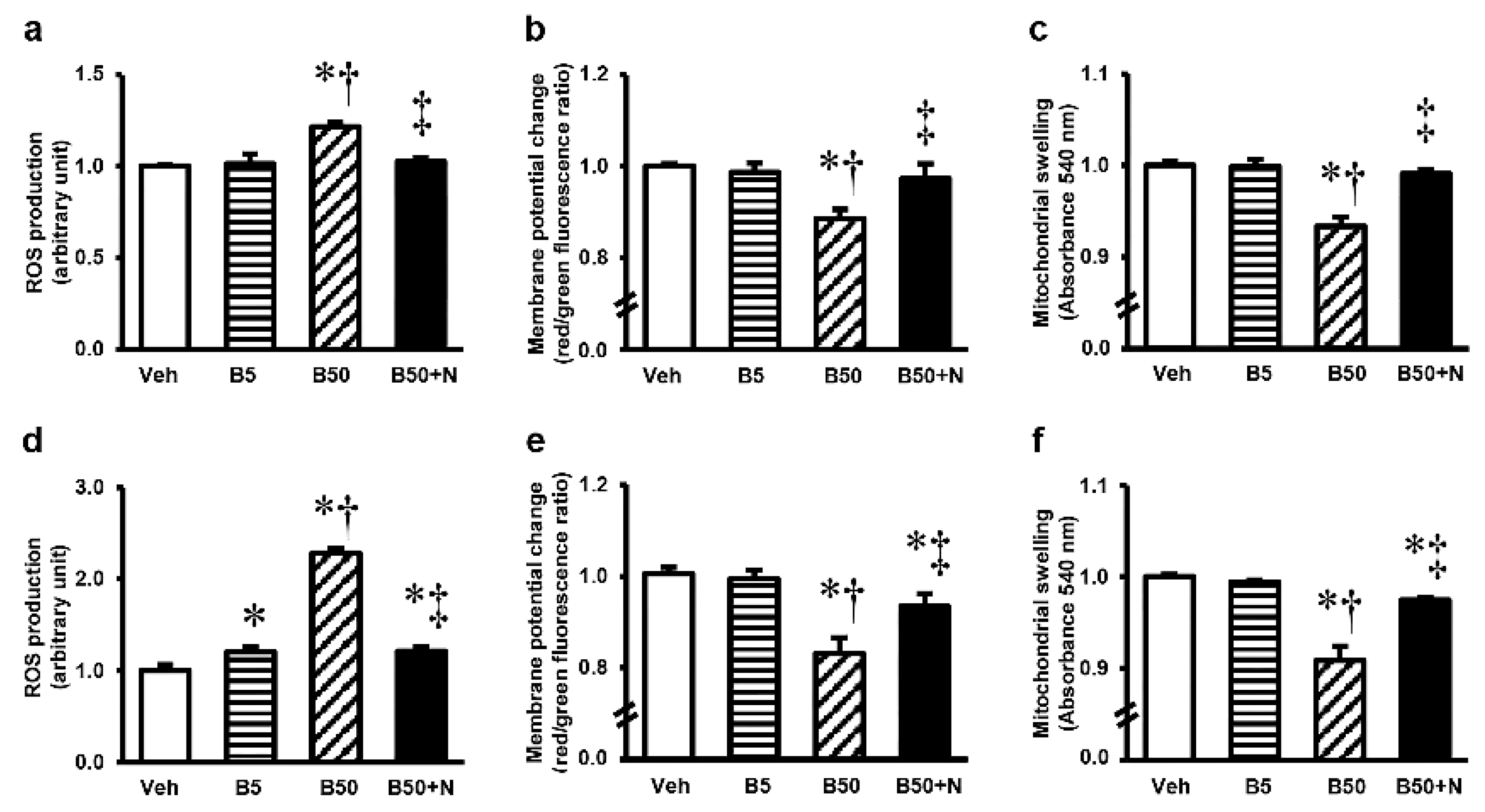
2.5. Effects of BPA Exposure and NAC Treatment on Kidney and Liver Mitochondrial Functions
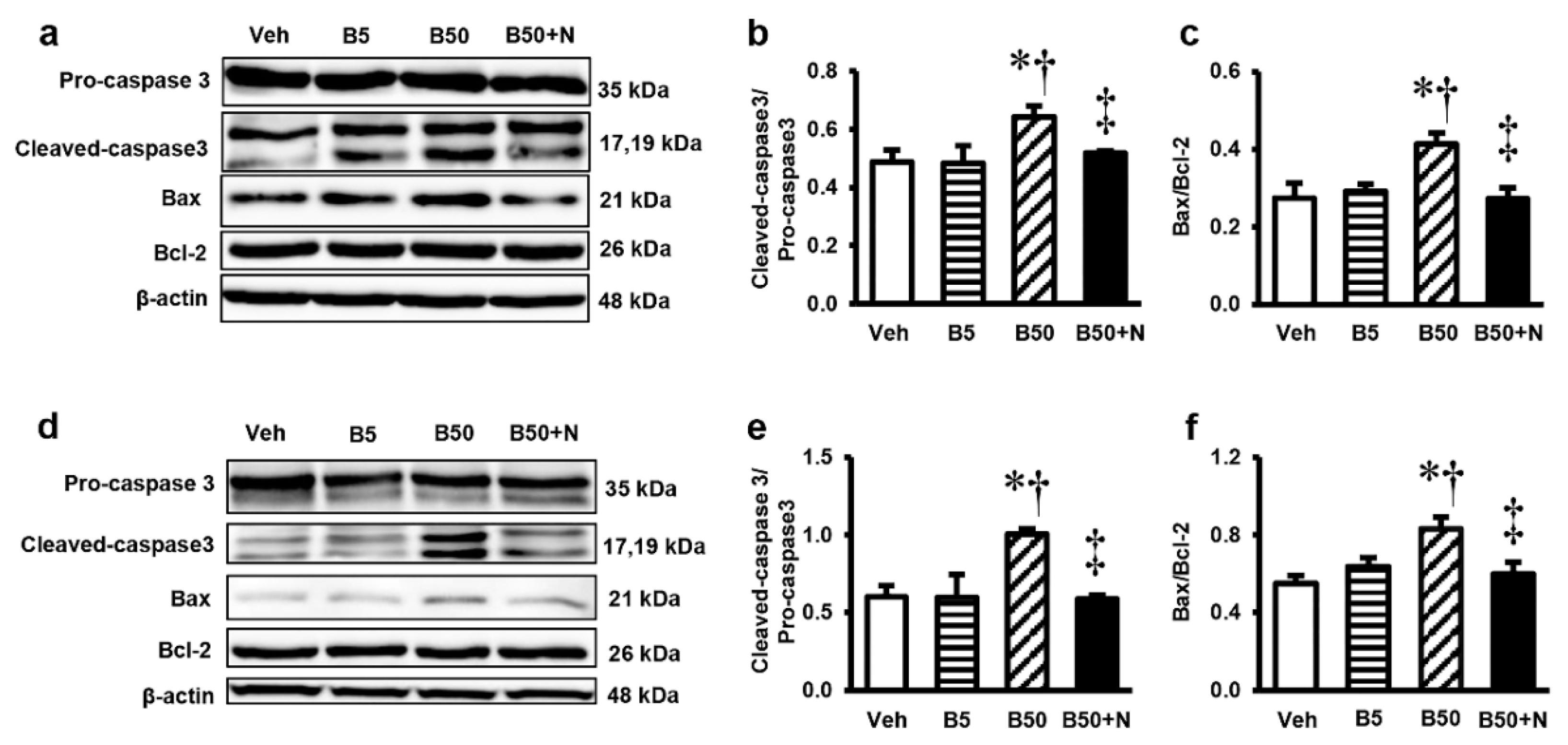
2.6. Effects of BPA Exposure and NAC Treatment on the Levels of Pro-Caspase3, Cleaved-Caspase3, Pro-Apoptotic Bax and Anti-Apoptotic Bcl-2 in the Kidney and Liver
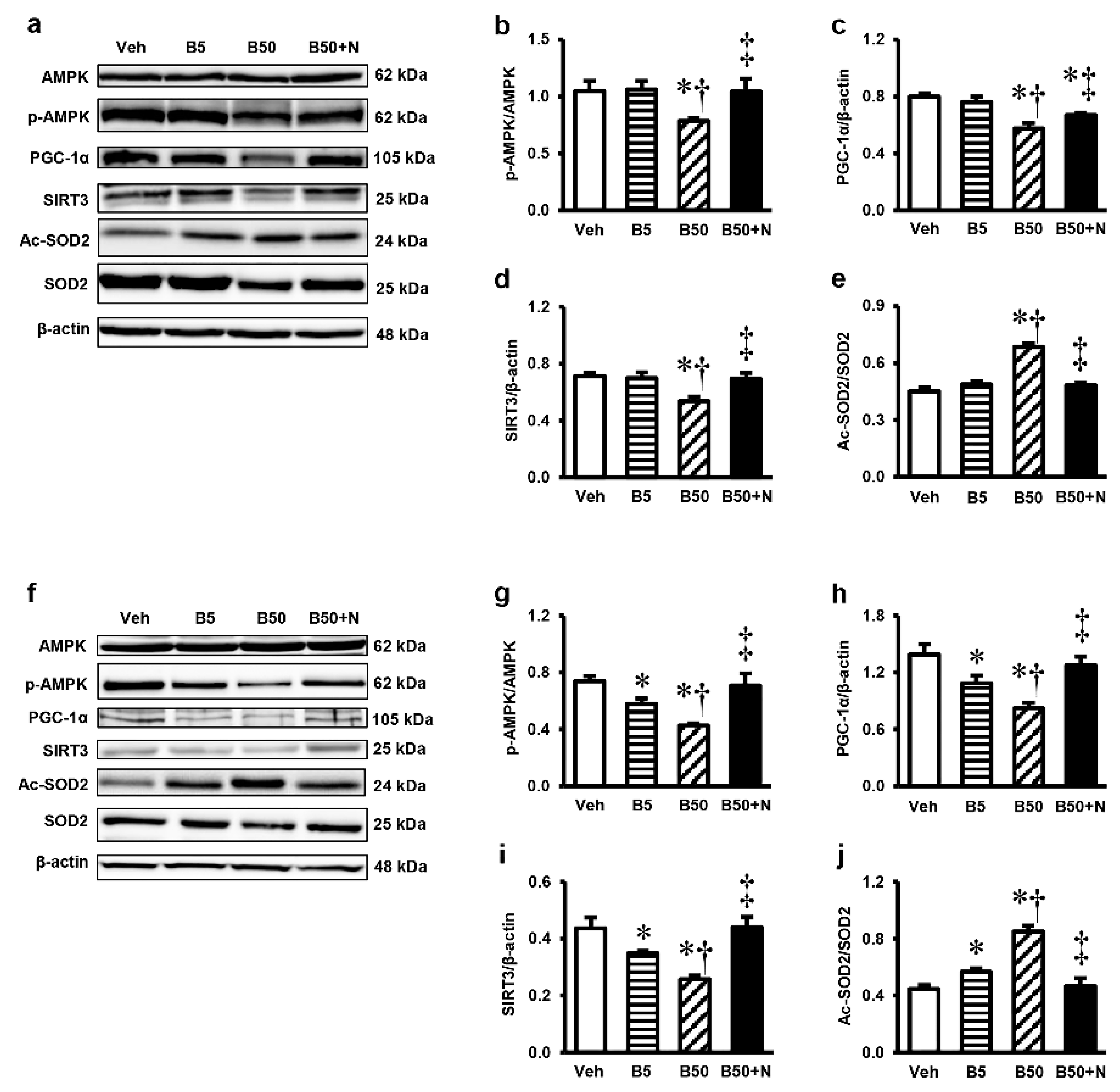
2.7. Effects of BPA Exposure and NAC Treatment on p-AMPK, AMPK, PGC-1α, SIRT3, Ac-SOD2 and SOD2 Expressions in the Kidney and Liver
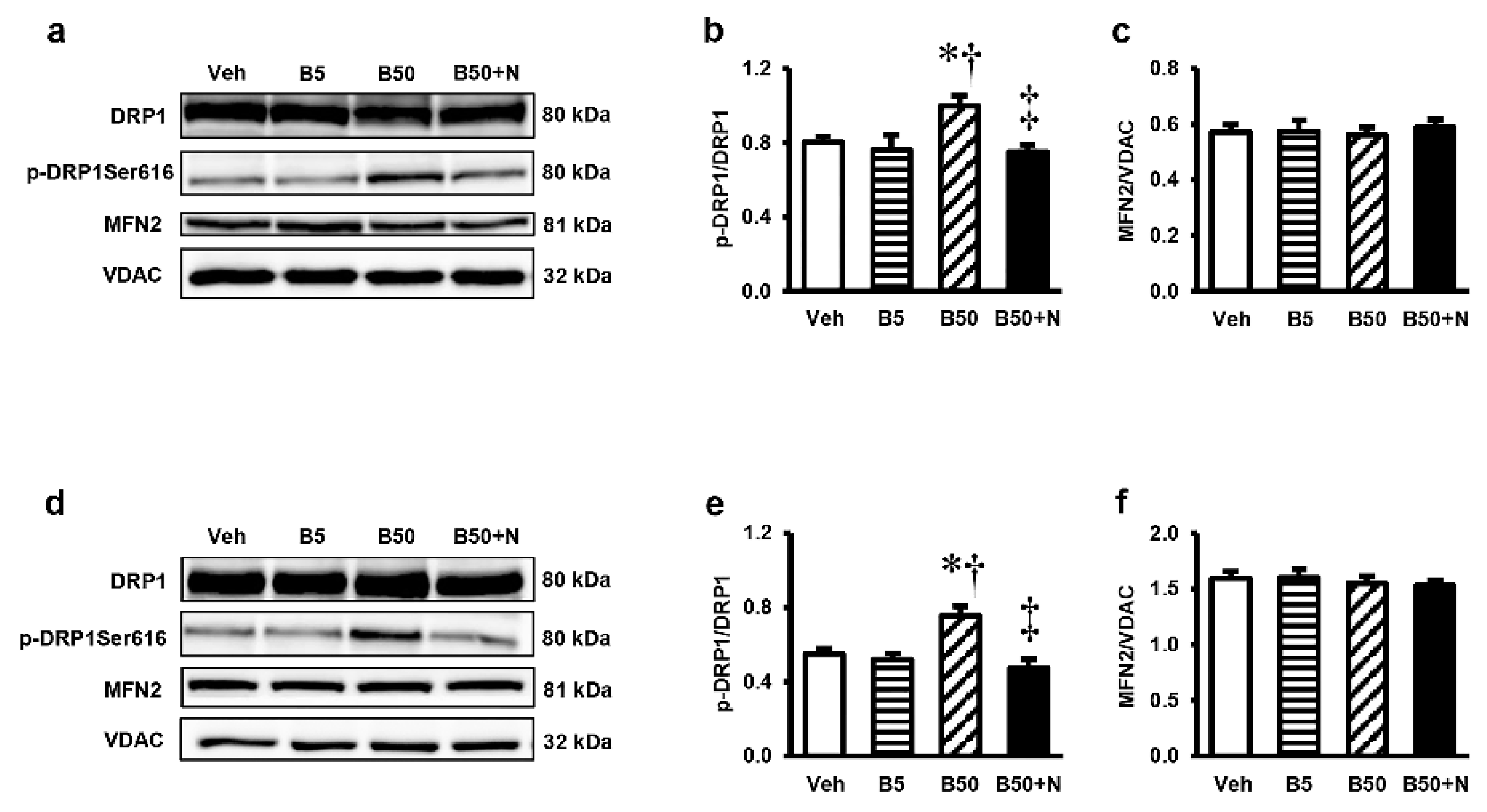
2.8. Effects of BPA Exposure and NAC Treatment on p-DRP, DRP1, and MFN2 in the Kidney and Liver
3. Discussion
4. Materials and Methods
4.1. Drugs and Chemicals
4.2. Animals
4.3. Experimental Designs
4.4. Determinations of Renal and Liver Functions
4.5. Determinations of Renal and Liver Oxidative Stress
4.6. Histopathological Studies
4.7. Electron Microscopic Studies
4.8. Preparation of Mitochondrial Fractions and Mitochondrial Proteins
4.9. Determination of Mitochondrial ROS Production
4.10. Determination of Mitochondrial Membrane Potential (ΔΨm)
4.11. Determination of Mitochondrial Swelling
4.12. Western Blot Analysis
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Ac-SOD2 | Acetylated superoxide dismutase 2 |
| AMPK | AMP-activated protein kinase |
| BPA | Bisphenol A |
| DCFDA | Dichlorofluorescin diacetate |
| DRP1 | Dynamin-related protein 1 |
| GSH | Reduced glutathione |
| JC-1 | 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimi-dazocarbocyanine iodide |
| MDA | Malondialdehyde |
| MFN2 | Mitofusin 2 |
| MnSOD | Manganese superoxide dismutase |
| NAC | N-acetylcysteine |
| NO | Nitric oxide |
| NOAEL | No observed adverse effect level |
| p-AMPK | Phosphorylated AMP-activated protein kinase |
| p-DRP1 | Phosphorylated dynamin-related protein 1 |
| PGC-1α | Peroxisome proliferator-activated gamma receptor coactivator-1α |
| SIRT3 | Silent information regulator T3 or sirtuin 3 |
| SOD2 | Superoxide dismutase 2 |
| TBST | Tris-buffered saline-Tween 20 |
| UPCR | Urine protein-to-creatinine ratio |
| VDAC | Voltage-dependent anion channel |
References
- Kataria, A.; Trasande, L.; Trachtman, H. The effects of environmental chemicals on renal function. Nat. Rev. Nephrol. 2015, 11, 610–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trasande, L.; Attina, T.M.; Trachtman, H. Bisphenol A exposure is associated with low-grade urinary albumin excretion in children of the United States. Kidney Int. 2013, 83, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Hines, C.J.; Jackson, M.V.; Deddens, J.A.; Clark, J.C.; Ye, X.; Christianson, A.L.; Meadows, J.W.; Calafat, A.M. Urinary Bisphenol A (BPA) Concentrations among Workers in Industries that Manufacture and Use BPA in the USA. Ann. Work Expo. Health 2017, 61, 164–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Beigh, S.; Chaudhari, B.P.; Sharma, S.; Aliul Hasan Abdi, S.; Ahmad, S.; Ahmad, F.; Parvez, S.; Raisuddin, S. Mitochondrial dysfunction induced by Bisphenol A is a factor of its hepatotoxicity in rats. Environ. Toxicol. 2016, 31, 1922–1934. [Google Scholar] [CrossRef] [PubMed]
- Kobroob, A.; Peerapanyasut, W.; Chattipakorn, N.; Wongmekiat, O. Damaging Effects of Bisphenol A on the Kidney and the Protection by Melatonin: Emerging Evidences from In Vivo and In Vitro Studies. Oxid. Med. Cell. Longev. 2018, 2018, 3082438. [Google Scholar] [CrossRef] [PubMed]
- Rezg, R.; El-Fazaa, S.; Gharbi, N.; Mornagui, B. Bisphenol A and human chronic diseases: Current evidences, possible mechanisms, and future perspectives. Environ. Int. 2014, 64, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial energetics in the kidney. Nat. Rev. Nephrol. 2017, 13, 629–646. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, K.Q.; Moura, F.A.; dos Santos, J.M.; de Araujo, O.R.; de Farias Santos, J.C.; Goulart, M.O. Oxidative Stress and Inflammation in Hepatic Diseases: Therapeutic Possibilities of N-Acetylcysteine. Int. J. Mol. Sci. 2015, 16, 30269–30308. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Kaur, T.; Singla, S.K. Protective effects of N-acetylcysteine against hyperoxaluria induced mitochondrial dysfunction in male wistar rats. Mol. Cell Biochem. 2015, 405, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Ferrin, G.; Hidalgo, A.B.; Ranchal, I.; Lopez-Cillero, P.; Santos-Gonzalez, M.; Lopez-Lluch, G.; Briceno, J.; Gomez, M.A.; Poyato, A.; et al. N-acetylcysteine, coenzyme Q10 and superoxide dismutase mimetic prevent mitochondrial cell dysfunction and cell death induced by d-galactosamine in primary culture of human hepatocytes. Chem. Biol. Interact. 2009, 181, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Wongjaikam, S.; Kumfu, S.; Khamseekaew, J.; Sripetchwandee, J.; Srichairatanakool, S.; Fucharoen, S.; Chattipakorn, S.C.; Chattipakorn, N. Combined Iron Chelator and Antioxidant Exerted Greater Efficacy on Cardioprotection Than Monotherapy in Iron-Overloaded Rats. PLoS ONE 2016, 11, e0159414. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Kumar, C.H.; Suranagi, U.D.; Mediratta, P.K. Protective effect of N-acetylcysteine on bisphenol A-induced cognitive dysfunction and oxidative stress in rats. Food Chem. Toxicol. 2011, 49, 1404–1409. [Google Scholar] [CrossRef] [PubMed]
- Morigi, M.; Perico, L.; Benigni, A. Sirtuins in Renal Health and Disease. J. Am. Soc. Nephrol. 2018, 29, 1799–1809. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.; Ladeira, C.; Viegas, S. Occupational Exposure to Bisphenol A (BPA): A Reality That Still Needs to Be Unveiled. Toxics 2017, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Tyl, R.W.; Myers, C.B.; Marr, M.C.; Thomas, B.F.; Keimowitz, A.R.; Brine, D.R.; Veselica, M.M.; Fail, P.A.; Chang, T.Y.; Seely, J.C.; et al. Three-generation reproductive toxicity study of dietary bisphenol A in CD Sprague-Dawley rats. Toxicol. Sci. 2002, 68, 121–146. [Google Scholar] [CrossRef] [PubMed]
- Vahdati Hassani, F.; Mehri, S.; Abnous, K.; Birner-Gruenberger, R.; Hosseinzadeh, H. Protective effect of crocin on BPA-induced liver toxicity in rats through inhibition of oxidative stress and downregulation of MAPK and MAPKAP signaling pathway and miRNA-122 expression. Food Chem. Toxicol. 2017, 107 Pt A, 395–405. [Google Scholar] [CrossRef]
- Pottenger, L.H.; Domoradzki, J.Y.; Markham, D.A.; Hansen, S.C.; Cagen, S.Z.; Waechter, J.M., Jr. The relative bioavailability and metabolism of bisphenol A in rats is dependent upon the route of administration. Toxicol. Sci. 2000, 54, 3–18. [Google Scholar] [CrossRef]
- Saura, M.; Marquez, S.; Reventun, P.; Olea-Herrero, N.; Arenas, M.I.; Moreno-Gomez-Toledano, R.; Gomez-Parrizas, M.; Munoz-Moreno, C.; Gonzalez-Santander, M.; Zaragoza, C.; et al. Oral administration of bisphenol A induces high blood pressure through angiotensin II/CaMKII-dependent uncoupling of eNOS. FASEB J. 2014, 28, 4719–4728. [Google Scholar] [CrossRef] [Green Version]
- Olea-Herrero, N.; Arenas, M.I.; Munoz-Moreno, C.; Moreno-Gomez-Toledano, R.; Gonzalez-Santander, M.; Arribas, I.; Bosch, R.J. Bisphenol-A induces podocytopathy with proteinuria in mice. J. Cell. Physiol. 2014, 229, 2057–2066. [Google Scholar] [CrossRef]
- Hassan, Z.K.; Elobeid, M.A.; Virk, P.; Omer, S.A.; ElAmin, M.; Daghestani, M.H.; AlOlayan, E.M. Bisphenol A induces hepatotoxicity through oxidative stress in rat model. Oxid. Med. Cell. Longev. 2012, 2012, 194829. [Google Scholar] [CrossRef]
- Perico, L.; Morigi, M.; Benigni, A. Mitochondrial Sirtuin 3 and Renal Diseases. Nephron 2016, 134, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Benigni, A.; Perico, L.; Macconi, D. Mitochondrial Dynamics Is Linked to Longevity and Protects from End-Organ Injury: The Emerging Role of Sirtuin 3. Antioxid. Redox Signal 2016, 25, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Gong, B.; Duan, W.; Fan, C.; Zhang, J.; Li, Z.; Xue, X.; Xu, Y.; Meng, D.; Li, B.; et al. Melatonin ameliorates myocardial ischemia/reperfusion injury in type 1 diabetic rats by preserving mitochondrial function: Role of AMPK-PGC-1alpha-SIRT3 signaling. Sci. Rep. 2017, 7, 41337. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.Y.; Zhang, L.; Sui, M.X.; Zhu, Y.H.; Zeng, L. Protective effects of sirtuin 3 in a murine model of sepsis-induced acute kidney injury. Sci. Rep. 2016, 6, 33201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morigi, M.; Perico, L.; Rota, C.; Longaretti, L.; Conti, S.; Rottoli, D.; Novelli, R.; Remuzzi, G.; Benigni, A. Sirtuin 3-dependent mitochondrial dynamic improvements protect against acute kidney injury. J. Clin. Investig. 2015, 125, 715–726. [Google Scholar] [CrossRef]
- Peerapanyasut, W.; Thamprasert, K.; Wongmekiat, O. Ubiquinol supplementation protects against renal ischemia and reperfusion injury in rats. Free Radic. Res. 2014, 48, 180–189. [Google Scholar] [CrossRef]
- Kobroob, A.; Chattipakorn, N.; Wongmekiat, O. Caffeic acid phenethyl ester ameliorates cadmium-induced kidney mitochondrial injury. Chem. Biol. Interact. 2012, 200, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, I.; Parvez, S.; Winkler-Stuck, K.; Seitz, G.; Trieu, I.; Wallesch, C.W.; Schonfeld, P.; Siemen, D. Patch clamp reveals powerful blockade of the mitochondrial permeability transition pore by the D2-receptor agonist pramipexole. FASEB J. 2006, 20, 556–558. [Google Scholar] [CrossRef]
- Walker, J.E. The regulation of catalysis in ATP synthase. Curr. Opin. Struct. Biol. 1994, 4, 912–918. [Google Scholar] [CrossRef]
- Beavis, A.D.; Brannan, R.D.; Garlid, K.D. Swelling and contraction of the mitochondrial matrix. I. A structural interpretation of the relationship between light scattering and matrix volume. J. Biol. Chem. 1985, 260, 13424–13433. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Veh | B5 | B50 | B50 + N |
|---|---|---|---|---|
| Initial BW (g) | 240.00 ± 2.89 | 238.33 ± 2.79 | 239.17 ± 2.39 | 241.67 ± 1.05 |
| BW gain (%) | 60.80 ± 1.40 | 61.70 ± 1.56 | 59.62 ± 2.43 | 59.47 ± 1.41 |
| KW/BW (*100) | 0.59 ± 0.01 | 0.58 ± 0.01 | 0.59 ± 0.02 | 0.58 ± 0.01 |
| LW/BW (*100) | 4.12 ± 0.13 | 3.97 ± 0.08 | 4.15 ± 0.21 | 3.98 ± 0.15 |
| Food intake (g/day) | 26.56 ± 0.65 | 27.44 ± 0.58 | 27.22 ± 0.78 | 26.72 ± 0.50 |
| Water intake (mL/day) | 39.44 ± 1.27 | 38.89 ± 0.56 | 38.06 ± 1.32 | 38.61 ± 1.69 |
| Parameters | Veh | B5 | B50 | B50 + N |
|---|---|---|---|---|
| BUN (mg/dL) | 21.85 ± 0.72 | 21.40 ± 0.50 | 21.35 ± 0.47 | 22.15 ± 1.08 |
| SCr (mg/dL) | 0.29 ± 0.01 | 0.30 ± 0.01 | 0.30 ± 0.01 | 0.28 ± 0.01 |
| CCr (ml/min/g kidney wt) | 1.29 ± 0.08 | 1.28 ± 0.11 | 1.58 ± 0.04 *† | 1.23 ± 0.10 ‡ |
| UPCR | 0.86 ± 0.08 | 0.83 ± 0.04 | 2.49 ± 0.41 *† | 0.98 ± 0.09 ‡ |
| UProtV (mg/24 h/g kidney wt) | 3.78 ± 0.38 | 3.35 ± 0.35 | 8.00 ± 0.74 *† | 3.55 ± 0.62 ‡ |
| AST (U/L) | 80.33 ± 4.13 | 81.50 ± 4.15 | 114.83 ± 3.9 *† | 91.50 ± 1.86 *‡ |
| ALT (U/L) | 24.60 ± 1.33 | 25.00 ± 1.64 | 33.20 ± 0.80 *† | 29.60 ± 0.24 *‡ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peerapanyasut, W.; Kobroob, A.; Palee, S.; Chattipakorn, N.; Wongmekiat, O. Activation of Sirtuin 3 and Maintenance of Mitochondrial Integrity by N-Acetylcysteine Protects Against Bisphenol A-Induced Kidney and Liver Toxicity in Rats. Int. J. Mol. Sci. 2019, 20, 267. https://doi.org/10.3390/ijms20020267
Peerapanyasut W, Kobroob A, Palee S, Chattipakorn N, Wongmekiat O. Activation of Sirtuin 3 and Maintenance of Mitochondrial Integrity by N-Acetylcysteine Protects Against Bisphenol A-Induced Kidney and Liver Toxicity in Rats. International Journal of Molecular Sciences. 2019; 20(2):267. https://doi.org/10.3390/ijms20020267
Chicago/Turabian StylePeerapanyasut, Wachirasek, Anongporn Kobroob, Siripong Palee, Nipon Chattipakorn, and Orawan Wongmekiat. 2019. "Activation of Sirtuin 3 and Maintenance of Mitochondrial Integrity by N-Acetylcysteine Protects Against Bisphenol A-Induced Kidney and Liver Toxicity in Rats" International Journal of Molecular Sciences 20, no. 2: 267. https://doi.org/10.3390/ijms20020267
APA StylePeerapanyasut, W., Kobroob, A., Palee, S., Chattipakorn, N., & Wongmekiat, O. (2019). Activation of Sirtuin 3 and Maintenance of Mitochondrial Integrity by N-Acetylcysteine Protects Against Bisphenol A-Induced Kidney and Liver Toxicity in Rats. International Journal of Molecular Sciences, 20(2), 267. https://doi.org/10.3390/ijms20020267