Potential Interplay of the Gatipotuzumab Epitope TA-MUC1 and Estrogen Receptors in Ovarian Cancer

 ,
,  and
and
Abstract
:1. Introduction
2. Results
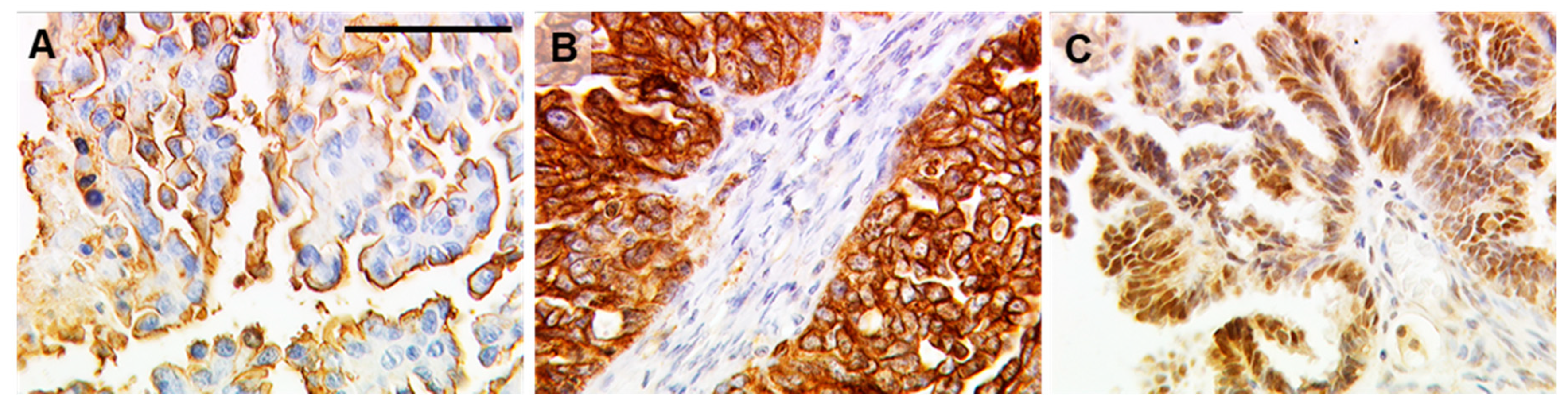
2.1. TA-MUC1 as Stained by Gatipotuzumab Correlates with Estrogen Receptor Expression
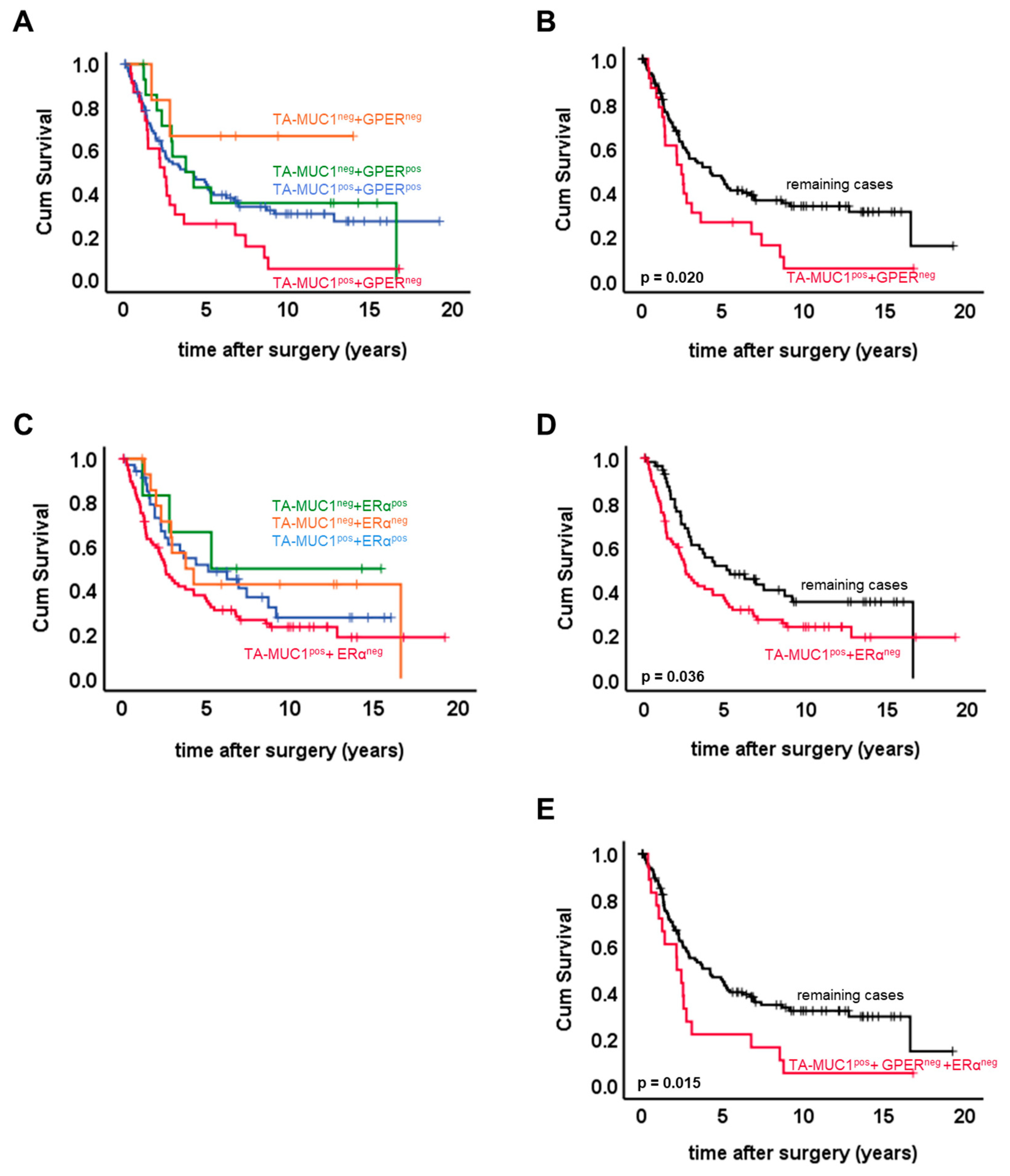
2.2. Combined Expression Patterns of TA-MUC1 and Estrogen Receptors
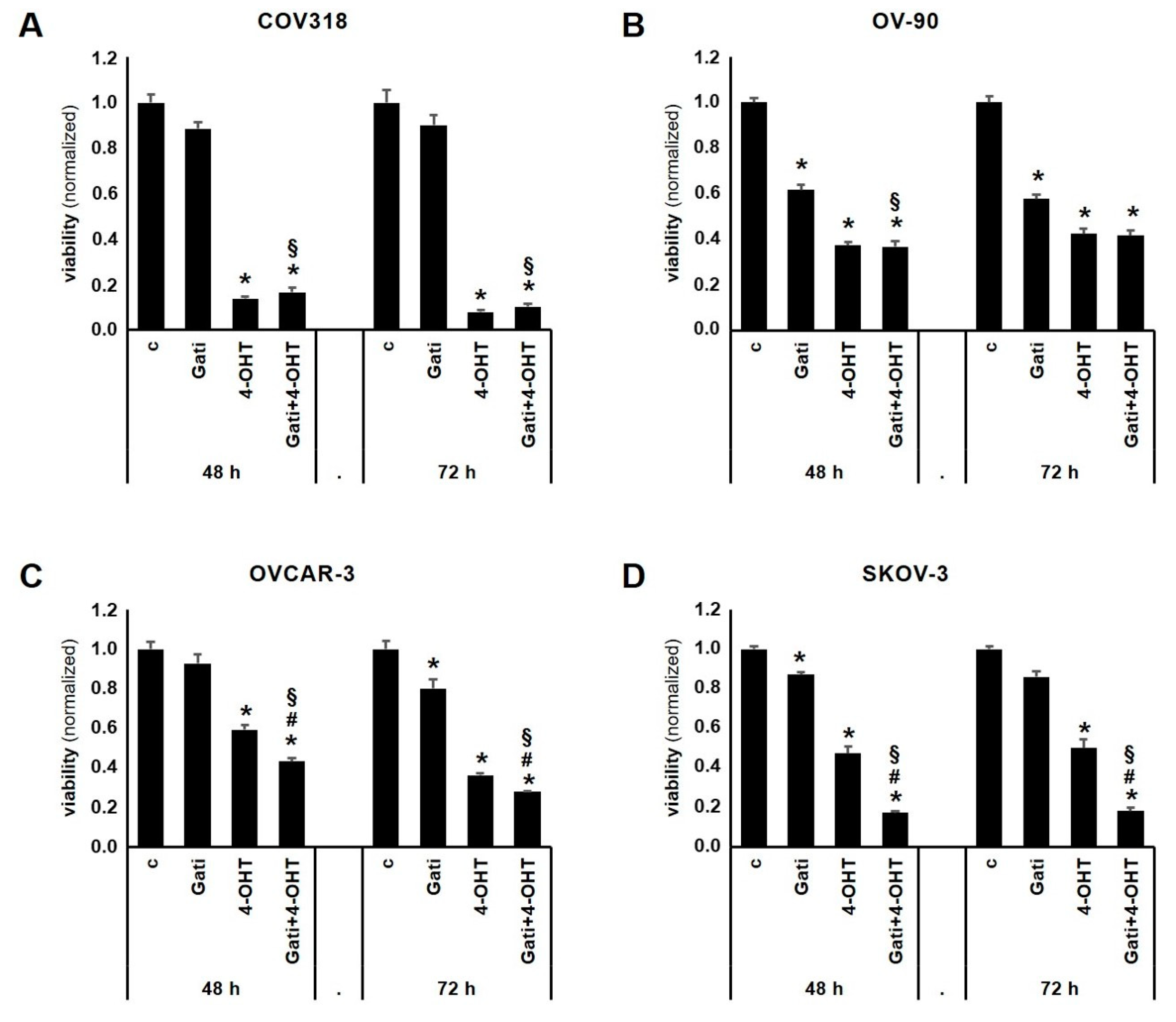
2.3. Combination of Tamoxifen and Gatipotuzumab Reduces Viability of OC Cell Lines
3. Discussion
4. Materials and Methods
4.1. Study Panel
4.2. Ethical Approval
4.3. Immunohistochemistry
4.4. Cell Culture
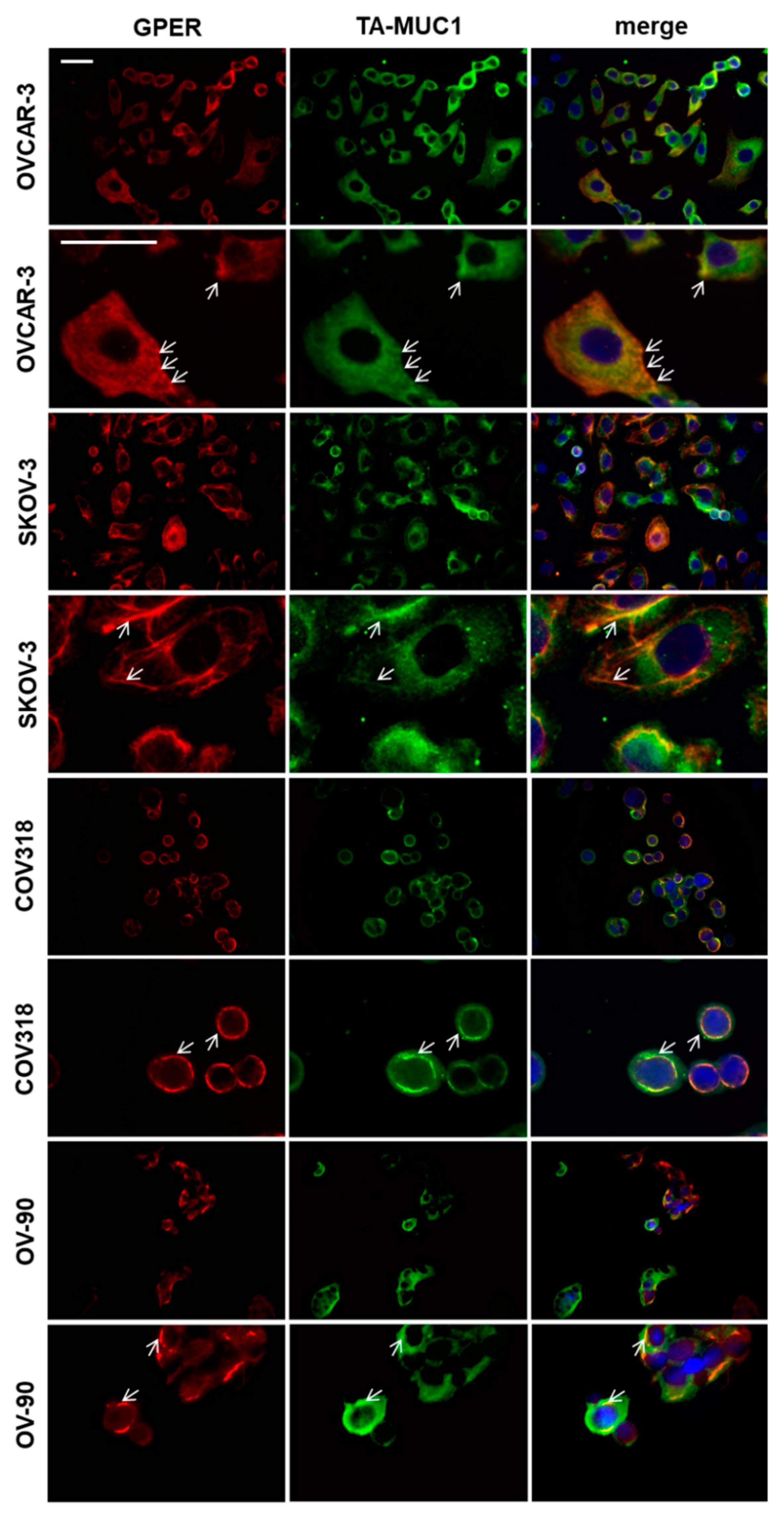
4.5. Double Immuno-Fluorescence
4.6. Determination of Cell Viability Using Water Soluble Tetrazolium (WST-1 Assay)
4.7. Statistical Analysis Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 4-OHT | 4-hydroxy-Tamoxifen |
| ER | estrogen receptor |
| FBS | fetal bovine serum |
| FFPE | formalin-fixed, paraffin-embedded |
| FIGO | Fédération Internationale de Gynécologie et d’Obstétrique |
| Gati | Gatipotuzumab |
| GPER | G-protein-coupled estrogen receptor |
| IHC | immunohistochemistry |
| MUC1 | mucin-1 |
| OC | ovarian cancer |
| OS | overall survival |
| PBS | phosphate buffered saline |
| REMARK | Reporting recommendations for tumor marker prognostic studies |
| TA-MUC1 | tumor-associated mucin-1 |
References
- Cao, Y.; Blohm, D.; Ghadimi, B.M.; Stosiek, P.; Xing, P.X.; Karsten, U. Mucins (MUC1 and MUC3) of gastrointestinal and breast epithelia reveal different and heterogeneous tumor-associated aberrations in glycosylation. J. Histochem. Cytochem. 1997, 45, 1547–1557. [Google Scholar] [CrossRef] [PubMed]
- Danielczyk, A.; Stahn, R.; Faulstich, D.; Loffler, A.; Marten, A.; Karsten, U.; Goletz, S. PankoMab: A potent new generation anti-tumour MUC1 antibody. Cancer Immunol. Immunother. 2006, 55, 1337–1347. [Google Scholar] [CrossRef] [PubMed]
- Karsten, U.; von Mensdorff-Pouilly, S.; Goletz, S. What makes MUC1 a tumor antigen? Tumour Biol. 2005, 26, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Karsten, U.; Serttas, N.; Paulsen, H.; Danielczyk, A.; Goletz, S. Binding patterns of DTR-specific antibodies reveal a glycosylation-conditioned tumor-specific epitope of the epithelial mucin (MUC1). Glycobiology 2004, 14, 681–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.N.; Karsten, U.; Goletz, S.; Cao, Y. Reactivity of a humanized antibody (hPankoMab) towards a tumor-related MUC1 epitope (TA-MUC1) with various human carcinomas. Pathol. Res. Pract. 2010, 206, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Kuemmel, A.; Single, K.; Bittinger, F.; Faldum, A.; Schmidt, L.H.; Sebastian, M.; Micke, P.; Taube, C.; Buhl, R.; Wiewrodt, R. TA-MUC1 epitope in non-small cell lung cancer. Lung Cancer 2009, 63, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Dian, D.; Lenhard, M.; Mayr, D.; Heublein, S.; Karsten, U.; Goletz, S.; Kuhn, C.; Wiest, I.; Friese, K.; Weissenbacher, T.; et al. Staining of MUC1 in ovarian cancer tissues with PankoMab-GEX detecting the tumour-associated epitope, TA-MUC1, as compared to antibodies HMFG-1 and 115D8. Histol. Histopathol. 2013, 28, 239–244. [Google Scholar] [PubMed]
- Fiedler, W.; DeDosso, S.; Cresta, S.; Weidmann, J.; Tessari, A.; Salzberg, M.; Dietrich, B.; Baumeister, H.; Goletz, S.; Gianni, L.; et al. A phase I study of PankoMab-GEX, a humanised glyco-optimised monoclonal antibody to a novel tumour-specific MUC1 glycopeptide epitope in patients with advanced carcinomas. Eur. J. Cancer 2016, 63, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Ledermann, J.; Sehouli, J.; Zurawski, B.; Raspagliesi, F.; De Giorgi, U.; Banerjee, S.; Arranz Arija, J.; Romeo Marin, M.; Lisyanskaya, A.; Póka, R.L.; et al. LBA41A double-blind, placebo-controlled, randomized, phase 2 study to evaluate the efficacy and safety of switch maintenance therapy with the anti-TA-MUC1 antibody PankoMab-GEX after chemotherapy in patients with recurrent epithelial ovarian carcinoma. Ann. Oncol. 2017, 28 (Suppl. 5), mdx440.035. [Google Scholar] [CrossRef]
- Hisatsune, A.; Nakayama, H.; Kawasaki, M.; Horie, I.; Miyata, T.; Isohama, Y.; Kim, K.C.; Katsuki, H. Anti-MUC1 antibody inhibits EGF receptor signaling in cancer cells. Biochem. Biophys. Res. Commun. 2011, 405, 377–381. [Google Scholar] [CrossRef]
- Wang, L.; Chen, H.; Liu, F.; Madigan, M.C.; Power, C.A.; Hao, J.; Patterson, K.I.; Pourgholami, M.H.; O’Brien, P.M.; Perkins, A.C.; et al. Monoclonal antibody targeting MUC1 and increasing sensitivity to docetaxel as a novel strategy in treating human epithelial ovarian cancer. Cancer Lett. 2011, 300, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.K.; Murray, J.; Parker, J.; Finke, L.; Miles, D. Humoral immune-response to naturally occurring STn in metastatic breast cancer patients (MBC pts) treated with STn-KLH vaccine. J. Clin. Oncol. 2004, 22 (Suppl. 14), 2547. [Google Scholar] [CrossRef]
- Ibrahim, N.K.; Murray, J.L.; Zhou, D.; Mittendorf, E.A.; Sample, D.; Tautchin, M.; Miles, D. Survival Advantage in Patients with Metastatic Breast Cancer Receiving Endocrine Therapy plus Sialyl Tn-KLH Vaccine: Post Hoc Analysis of a Large Randomized Trial. J. Cancer 2013, 4, 577–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmberg, L.A.; Sandmaier, B.M. Vaccination with Theratope (STn-KLH) as treatment for breast cancer. Expert Rev. Vaccines 2004, 3, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Merikhian, P.; Ghadirian, R.; Farahmand, L.; Mansouri, S.; Majidzadeh, A.K. MUC1 induces tamoxifen resistance in estrogen receptor-positive breast cancer. Expert Rev. Anticancer Ther. 2017, 17, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Ojo, D.; Lin, X.; Wu, Y.; Cockburn, J.; Bane, A.; Tang, D. Polycomb complex protein BMI1 confers resistance to tamoxifen in estrogen receptor positive breast cancer. Cancer Lett. 2018, 426, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Ince, T.A.; Sousa, A.D.; Jones, M.A.; Harrell, J.C.; Agoston, E.S.; Krohn, M.; Selfors, L.M.; Liu, W.; Chen, K.; Yong, M.; et al. Characterization of twenty-five ovarian tumour cell lines that phenocopy primary tumours. Nat. Commun. 2015, 6, 7419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, V.M.; Castro, C.M.; Chung, J.; Miller, N.C.; Ullal, A.V.; Castano, M.D.; Penson, R.T.; Lee, H.; Birrer, M.J.; Weissleder, R. Ascites analysis by a microfluidic chip allows tumor-cell profiling. Proc. Natl. Acad. Sci. USA 2013, 110, E4978–E4986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klijn, C.; Durinck, S.; Stawiski, E.W.; Haverty, P.M.; Jiang, Z.; Liu, H.; Degenhardt, J.; Mayba, O.; Gnad, F.; Liu, J.; et al. A comprehensive transcriptional portrait of human cancer cell lines. Nat. Biotechnol. 2015, 33, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Kharbanda, A.; Rajabi, H.; Jin, C.; Raina, D.; Kufe, D. Oncogenic MUC1-C promotes tamoxifen resistance in human breast cancer. Mol. Cancer Res. 2013, 11, 714–723. [Google Scholar] [CrossRef]
- Wang, H.; Lillehoj, E.P.; Kim, K.C. MUC1 tyrosine phosphorylation activates the extracellular signal-regulated kinase. Biochem. Biophys. Res. Commun. 2004, 321, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Liao, X.; Lv, Y.; Pang, Z.; Wang, Y.; Li, Q.; Liao, Y.; Ye, Q.; Chen, G.; Zhao, K.; et al. MUC1 induces acquired chemoresistance by upregulating ABCB1 in EGFR-dependent manner. Cell Death Dis. 2017, 8, e2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, B.J.; Bowser, J.L.; Broaddus, R.R.; Carson, D.D. MUC1 stimulates EGFR expression and function in endometrial cancer. Oncotarget 2016, 7, 32796–32809. [Google Scholar] [CrossRef] [Green Version]
- Vivacqua, A.; Lappano, R.; De Marco, P.; Sisci, D.; Aquila, S.; De Amicis, F.; Fuqua, S.A.; Andò, S.; Maggiolini, M. G protein-coupled receptor 30 expression is up-regulated by EGF and TGF alpha in estrogen receptor alpha-positive cancer cells. Mol. Endocrinol. 2009, 23, 1815–1826. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wu, Y.Z.; Zhang, Y.M.; Ji, X.H.; Hao, Q. Activation of G-protein coupled estrogen receptor inhibits the proliferation of cervical cancer cells via sustained activation of ERK1/2. Cell Biochem. Funct. 2015, 33, 134–142. [Google Scholar] [CrossRef]
- Chandra, V.; Fatima, I.; Saxena, R.; Hussain, M.K.; Hajela, K.; Sankhwar, P.; Roy, B.G.; Chandna, S.; Dwivedi, A. Anti-tumorigenic action of 2-[piperidinoethoxyphenyl]-3-[4-hydroxyphenyl]-2H-benzo(b)pyran: Evidence for involvement of GPR30/EGFR signaling pathway. Gynecol. Oncol. 2013, 129, 433–442. [Google Scholar] [CrossRef]
- Noske, A.; Schwabe, M.; Weichert, W.; Darb-Esfahani, S.; Buckendahl, A.C.; Sehouli, J.; Braicu, E.I.; Budczies, J.; Dietel, M.; Denkert, C. An intracellular targeted antibody detects EGFR as an independent prognostic factor in ovarian carcinomas. BMC Cancer 2011, 11, 294. [Google Scholar] [CrossRef]
- Vivacqua, A.; Bonofiglio, D.; Recchia, A.G.; Musti, A.M.; Picard, D.; Andò, S.; Maggiolini, M. The G protein-coupled receptor GPR30 mediates the proliferative effects induced by 17β-estradiol and hydroxytamoxifen in endometrial cancer cells. Mol. Endocrinol. 2006, 20, 631–646. [Google Scholar] [CrossRef]
- Heublein, S.; Mayr, D.; Vrekoussis, T.; Friese, K.; Hofmann, S.S.; Jeschke, U.; Lenhard, M. The G-protein coupled estrogen receptor (GPER/GPR30) is a gonadotropin receptor dependent positive prognosticator in ovarian carcinoma patients. PLoS ONE 2013, 8, e71791. [Google Scholar] [CrossRef]
- Engelstaedter, V.; Heublein, S.; Schumacher, A.L.; Lenhard, M.; Engelstaedter, H.; Andergassen, U.; Guenthner-Biller, M.; Kuhn, C.; Rack, B.; Kupka, M.; et al. Mucin-1 and its relation to grade, stage and survival in ovarian carcinoma patients. BMC Cancer 2012, 12, 600. [Google Scholar] [CrossRef]
- Lenhard, M.; Tereza, L.; Heublein, S.; Ditsch, N.; Himsl, I.; Mayr, D.; Friese, K.; Jeschke, U. Steroid hormone receptor expression in ovarian cancer: Progesterone receptor B as prognostic marker for patient survival. BMC Cancer 2012, 12, 553. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, M.; Tsvilina, A.; Schumacher, L.; Kupka, M.; Ditsch, N.; Mayr, D.; Friese, K.; Jeschke, U. Human chorionic gonadotropin and its relation to grade, stage and patient survival in ovarian cancer. BMC Cancer 2012, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, M.; Lennerová, T.; Ditsch, N.; Kahlert, S.; Friese, K.; Mayr, D.; Jeschke, U. Opposed roles of follicle-stimulating hormone and luteinizing hormone receptors in ovarian cancer survival. Histopathology 2011, 58, 990–994. [Google Scholar] [CrossRef] [PubMed]
- Scholz, C.; Heublein, S.; Lenhard, M.; Friese, K.; Mayr, D.; Jeschke, U. Glycodelin A is a prognostic marker to predict poor outcome in advanced stage ovarian cancer patients. BMC Res. Notes 2012, 5, 551. [Google Scholar] [CrossRef] [PubMed]
- McShane, L.M.; Altman, D.G.; Sauerbrei, W.; Taube, S.E.; Gion, M.; Clark, G.M. Reporting recommendations for tumor marker prognostic studies (REMARK). J. Natl. Cancer Inst. 2005, 97, 1180–1184. [Google Scholar] [CrossRef] [PubMed]
- Heublein, S.; Mayr, D.; Egger, M.; Karsten, U.; Goletz, S.; Angele, M.; Gallwas, J.; Jeschke, U.; Ditsch, N. Immunoreactivity of the fully humanized therapeutic antibody PankoMab-GEX is an independent prognostic marker for breast cancer patients. J. Exp. Clin. Cancer Res. 2015, 34, 50. [Google Scholar] [CrossRef] [PubMed]
- Dian, D.; Janni, W.; Kuhn, C.; Mayr, D.; Karsten, U.; Mylonas, I.; Friese, K.; Jeschke, U. Evaluation of a novel anti-mucin 1 (MUC1) antibody (PankoMab) as a potential diagnostic tool in human ductal breast cancer; comparison with two established antibodies. Onkologie 2009, 32, 238–244. [Google Scholar] [CrossRef]
- Scholz, C.; Toth, B.; Barthell, E.; Mylonas, I.; Weissenbacher, T.; Friese, K.; Jeschke, U. Immunohistochemical expression of glycodelin in breast cancer correlates with estrogen-receptor alpha and progesterone-receptor A positivity. Hist. Histopathol. 2009, 24, 467–471. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cases with Data Available (n) | Characteristic | Category | n or median | % |
|---|---|---|---|---|
| 138 | Histology | serous | 95 | 68.8% |
| mucinous | 12 | 8.7% | ||
| endometrioid | 20 | 14.5% | ||
| clear cell | 11 | 8.0% | ||
| 133 | FIGO | I + II | 40 | 30.1% |
| III + IV | 93 | 69.9% | ||
| 84 | pN | neg. | 37 | 44.0% |
| pos. | 47 | 56.0% | ||
| 126 | Grade | 1 | 31 | 24.6% |
| 2 + 3 | 95 | 75.4% | ||
| 137 | age | ≤60 y | 73 | 53.3% |
| >60 y | 64 | 46.7% |
| GPCRs | Nuclear Steroid Hormone Receptors | Glycodelin | MUC1 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GPER | LHR | FSHR | ERα | ERβ | PRA | PRB | Gd C15 | Gd Q13 | GdA | 115D8 | HMFG1 | VU3C6 | VU4H5 | HER2 | ||
| TA MUC1 | c | 0.177 | 0.095 | 0.087 | 0.230 | 0.131 | 0.100 | 0.009 | 0.219 | 0.214 | 0.066 | 0.366 | 0.458 | 0.479 | 0.353 | 0.012 |
| p | 0.038 | 0.269 | 0.318 | 0.007 | 0.127 | 0.247 | 0.914 | 0.010 | 0.012 | 0.442 | <0.001 | <0.001 | <0.001 | <0.001 | 0.885 | |
| n | 138 | 136 | 134 | 138 | 136 | 135 | 137 | 137 | 137 | 137 | 133 | 136 | 136 | 134 | 136 | |
| TA-MUC1/GPER | TA-MUC1/ER | TA-MUC1/GPER/ER | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Remaining | TA-MUC1pos. + GPERneg. | p | Remaining | TA-MUC1pos + ERαneg | p | Remaining | TA-MUC1pos + ERαneg + GPERneg | p | ||
| Histology | other | 37 | 6 | ns | 24 | 19 | 0.037 | 38 | 5 | ns |
| serous | 78 | 17 | 35 | 60 | 82 | 13 | ||||
| FIGO | I + II | 39 | 1 | 0.004 | 20 | 20 | ns | 40 | 0 | 0.004 |
| III + IV | 72 | 21 | 38 | 55 | 76 | 17 | ||||
| pN | neg. | 35 | 2 | 0.024 | 18 | 19 | ns | 36 | 1 | 0.021 |
| pos. | 36 | 11 | 18 | 29 | 38 | 9 | ||||
| Grade | 31 | 0 | 0.004 | 19 | 12 | 0.007 | 31 | 0 | 0.011 | |
| 74 | 21 | 32 | 63 | 78 | 17 | |||||
| Age ≤ 60 y | 64 | 9 | ns | 30 | 43 | ns | 66 | 7 | ns | |
| Age > 60 y | 50 | 14 | 29 | 35 | 53 | 11 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heublein, S.; Page, S.; Mayr, D.; Schmoeckel, E.; Trillsch, F.; Marmé, F.; Mahner, S.; Jeschke, U.; Vattai, A. Potential Interplay of the Gatipotuzumab Epitope TA-MUC1 and Estrogen Receptors in Ovarian Cancer. Int. J. Mol. Sci. 2019, 20, 295. https://doi.org/10.3390/ijms20020295
Heublein S, Page S, Mayr D, Schmoeckel E, Trillsch F, Marmé F, Mahner S, Jeschke U, Vattai A. Potential Interplay of the Gatipotuzumab Epitope TA-MUC1 and Estrogen Receptors in Ovarian Cancer. International Journal of Molecular Sciences. 2019; 20(2):295. https://doi.org/10.3390/ijms20020295
Chicago/Turabian StyleHeublein, Sabine, Sabina Page, Doris Mayr, Elisa Schmoeckel, Fabian Trillsch, Frederik Marmé, Sven Mahner, Udo Jeschke, and Aurelia Vattai. 2019. "Potential Interplay of the Gatipotuzumab Epitope TA-MUC1 and Estrogen Receptors in Ovarian Cancer" International Journal of Molecular Sciences 20, no. 2: 295. https://doi.org/10.3390/ijms20020295
APA StyleHeublein, S., Page, S., Mayr, D., Schmoeckel, E., Trillsch, F., Marmé, F., Mahner, S., Jeschke, U., & Vattai, A. (2019). Potential Interplay of the Gatipotuzumab Epitope TA-MUC1 and Estrogen Receptors in Ovarian Cancer. International Journal of Molecular Sciences, 20(2), 295. https://doi.org/10.3390/ijms20020295