Epigenetic Mechanisms Influencing Epithelial to Mesenchymal Transition in Bladder Cancer





Abstract
:1. Introduction
1.1. Bladder Cancer
1.2. Epigenetics
1.3. Epithelial to Mesenchymal Transition
1.4. Influence of EMT Major Players in Bladder Cancer
2. Epigenetic Enzymes and Mechanisms Altering EMT in Bladder Cancer
2.1. Protein-Coding Genes
2.2. Non-Coding RNAs
3. Conclusions
Funding
Conflicts of Interest
Abbreviations
| 5AZA | 5-Aza-2’-deoxycytidine |
| BlCa | Bladder cancer |
| CAF | Cancer associated fibroblast |
| ceRNA | Competing endogenous RNA |
| circRNA | Circular RNA |
| DNMT | DNA methyltransferase |
| dUb | Histone deubiquitinase |
| ECAD | Cadherin 1, E-Cadherin |
| ELF5 | E74 Like ETS Transcription Factor 5 |
| EMT | Epithelial to mesenchymal transition |
| EMT-TF | EMT transcription factor |
| GDF15 | Growth Differentiation Factor 15 |
| HAT | Histone acetyltransferase |
| HDAC | Histone deacetylase |
| HDM | Histone demethylase |
| HTM | Histone methyltransferase |
| KLF4 | Kruppel Like Factor 4 |
| lncRNA | Long non-coding RNA |
| MAEL | Maelstrom Spermatogenic Transposon Silencer |
| MET | Mesenchymal to epithelial transition |
| MIBC | Muscle invasive bladder cancer |
| miR | Micro RNA |
| MTSS1 | Metastasis Supressor Protein 1 |
| NCAD | Cadherin 2, N-Cadherin |
| ncRNA | Non-coding RNA |
| NMIBC | Non-muscle invasive bladder cancer |
| PRC | Polycomb repressive complex |
| PRSS8 | Serine Protease 8 |
| Snail | Snail Family Transcriptional Repressor 1 |
| Slug | Snail Family Transcriptional Repressor 2 |
| sncRNA | Small non-coding RNA |
| TET | DNA demethylase |
| UbL | Histone ubiquitin ligase |
| VIM | Vimentin |
| ZEB1 | Zync Finger E-Box Binding Homeobox 1 |
| ZEB2 | Zync Finger E-Box Binding Homeobox 2 |
References
- Wong, M.C.S.; Fung, F.D.H.; Leung, C.; Cheung, W.W.L.; Goggins, W.B.; Ng, C.F. The global epidemiology of bladder cancer: A joinpoint regression analysis of its incidence and mortality trends and projection. Sci. Rep. 2018, 8, 1129. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Tomorrow; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Antoni, S.; Ferlay, J.; Soerjomataram, I.; Znaor, A.; Jemal, A.; Bray, F. Bladder Cancer Incidence and Mortality: A Global Overview and Recent Trends. Eur. Urol. 2017, 71, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Greiman, A.K.; Rosoff, J.S.; Prasad, S.M. Association of Human Development Index with global bladder, kidney, prostate and testis cancer incidence and mortality. BJU Int. 2017, 120, 799–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, J.; Luengo-Fernandez, R.; Sullivan, R.; Witjes, J.A. Economic Burden of Bladder Cancer Across the European Union. Eur. Urol. 2016, 69, 438–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanli, O.; Dobruch, J.; Knowles, M.A.; Burger, M.; Alemozaffar, M.; Nielsen, M.E.; Lotan, Y. Bladder cancer. Nat. Rev. Dis. Primers 2017, 3, 17022. [Google Scholar] [CrossRef] [Green Version]
- Moch, H.; Ulbright, T.; Humphrey, P.; Reuter, V. WHO Classification of Tumours of the Urinary System and Male Genital Organs, 4th ed.; IARC: Lyon, France, 2016. [Google Scholar]
- Meissner, A. Epigenetic modifications in pluripotent and differentiated cells. Nat. Biotechnol. 2010, 28, 1079–1088. [Google Scholar] [CrossRef]
- Skinner, M.K. Role of epigenetics in developmental biology and transgenerational inheritance. Birth Defects Res. C Embryo Today 2011, 93, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Saitou, M.; Kagiwada, S.; Kurimoto, K. Epigenetic reprogramming in mouse pre-implantation development and primordial germ cells. Development 2012, 139, 15–31. [Google Scholar] [CrossRef]
- Kelly, A.D.; Issa, J.J. The promise of epigenetic therapy: Reprogramming the cancer epigenome. Curr. Opin. Genet. Dev. 2017, 42, 68–77. [Google Scholar] [CrossRef]
- Baylin, S.B.; Jones, P.A. Epigenetic Determinants of Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef]
- Esteller, M. Epigenetics in cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Mossanen, M.; Gore, J.L. The burden of bladder cancer care: Direct and indirect costs. Curr. Opin. Urol. 2014, 24, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Kojima, T.; Kawai, K.; Miyazaki, J.; Nishiyama, H. Biomarkers for precision medicine in bladder cancer. Int. J. Clin. Oncol. 2017, 22, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Dudziec, E.; Goepel, J.R.; Catto, J.W. Global epigenetic profiling in bladder cancer. Epigenomics 2011, 3, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Schulz, W.A.; Koutsogiannouli, E.A.; Niegisch, G.; Hoffmann, M.J. Epigenetics of urothelial carcinoma. Methods Mol. Biol. 2015, 1238, 183–215. [Google Scholar] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieto, M.A.; Huang, R.Y.; Jackson, R.A.; Thiery, J.P. Emt: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Tam, W.L.; Weinberg, R.A. The epigenetics of epithelial-mesenchymal plasticity in cancer. Nat. Med. 2013, 19, 1438–1449. [Google Scholar] [CrossRef] [Green Version]
- Thiery, J.P.; Sleeman, J.P. Complex networks orchestrate epithelial-mesenchymal transitions. Nat. Rev. Mol. Cell Biol. 2006, 7, 131–142. [Google Scholar] [CrossRef]
- De Craene, B.; Berx, G. Regulatory networks defining EMT during cancer initiation and progression. Nat. Rev. Cancer 2013, 13, 97–110. [Google Scholar] [CrossRef]
- Van Roy, F.; Berx, G. The cell-cell adhesion molecule E-cadherin. Cell. Mol. Life Sci. 2008, 65, 3756–3788. [Google Scholar] [CrossRef] [PubMed]
- Bracken, C.P.; Gregory, P.A.; Kolesnikoff, N.; Bert, A.G.; Wang, J.; Shannon, M.F.; Goodall, G.J. A double-negative feedback loop between ZEB1-SIP1 and the microRNA-200 family regulates epithelial-mesenchymal transition. Cancer Res. 2008, 68, 7846–7854. [Google Scholar] [CrossRef] [PubMed]
- Chaffer, C.L.; Brennan, J.P.; Slavin, J.L.; Blick, T.; Thompson, E.W.; Williams, E.D. Mesenchymal-to-epithelial transition facilitates bladder cancer metastasis: Role of fibroblast growth factor receptor-2. Cancer Res. 2006, 66, 11271–11278. [Google Scholar] [CrossRef]
- Choi, J.; Park, S.Y.; Joo, C.K. Transforming growth factor-beta1 represses E-cadherin production via slug expression in lens epithelial cells. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2708–2718. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Yu, J.; Dhanasekaran, S.M.; Kim, J.H.; Mani, R.S.; Tomlins, S.A.; Mehra, R.; Laxman, B.; Cao, X.; Yu, J.; et al. Repression of E-cadherin by the polycomb group protein EZH2 in cancer. Oncogene 2008, 27, 7274–7284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, N.; Tiwari, V.K.; Waldmeier, L.; Balwierz, P.J.; Arnold, P.; Pachkov, M.; Meyer-Schaller, N.; Schubeler, D.; van Nimwegen, E.; Christofori, G. Sox4 is a master regulator of epithelial-mesenchymal transition by controlling Ezh2 expression and epigenetic reprogramming. Cancer Cell 2013, 23, 768–783. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xu, Z.; Zhong, L.; Wang, H.; Jiang, S.; Long, Q.; Xu, J.; Guo, J. Enhancer of zeste homolog 2 (EZH2) promotes tumour cell migration and invasion via epigenetic repression of E-cadherin in renal cell carcinoma. BJU Int. 2016, 117, 351–362. [Google Scholar] [CrossRef]
- Liu, X.; Wang, C.; Chen, Z.; Jin, Y.; Wang, Y.; Kolokythas, A.; Dai, Y.; Zhou, X. MicroRNA-138 suppresses epithelial-mesenchymal transition in squamous cell carcinoma cell lines. Biochem. J. 2011, 440, 23–31. [Google Scholar] [CrossRef]
- Luo, M.; Li, Z.; Wang, W.; Zeng, Y.; Liu, Z.; Qiu, J. Long non-coding RNA H19 increases bladder cancer metastasis by associating with EZH2 and inhibiting E-cadherin expression. Cancer Lett. 2013, 333, 213–221. [Google Scholar] [CrossRef]
- Kottakis, F.; Polytarchou, C.; Foltopoulou, P.; Sanidas, I.; Kampranis, S.C.; Tsichlis, P.N. FGF-2 regulates cell proliferation, migration, and angiogenesis through an NDY1/KDM2B-miR-101-EZH2 pathway. Mol. Cell 2011, 43, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M.; Liang, G.; Liu, C.C.; Wolff, E.M.; Tsai, Y.C.; Ye, W.; Zhou, X.; Jones, P.A. The putative tumor suppressor microRNA-101 modulates the cancer epigenome by repressing the polycomb group protein EZH2. Cancer Res. 2009, 69, 2623–2629. [Google Scholar] [CrossRef] [PubMed]
- Varambally, S.; Cao, Q.; Mani, R.S.; Shankar, S.; Wang, X.; Ateeq, B.; Laxman, B.; Cao, X.; Jing, X.; Ramnarayanan, K.; et al. Genomic loss of microRNA-101 leads to overexpression of histone methyltransferase EZH2 in cancer. Science 2008, 322, 1695–1699. [Google Scholar] [CrossRef] [PubMed]
- McNiel, E.A.; Tsichlis, P.N. Analyses of publicly available genomics resources define FGF-2-expressing bladder carcinomas as EMT-prone, proliferative tumors with low mutation rates and high expression of CTLA-4, PD-1 and PD-L1. Signal Transduct. Target. Ther. 2017, 2. [Google Scholar] [CrossRef]
- Lee, S.R.; Roh, Y.G.; Kim, S.K.; Lee, J.S.; Seol, S.Y.; Lee, H.H.; Kim, W.T.; Kim, W.J.; Heo, J.; Cha, H.J.; et al. Activation of EZH2 and SUZ12 Regulated by E2F1 Predicts the Disease Progression and Aggressive Characteristics of Bladder Cancer. Clin. Cancer Res. 2015, 21, 5391–5403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Liu, D.; Tao, D.; Xiang, W.; Xiao, X.; Wang, M.; Wang, L.; Luo, G.; Li, Y.; Zeng, F.; et al. BRD4 Regulates EZH2 Transcription through Upregulation of C-MYC and Represents a Novel Therapeutic Target in Bladder Cancer. Mol. Cancer Ther. 2016, 15, 1029–1042. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.A.; Varambally, S.; Arend, R.C. Histone Methyltransferase EZH2: A Therapeutic Target for Ovarian Cancer. Mol. Cancer Ther. 2018, 17, 591–602. [Google Scholar] [CrossRef]
- ORIOn-E: A Study Evaluating CPI-1205 in Patients with Advanced Solid Tumors. Available online: https://clinicaltrials.gov/show/NCT03525795 (accessed on 03 December 2018).
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [Green Version]
- Leyvraz, C.; Charles, R.P.; Rubera, I.; Guitard, M.; Rotman, S.; Breiden, B.; Sandhoff, K.; Hummler, E. The epidermal barrier function is dependent on the serine protease CAP1/Prss8. J. Cell Biol. 2005, 170, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.M.; Verity, N.J.; Chai, K.X. Loss of prostasin (PRSS8) in human bladder transitional cell carcinoma cell lines is associated with epithelial-mesenchymal transition (EMT). BMC Cancer 2009, 9, 377. [Google Scholar] [CrossRef]
- Hemberger, M.; Udayashankar, R.; Tesar, P.; Moore, H.; Burton, G.J. ELF5-enforced transcriptional networks define an epigenetically regulated trophoblast stem cell compartment in the human placenta. Hum. Mol. Genet. 2010, 19, 2456–2467. [Google Scholar] [CrossRef] [Green Version]
- Ng, R.K.; Dean, W.; Dawson, C.; Lucifero, D.; Madeja, Z.; Reik, W.; Hemberger, M. Epigenetic restriction of embryonic cell lineage fate by methylation of Elf5. Nat. Cell Biol. 2008, 10, 1280–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Cao, X.; Liang, X.; Zhang, X.; Zhang, W.; Sun, G.; Wang, D. Epigenetic regulation of Elf5 is associated with epithelial-mesenchymal transition in urothelial cancer. PLoS ONE 2015, 10, e0117510. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Zhang, J.X.; Jiang, L.J.; Wang, F.W.; Liu, L.L.; Liao, Y.J.; Jin, X.H.; Chen, W.H.; Chen, X.; Guo, S.J.; et al. Overexpression of maelstrom promotes bladder urothelial carcinoma cell aggressiveness by epigenetically downregulating MTSS1 through DNMT3B. Oncogene 2016, 35, 6281–6292. [Google Scholar] [CrossRef] [PubMed]
- Tsui, K.H.; Hsu, S.Y.; Chung, L.C.; Lin, Y.H.; Feng, T.H.; Lee, T.Y.; Chang, P.L.; Juang, H.H. Growth differentiation factor-15: A p53- and demethylation-upregulating gene represses cell proliferation, invasion, and tumorigenesis in bladder carcinoma cells. Sci. Rep. 2015, 5, 12870. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, J.; Xiao, W.; Xia, D.; Lang, B.; Wang, T.; Guo, X.; Hu, Z.; Ye, Z.; Xu, H. Epigenetic inactivation of KLF4 is associated with urothelial cancer progression and early recurrence. J. Urol. 2014, 191, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, J.; Zhu, Y.; Xie, B.; Wang, X.; Wang, S.; Xie, H.; Yan, H.; Ying, Y.; Lin, Y.; et al. CRISPR-ON-Mediated KLF4 overexpression inhibits the proliferation, migration and invasion of urothelial bladder cancer in vitro and in vivo. Oncotarget 2017, 8, 102078–102087. [Google Scholar] [CrossRef] [Green Version]
- Costa, V.L.; Henrique, R.; Danielsen, S.A.; Duarte-Pereira, S.; Eknaes, M.; Skotheim, R.I.; Rodrigues, A.; Magalhaes, J.S.; Oliveira, J.; Lothe, R.A.; et al. Three epigenetic biomarkers, GDF15, TMEFF2, and VIM, accurately predict bladder cancer from DNA-based analyses of urine samples. Clin. Cancer Res. 2010, 16, 5842–5851. [Google Scholar] [CrossRef]
- Monteiro-Reis, S.; Leca, L.; Almeida, M.; Antunes, L.; Monteiro, P.; Dias, P.C.; Morais, A.; Oliveira, J.; Henrique, R.; Jeronimo, C. Accurate detection of upper tract urothelial carcinoma in tissue and urine by means of quantitative GDF15, TMEFF2 and VIM promoter methylation. Eur. J. Cancer 2014, 50, 226–233. [Google Scholar] [CrossRef]
- Wei, D.; Gong, W.; Kanai, M.; Schlunk, C.; Wang, L.; Yao, J.C.; Wu, T.T.; Huang, S.; Xie, K. Drastic down-regulation of Kruppel-like factor 4 expression is critical in human gastric cancer development and progression. Cancer Res. 2005, 65, 2746–2754. [Google Scholar] [CrossRef]
- Yang, Y.; Goldstein, B.G.; Chao, H.H.; Katz, J.P. KLF4 and KLF5 regulate proliferation, apoptosis and invasion in esophageal cancer cells. Cancer Biol. Ther. 2005, 4, 1216–1221. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Place, R.F.; Huang, V.; Wang, X.; Noonan, E.J.; Magyar, C.E.; Huang, J.; Li, L.C. Prognostic value and function of KLF4 in prostate cancer: RNAa and vector-mediated overexpression identify KLF4 as an inhibitor of tumor cell growth and migration. Cancer Res. 2010, 70, 10182–10191. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Hofstetter, W.L.; Li, H.; Zhou, Y.; He, Y.; Pataer, A.; Wang, L.; Xie, K.; Swisher, S.G.; Fang, B. Putative tumor-suppressive function of Kruppel-like factor 4 in primary lung carcinoma. Clin. Cancer Res. 2009, 15, 5688–5695. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Tao, Y.; Gao, X.; Zhang, L.; Li, X.; Zou, W.; Ruan, K.; Wang, F.; Xu, G.L.; Hu, R. A CRISPR-based approach for targeted DNA demethylation. Cell Discov. 2016, 2, 16009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Kazemier, H.G.; de Groote, M.L.; Ruiters, M.H.; Xu, G.L.; Rots, M.G. Induced DNA demethylation by targeting Ten-Eleven Translocation 2 to the human ICAM-1 promoter. Nucleic Acids Res. 2014, 42, 1563–1574. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Rinn, J.L. Discovery and annotation of long noncoding RNAs. Nat. Struct. Mol. Biol. 2015, 22, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Ramalho-Carvalho, J.; Fromm, B.; Henrique, R.; Jeronimo, C. Deciphering the function of non-coding RNAs in prostate cancer. Cancer Metastasis Rev. 2016, 35, 235–262. [Google Scholar] [CrossRef] [PubMed]
- Peschansky, V.J.; Wahlestedt, C. Non-coding RNAs as direct and indirect modulators of epigenetic regulation. Epigenetics 2014, 9, 3–12. [Google Scholar] [CrossRef]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: An expanding universe. Nat. Rev. Genet. 2009, 10, 94–108. [Google Scholar] [CrossRef]
- Falzone, L.; Candido, S.; Salemi, R.; Basile, M.S.; Scalisi, A.; McCubrey, J.A.; Torino, F.; Signorelli, S.S.; Montella, M.; Libra, M. Computational identification of microRNAs associated to both epithelial to mesenchymal transition and NGAL/MMP-9 pathways in bladder cancer. Oncotarget 2016, 7, 72758–72766. [Google Scholar] [CrossRef] [PubMed]
- Adam, L.; Zhong, M.; Choi, W.; Qi, W.; Nicoloso, M.; Arora, A.; Calin, G.; Wang, H.; Siefker-Radtke, A.; McConkey, D.; et al. miR-200 expression regulates epithelial-to-mesenchymal transition in bladder cancer cells and reverses resistance to epidermal growth factor receptor therapy. Clin. Cancer Res. 2009, 15, 5060–5072. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, S.; Lin, Y.; Chen, H.; Hu, Z.; Mao, Y.; Xu, X.; Wu, J.; Zhu, Y.; Zheng, X.; et al. MicroRNA-124-3p inhibits cell migration and invasion in bladder cancer cells by targeting ROCK1. J. Transl. Med. 2013, 11, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, M.N.; Choi, W.; Wszolek, M.F.; Navai, N.; Lee, I.L.; Nitti, G.; Wen, S.; Flores, E.R.; Siefker-Radtke, A.; Czerniak, B.; et al. The p63 protein isoform DeltaNp63alpha inhibits epithelial-mesenchymal transition in human bladder cancer cells: Role of MIR-205. J. Biol. Chem. 2013, 288, 3275–3288. [Google Scholar] [CrossRef] [PubMed]
- Majid, S.; Dar, A.A.; Saini, S.; Deng, G.; Chang, I.; Greene, K.; Tanaka, Y.; Dahiya, R.; Yamamura, S. MicroRNA-23b functions as a tumor suppressor by regulating Zeb1 in bladder cancer. PLoS ONE 2013, 8, e67686. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Qiu, M.; Tan, G.; Liang, Z.; Qin, Y.; Chen, L.; Chen, H.; Liu, J. miR-200c inhibits invasion, migration and proliferation of bladder cancer cells through down-regulation of BMI-1 and E2F3. J. Transl. Med. 2014, 12, 305. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Peng, L.; Chao, C.; Fu, B.; Wang, G.; Wang, Y.; Zhu, X. miR-451 inhibits invasion and proliferation of bladder cancer by regulating EMT. Int. J. Clin. Exp. Pathol. 2014, 7, 7653–7662. [Google Scholar]
- Zhang, S.; Zhang, C.; Liu, W.; Zheng, W.; Zhang, Y.; Wang, S.; Huang, D.; Liu, X.; Bai, Z. MicroRNA-24 upregulation inhibits proliferation, metastasis and induces apoptosis in bladder cancer cells by targeting CARMA3. Int. J. Oncol. 2015, 47, 1351–1360. [Google Scholar] [CrossRef]
- Yu, G.; Xu, K.; Xu, S.; Zhang, X.; Huang, Q.; Lang, B. MicroRNA-34a regulates cell cycle by targeting CD44 in human bladder carcinoma cells. Nan Fang Yi Ke Da Xue Xue Bao 2015, 35, 935–940. [Google Scholar]
- Yao, K.; He, L.; Gan, Y.; Zeng, Q.; Dai, Y.; Tan, J. MiR-186 suppresses the growth and metastasis of bladder cancer by targeting NSBP1. Diagn. Pathol. 2015, 10, 146. [Google Scholar] [CrossRef]
- Martinez-Fernandez, M.; Duenas, M.; Feber, A.; Segovia, C.; Garcia-Escudero, R.; Rubio, C.; Lopez-Calderon, F.F.; Diaz-Garcia, C.; Villacampa, F.; Duarte, J.; et al. A Polycomb-mir200 loop regulates clinical outcome in bladder cancer. Oncotarget 2015, 6, 42258–42275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.F.; Zeng, F.; Qi, L.; Zu, X.B.; Wang, J.; Liu, L.F.; Li, Y. Transforming growth factorβ1 induces epithelialmesenchymal transition and increased expression of matrix metalloproteinase16 via miR200b downregulation in bladder cancer cells. Mol. Med. Rep. 2014, 10, 1549–1554. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, J.; Zhao, X. miR-221 facilitates the TGFbeta1-induced epithelial-mesenchymal transition in human bladder cancer cells by targeting STMN1. BMC Urol. 2015, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.T.; Lin, W.Y.; Chang, Y.H.; Lin, P.Y.; Chen, W.C.; Chen, M.F. DNMT1-dependent suppression of microRNA424 regulates tumor progression in human bladder cancer. Oncotarget 2015, 6, 24119–24131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Li, Q.; Wang, S.; Zhang, J. miR4855p inhibits bladder cancer metastasis by targeting HMGA2. Int. J. Mol. Med. 2015, 36, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ke, C.; Ma, X.; Zhao, Q.; Yang, M.; Zhang, W.; Wang, J. MicroRNA-92 promotes invasion and chemoresistance by targeting GSK3β and activating Wnt signaling in bladder cancer cells. Tumour Biol. 2016. [Google Scholar] [CrossRef]
- Huang, J.; Wang, B.; Hui, K.; Zeng, J.; Fan, J.; Wang, X.; Hsieh, J.T.; He, D.; Wu, K. miR-92b targets DAB2IP to promote EMT in bladder cancer migration and invasion. Oncol. Rep. 2016, 36, 1693–1701. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, S.; Hu, L.; Liu, F.; Zhang, Q.; Zhang, D. miR-199a-5p suppresses human bladder cancer cell metastasis by targeting CCR7. BMC Urol. 2016, 16, 64. [Google Scholar] [CrossRef]
- Wang, X.; Liang, Z.; Xu, X.; Li, J.; Zhu, Y.; Meng, S.; Li, S.; Wang, S.; Xie, B.; Ji, A.; et al. miR-148a-3p represses proliferation and EMT by establishing regulatory circuits between ERBB3/AKT2/c-myc and DNMT1 in bladder cancer. Cell Death Dis. 2016, 7, e2503. [Google Scholar] [CrossRef]
- Wu, C.L.; Ho, J.Y.; Chou, S.C.; Yu, D.S. MiR-429 reverses epithelial-mesenchymal transition by restoring E-cadherin expression in bladder cancer. Oncotarget 2016, 7, 26593–26603. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhu, Y.; Liang, Z.; Li, S.; Xu, X.; Wang, X.; Wu, J.; Hu, Z.; Meng, S.; Liu, B.; et al. c-Met and CREB1 are involved in miR-433-mediated inhibition of the epithelial-mesenchymal transition in bladder cancer by regulating Akt/GSK-3β/Snail signaling. Cell Death Dis. 2016, 7, e2088. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, J.; Xiao, M.; Yang, J.; Zhang, N. MiR-203 Suppresses Bladder Cancer Cell Growth and Targets the Twist1. Oncol. Res. 2017, 26, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Gao, H.; Liu, S.; Chen, M.; Xu, W.; Ling, X.; Deng, X.; Rao, C. Down-regulation of microRNA-224 -inhibites growth and epithelial-to-mesenchymal transition phenotype -via modulating SUFU expression in bladder cancer cells. Int. J. Biol. Macromol. 2018, 106, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xu, X.; Meng, S.; Liang, Z.; Wang, X.; Xu, M.; Wang, S.; Li, S.; Zhu, Y.; Xie, B.; et al. MET/SMAD3/SNAIL circuit mediated by miR-323a-3p is involved in regulating epithelial-mesenchymal transition progression in bladder cancer. Cell Death Dis. 2017, 8, e3010. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Hu, X.; Liu, J.; Zhu, W.; Zhan, X.; Sun, S. MicroRNA-497 upregulation inhibits cell invasion and metastasis in T24 and BIU-87 bladder cancer cells. Mol. Med. Rep. 2017, 16, 2055–2060. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Duan, P.; Zhu, H.; Rao, D. miR-613 inhibits bladder cancer proliferation and migration through targeting SphK1. Am. J. Transl. Res. 2017, 9, 1213–1221. [Google Scholar] [PubMed]
- Xu, M.; Li, J.; Wang, X.; Meng, S.; Shen, J.; Wang, S.; Xu, X.; Xie, B.; Liu, B.; Xie, L. MiR-22 suppresses epithelial-mesenchymal transition in bladder cancer by inhibiting Snail and MAPK1/Slug/vimentin feedback loop. Cell Death Dis. 2018, 9, 209. [Google Scholar] [CrossRef]
- Mao, X.W.; Xiao, J.Q.; Li, Z.Y.; Zheng, Y.C.; Zhang, N. Effects of microRNA-135a on the epithelial-mesenchymal transition, migration and invasion of bladder cancer cells by targeting GSK3β through the Wnt/β-catenin signaling pathway. Exp. Mol. Med. 2018, 50, e429. [Google Scholar] [CrossRef]
- Huang, X.; Zhu, H.; Gao, Z.; Li, J.; Zhuang, J.; Dong, Y.; Shen, B.; Li, M.; Zhou, H.; Guo, H.; et al. Wnt7a activates canonical Wnt signaling, promotes bladder cancer cell invasion, and is suppressed by miR-370-3p. J. Biol. Chem. 2018. [Google Scholar] [CrossRef]
- Liu, W.; Qi, L.; Lv, H.; Zu, X.; Chen, M.; Wang, J.; Liu, L.; Zeng, F.; Li, Y. MiRNA-141 and miRNA-200b are closely related to invasive ability and considered as decision-making biomarkers for the extent of PLND during cystectomy. BMC Cancer 2015, 15, 92. [Google Scholar] [CrossRef]
- Yan, L.; Wang, Y.; Liang, J.; Liu, Z.; Sun, X.; Cai, K. MiR-301b promotes the proliferation, mobility, and epithelial-to-mesenchymal transition of bladder cancer cells by targeting EGR1. Biochem. Cell Biol. 2017, 95, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ge, Q.; Chen, Z.; Hu, J.; Li, F.; Ye, Z. Promoter-associated endogenous and exogenous small RNAs suppress human bladder cancer cell metastasis by activating p21 (CIP1/WAF1) expression. Tumour Biol. 2016, 37, 6589–6598. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Chen, Y.; Huang, B.; Mao, S.; Cai, K.; Wang, L.; Yao, X. Tumor-suppressing effects of microRNA-612 in bladder cancer cells by targeting malic enzyme 1 expression. Int. J. Oncol. 2018, 52, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Huang, M.; Lv, M.; He, Y.; Duan, C.; Zhang, L.; Chen, J. Circular RNA MYLK as a competing endogenous RNA promotes bladder cancer progression through modulating VEGFA/VEGFR2 signaling pathway. Cancer Lett. 2017, 403, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Zhu, J.L.; Bao, W.S.; Chen, D.K.; Huang, W.W.; Weng, Z.L. Long noncoding RNA GHET1 promotes the development of bladder cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 7196–7205. [Google Scholar]
- Berrondo, C.; Flax, J.; Kucherov, V.; Siebert, A.; Osinski, T.; Rosenberg, A.; Fucile, C.; Richheimer, S.; Beckham, C.J. Expression of the Long Non-Coding RNA HOTAIR Correlates with Disease Progression in Bladder Cancer and Is Contained in Bladder Cancer Patient Urinary Exosomes. PLoS ONE 2016, 11, e0147236. [Google Scholar] [CrossRef]
- Lv, M.; Zhong, Z.; Huang, M.; Tian, Q.; Jiang, R.; Chen, J. lncRNA H19 regulates epithelial-mesenchymal transition and metastasis of bladder cancer by miR-29b-3p as competing endogenous RNA. Biochim. Biophys. Acta 2017, 1864, 1887–1899. [Google Scholar] [CrossRef]
- Fan, Y.; Shen, B.; Tan, M.; Mu, X.; Qin, Y.; Zhang, F.; Liu, Y. TGF-beta-induced upregulation of malat1 promotes bladder cancer metastasis by associating with suz12. Clin. Cancer Res. 2014, 20, 1531–1541. [Google Scholar] [CrossRef]
- Chen, Y.; Peng, Y.; Xu, Z.; Ge, B.; Xiang, X.; Zhang, T.; Gao, L.; Shi, H.; Wang, C.; Huang, J. LncROR Promotes Bladder Cancer Cell Proliferation, Migration, and Epithelial-Mesenchymal Transition. Cell. Physiol. Biochem. 2017, 41, 2399–2410. [Google Scholar] [CrossRef] [Green Version]
- Tuo, Z.; Zhang, J.; Xue, W. LncRNA TP73-AS1 predicts the prognosis of bladder cancer patients and functions as a suppressor for bladder cancer by EMT pathway. Biochem. Biophys. Res. Commun. 2018, 499, 875–881. [Google Scholar] [CrossRef]
- Tan, J.; Qiu, K.; Li, M.; Liang, Y. Double-negative feedback loop between long non-coding RNA TUG1 and miR-145 promotes epithelial to mesenchymal transition and radioresistance in human bladder cancer cells. FEBS Lett. 2015, 589, 3175–3181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, M.; Pang, H.; Li, X.; Li, H.; Pan, J.; Chen, W. Long non-coding RNA urothelial cancer-associated 1 promotes bladder cancer cell migration and invasion by way of the hsa-miR-145-ZEB1/2-FSCN1 pathway. Cancer Sci. 2016, 107, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Chen, J.; Li, H.; Yang, Y.; Yun, H.; Yang, S.; Mao, X. LncRNA UCA1 promotes the invasion and EMT of bladder cancer cells by regulating the miR-143/HMGB1 pathway. Oncol. Lett. 2017, 14, 5556–5562. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Zhu, X.; Chen, F.; Huang, C.; Ai, K.; Wu, H.; Zhang, L.; Zhao, X. LncRNA XIST/miR-200c regulates the stemness properties and tumourigenicity of human bladder cancer stem cell-like cells. Cancer Cell Int. 2018, 18, 41. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, J.; Lu, Q.; Shen, B.; Huang, X.; Shen, L.; Zheng, X.; Huang, R.; Yan, J.; Guo, H. TGFβ1 secreted by cancer-associated fibroblasts induces epithelial-mesenchymal transition of bladder cancer cells through lncRNA-ZEB2NAT. Sci. Rep. 2015, 5, 11924. [Google Scholar] [CrossRef] [Green Version]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Shimono, Y.; Zabala, M.; Cho, R.W.; Lobo, N.; Dalerba, P.; Qian, D.; Diehn, M.; Liu, H.; Panula, S.P.; Chiao, E.; et al. Downregulation of miRNA-200c links breast cancer stem cells with normal stem cells. Cell 2009, 138, 592–603. [Google Scholar] [CrossRef]
- Leskela, S.; Leandro-Garcia, L.J.; Mendiola, M.; Barriuso, J.; Inglada-Perez, L.; Munoz, I.; Martinez-Delgado, B.; Redondo, A.; de Santiago, J.; Robledo, M.; et al. The miR-200 family controls beta-tubulin III expression and is associated with paclitaxel-based treatment response and progression-free survival in ovarian cancer patients. Endocr. Relat. Cancer 2011, 18, 85–95. [Google Scholar] [CrossRef]
- Vallejo, D.M.; Caparros, E.; Dominguez, M. Targeting Notch signalling by the conserved miR-8/200 microRNA family in development and cancer cells. EMBO J. 2011, 30, 756–769. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Park, Y.A.; Choi, J.J.; Lee, Y.Y.; Kim, C.J.; Choi, C.; Kim, T.J.; Lee, N.W.; Kim, B.G.; Bae, D.S. The expression of the miRNA-200 family in endometrial endometrioid carcinoma. Gynecol. Oncol. 2011, 120, 56–62. [Google Scholar] [CrossRef]
- Santos, M.; Martinez-Fernandez, M.; Duenas, M.; Garcia-Escudero, R.; Alfaya, B.; Villacampa, F.; Saiz-Ladera, C.; Costa, C.; Oteo, M.; Duarte, J.; et al. In vivo disruption of an Rb-E2F-Ezh2 signaling loop causes bladder cancer. Cancer Res. 2014, 74, 6565–6577. [Google Scholar] [CrossRef] [PubMed]
- Padrao, N.A.; Monteiro-Reis, S.; Torres-Ferreira, J.; Antunes, L.; Leca, L.; Montezuma, D.; Ramalho-Carvalho, J.; Dias, P.C.; Monteiro, P.; Oliveira, J.; et al. MicroRNA promoter methylation: A new tool for accurate detection of urothelial carcinoma. Br. J. Cancer 2017, 116, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.V.; Mattick, J.S. The rise of regulatory RNA. Nat. Rev. Genet. 2014, 15, 423–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]



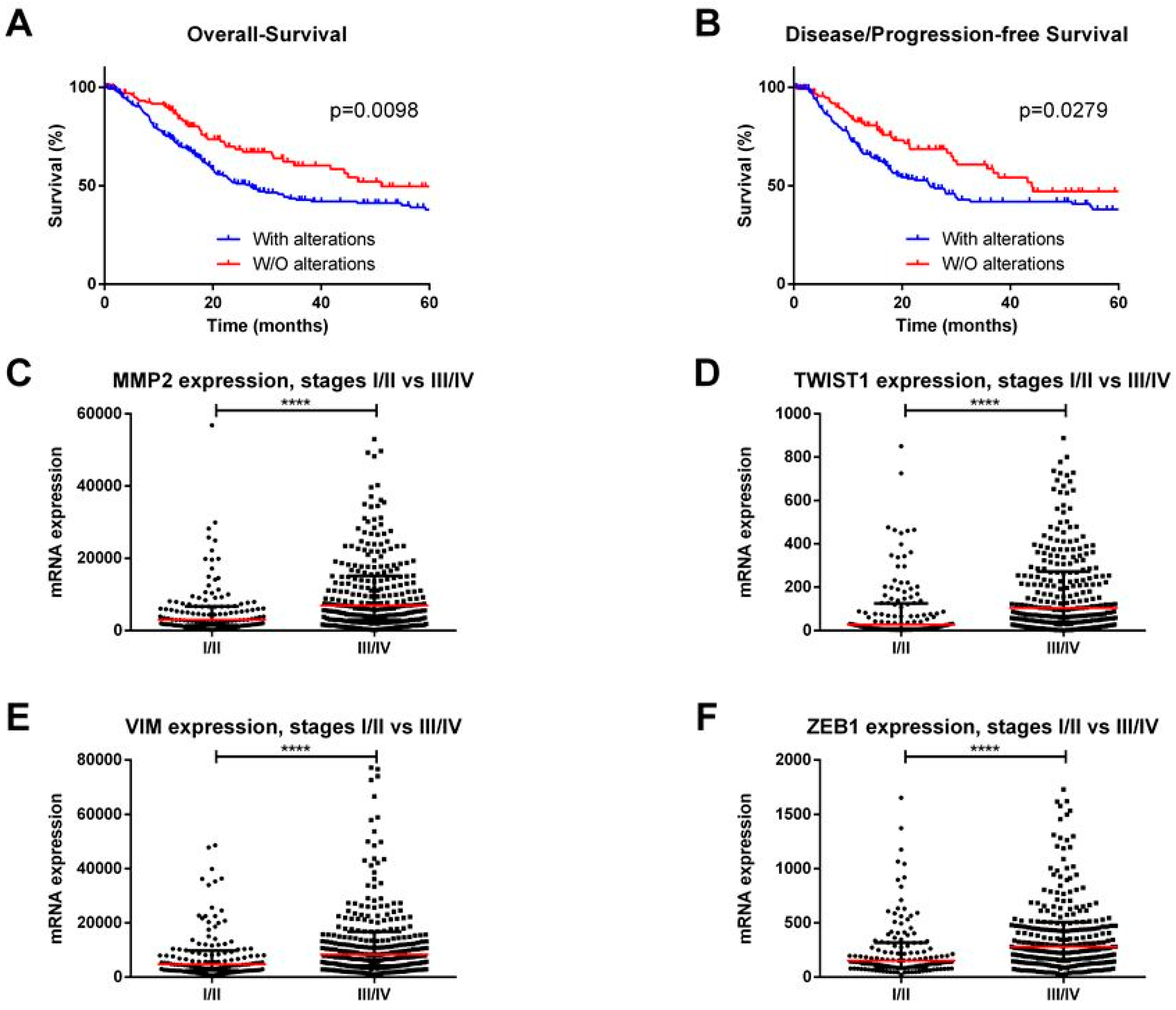{kind=link}
{kind=link}
| Gene | Expression in BlCa | Effect on EMT | Epigenetic Regulation | Sample Type and Size | Author |
|---|---|---|---|---|---|
| MAEL | Upregulated | ↑EMT (↓ECAD, ↓β-catenin, ↑Fibronectin, ↑VIM) Recruitment of DNMT3B and HDAC1/2 to MTSS1 promoter) | Downregulated by miR186 | 184 primary tumors, in vitro and in vivo assays | Li, X.D., 2016 [47] |
| GDF15 | Downregulated | ↓EMT (knockdown cells with ↓ECAD, ↑NCAD, ↑Snail, ↑Slug) | Upregulated by demethylation | In vivo assays | Tsui, K.H. and Hsu, S.Y., 2015 [48] |
| KLF4 | Downregulated | ↓EMT (↑ECAD, ↓NCAD, ↓ β-catenin, ↓VIM, ↓Snail, ↓Slug) | Promoter methylation; Upregulated by 5AZA treatment | 139 non-muscle invasive primary tumors, in vitro and in vivo assays | Li, H. and Wang, J., 2013 [49] |
| ↓EMT (Upregulation) | Promoter methylation confirmed by BSP | In vitro assays | Xu, X., 2017 [50] | ||
| PRSS8 | Downregulated | ↓EMT (↑ECAD in cells with forced PRSS8 expression) | Promoter methylation. Upregulated by 5AZA and TSA treatment | 40 primary tumors and in vivo assays | Chen, L.M., 2009 [43] |
| ELF5 | Downregulated | ↓EMT (↑ECAD, ↓NCAD, ↓VIM, ↓Snail, ↓ZEB1) | Promoter methylation. Upregulated by 5AZA treatment | 182 FFPE + 50 FF primary tumors and in vivo assays | Wu, B., 2015 [46] |
| Non-Coding RNA | Effect on EMT (and Others) | Main Regulators | Main Targets/Pathways | Sample Type and Size | Author |
|---|---|---|---|---|---|
| Small Non-Coding RNAs | |||||
| miR22 | ↓EMT, diagnostic value (↓ in tumor, vs. normal) | Snail and MAPK/Slug/VIM | 13 primary tumors, in vitro and in vivo assays | Xu, M., 2018 [90] | |
| miR23b | ↓EMT, diagnostic (↓ in tumor vs. normal) and prognostic (↑OS) value | ZEB1 | 20 primary tumors and in vivo assays | Majid, S., 2013 [68] | |
| miR24 | ↓EMT, diagnostic value (↓ in tumor, vs. normal) | CARMA3 | In vitro assays | Zhang, S., 2015 [71] | |
| miR34a | ↓EMT, diagnostic value (↓ in tumor, vs. normal) | CD44 | 8 primary tumors, in vitro and in vivo assays | Yu, G., 2014 [72] | |
| miR92 (family) | ↑EMT, diagnostic (↑ in tumor vs. normal) value, induces cisplatin resistance | GSK-3β/ Wnt/c-myc/MMP7 | 20 primary tumors and In vitro assays | Wang, H., 2016 [79] | |
| miR92b | ↑EMT | DAB2IP | In vitro assays | Huang, J., 2016 [80] | |
| miR-124-3p | ↓EMT, diagnostic value (↓ in tumor, vs. normal) | ROCK1, MMP2, MMP9 | 13 primary tumors and in vitro assays | Xu, X., 2013 [66] | |
| miR135a | ↑EMT | GSK-3β | 165 primary tumors and in vitro assays | Mao, X.W., 2018 [91] | |
| miR141 | ↓EMT, prognostic value (LN metastases) | MMP2 and 9, Vimentin, N-Cadherin, E-Cadherin | 30 primary tumors, 78 urine samples and in vitro assays | Liu, W. and Qi, L., 2015 [93] | |
| miR-148a-3p | ↓EMT, diagnostic value (↓ in tumor, vs. normal) | ↓expression mediated by DNA methylation (DNMT1) – ↑expression with 5AZA | ERBB3-AKT2-c-myc/SNAIL axis | 59 primary tumors, in vitro and in vivo assays | Wang, X., 2016 [82] |
| miR186 | ↓EMT, diagnostic value (↓ in tumor, vs. normal) | NSBP1 | 20 primary tumors and in vitro assays | Yao, K., 2015 [73] | |
| miR-199a-5p | ↓EMT, diagnostic (↓ in tumor vs. normal) and prognostic (stage, grade) value | CCR7, MMP9 | 40 primary tumors and in vitro assays | Zhou, M., 2016 [81] | |
| miR200 (family) | ↓EMT, prognostic value (↑ survival) | ↓expression mediated by EZH2 and BMI-1 | BMI-1, ZEB1, ZEB2 | 87 primary tumors and in vitro assays | Martínez-Fernández, M. and Duenas, M., 2015 [74] |
| ↓EMT and proliferation, diagnostic (↓ in tumor vs. normal) and prognostic (↑ survival) value | BMI-1 and E2F3 | 15 primary tumors and in vitro assays | Liu, L., 2014 [69] | ||
| miR200b | ↓EMT, prognostic value (LN metastases) | MMP2 and 9, Vimentin, N-Cadherin, E-Cadherin | 30 primary tumors, 78 urine samples and in vitro assays | Liu, W. and Qi, L., 2015 [93] | |
| ↓EMT | ↓expression mediated by TGF-β1 | MMP16 | In vitro assays | Chen, M.F. and Zeng, F., 2014 [75] | |
| miR200c | ↓EMT, restores sensitivity to EGFR inhibitors | ZEB1, ZEB2 and ERRFI-1 | In vitro assays | Adam, L., 2009 [65] | |
| miR203 | ↓EMT, diagnostic value (↓ in tumor, vs. normal) | Twist1 | 24 primary tumors and in vitro assays | Shen, J., 2017 [85] | |
| miR205 | ↓EMT, poor prognosis | ↑expression mediated by p63 isoform ΔNp63α | ZEB1, ZEB2 | 98 primary tumors and in vitro assays | Tran, M.N., 2013 [67] |
| miR221 | ↑EMT | ↑expression mediated by TGF-β1 | STMN1 | In vitro assays | Liu, J., 2015 [76] |
| miR224 | ↑EMT, diagnostic (↑ in tumor vs. normal) and prognostic (stage, metastases, ↓survival) value | SUFU/Hedgehog pathway | 97 primary tumors, in vitro and in vivo assays | Miao, X., Gao, H. and Liu, S., 2018 [86] | |
| miR301b | ↑EMT, diagnostic value (↑ in tumor, vs. normal) | EGR1 | In vitro assays | Yan, L., 2017 [94] | |
| miR-323a-3p | ↓EMT, diagnostic (↓ in tumor vs. normal) and prognostic (↑OS) value | ↓expression mediated by methylation of IG-DMR | Met/SMAD3/Snail | 9 primary tumors and in vivo assays | Li, J., 2017 [87] |
| miR-370-3p | ↓EMT | Wnt7a | 41 primary tumors in vitro and in vivo assays | Huang, X. and Zhu, H., 2018 [92] | |
| miR-370-5p | ↓EMT | p21 | In vitro assays | Wang, C., 2016 [95] | |
| miR424 | ↓EMT, diagnostic (↓ in tumor vs. normal) and prognostic (stage, ↑OS and DFS) value | ↓expression mediated by DNMT1 | EGFR pathway | 124 primary tumors, in vitro and in vivo assays | Wu, C.T., 2015 [77] |
| miR429 | ↓EMT | ZEB1/βcatenin axis | In vitro assays | Wu, C.L., 2016 [83] | |
| miR433 | ↓EMT, diagnostic value (↓ in tumor, vs. normal) | c-Met/CREB1-Akt/GSK-3β/Snail | 13 primary tumors and in vitro assays | Xu, X., 2016 [84] | |
| miR451 | ↓EMT, diagnostic (↓ in tumor vs. normal) and prognostic (grade and stage) value | E-Cadherin, N-Cadherin | 40 primary tumors and in vitro assays | Zeng, T. and Peng, L., 2014 [70] | |
| miR-485-5p | ↓EMT, diagnostic value (↓ in tumor vs. normal) | HMGA2 | 15 primary tumors and in vitro assays | Chen, Z., 2015 [78] | |
| miR497 | ↓EMT, diagnostic (↓ in tumor vs. normal) and prognostic (stage, metastases) value | E-Cadherin, Vimentin | 50 primary tumors and in vitro assays | Wei, Z., 2017 [88] | |
| miR612 | ↓EMT, diagnostic (↓ in tumor vs. normal) and prognostic (stage, metastases) value | ME1 | 46 primary tumors and in vitro assays | Liu, M. and Chen, Y., 2018 [96] | |
| miR613 | ↓EMT, diagnostic value (↓ in tumor vs. normal) | SphK1 | 35 primary tumors and in vitro assays | Yu, H., 2017 [89] | |
| Long non-coding RNAs | |||||
| circRNA MYLK | ↑EMT, prognostic value (stage, grade) | miR29a/VEGFA/VEGFR2 axis | 32 primary tumors, in vitro and in vivo assays | Zhong, Z., 2017 [97] | |
| lncRNA GHET1 | ↑EMT, diagnostic (↑ in tumor vs. normal) and prognostic (grade, stage, metastases, ↓OS) value | E-Cadherin, Vimentin, Fibronectin, Slug, Twist, Snail, ZEB1 | 80 primary tumors and in vitro assays | Li, L.J., 2014 [98] | |
| lncRNA HOTAIR | ↑EMT | Various EMT players | 10 primary tumors and in vitro assays | Berrondo, C., 2016 [99] | |
| lncRNA H19 | ↑EMT, diagnostic value (↑ in tumor vs. normal) | miR-29b-3p/DNMT3B axis | 35 primary tumors, in vitro and in vivo assays | Lv, M., 2017 [100] | |
| lncRNA Malat1 | ↑EMT, poor prognosis | ↑expression mediated by TGF-β | suz12 | 95 primary tumors, in vitro and in vivo assays | Fan, Y., 2014 [101] |
| lncRNA ROR | ↑EMT, diagnostic value (↑ in tumor vs. normal) | ZEB1 | 36 primary tumors and in vitro assays | Chen, Y., 2017 [102] | |
| lncRNA TP73-AS1 | ↓EMT, diagnostic (↓ in tumor vs. normal) and prognostic (↑OS and PFS) value | Various EMT players | 128 primary tumors and in vitro assays | Tuo, Z., 2018 [103] | |
| lncRNA TUG1 | ↑EMT, diagnostic (↑ in tumor vs. normal) and prognostic (stage, ↓OS) value, promotes radio-resistance | miR145/ ZEB2 axis | 54 primary tumors, in vitro and in vivo assays | Tan, J., 2015 [104] | |
| lncRNA UCA1 | ↑EMT | miR145-ZEB1/2-FSCN1 axis | In vitro assays | Xue, M., 2016 [105] | |
| miR143/HMGB1 | 52 primary tumors and in vitro assays | Luo, J., 2017 [106] | |||
| lncRNA XIST | ↑EMT | miR200c | In vitro and in vivo assays | Xu, R., 2018 [107] | |
| lncRNA ZEB2NAT | ↑EMT, diagnostic value (↑ in tumor vs. normal) | ↑expression mediated by TGF-β1 | ZEB2 | 30 primary tumors and in vitro assays | Zhuang, J. and Lu, Q., 2015 [108] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro-Reis, S.; Lobo, J.; Henrique, R.; Jerónimo, C. Epigenetic Mechanisms Influencing Epithelial to Mesenchymal Transition in Bladder Cancer. Int. J. Mol. Sci. 2019, 20, 297. https://doi.org/10.3390/ijms20020297
Monteiro-Reis S, Lobo J, Henrique R, Jerónimo C. Epigenetic Mechanisms Influencing Epithelial to Mesenchymal Transition in Bladder Cancer. International Journal of Molecular Sciences. 2019; 20(2):297. https://doi.org/10.3390/ijms20020297
Chicago/Turabian StyleMonteiro-Reis, Sara, João Lobo, Rui Henrique, and Carmen Jerónimo. 2019. "Epigenetic Mechanisms Influencing Epithelial to Mesenchymal Transition in Bladder Cancer" International Journal of Molecular Sciences 20, no. 2: 297. https://doi.org/10.3390/ijms20020297
APA StyleMonteiro-Reis, S., Lobo, J., Henrique, R., & Jerónimo, C. (2019). Epigenetic Mechanisms Influencing Epithelial to Mesenchymal Transition in Bladder Cancer. International Journal of Molecular Sciences, 20(2), 297. https://doi.org/10.3390/ijms20020297