Characterization of QTLs for Seedling Resistance to Tan Spot and Septoria Nodorum Blotch in the PBW343/Kenya Nyangumi Wheat Recombinant Inbred Lines Population


 , , and
, , and
Abstract
:1. Introduction
2. Results
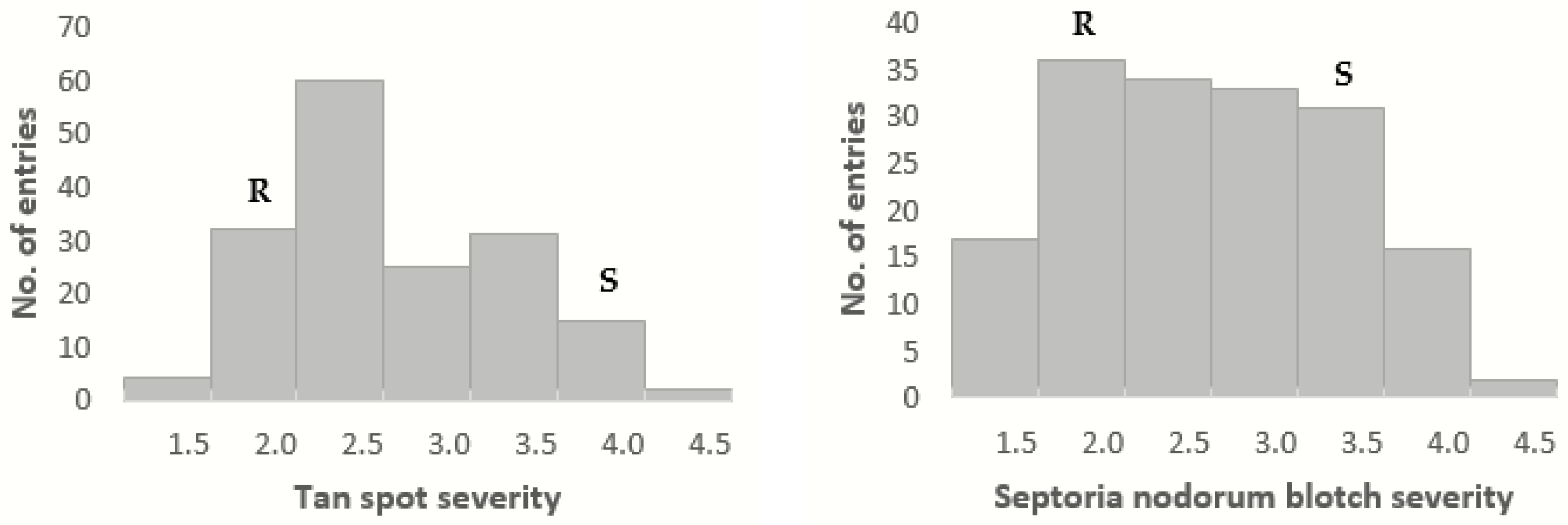
2.1. Disease Phenotyping
2.2. Molecular Analysis
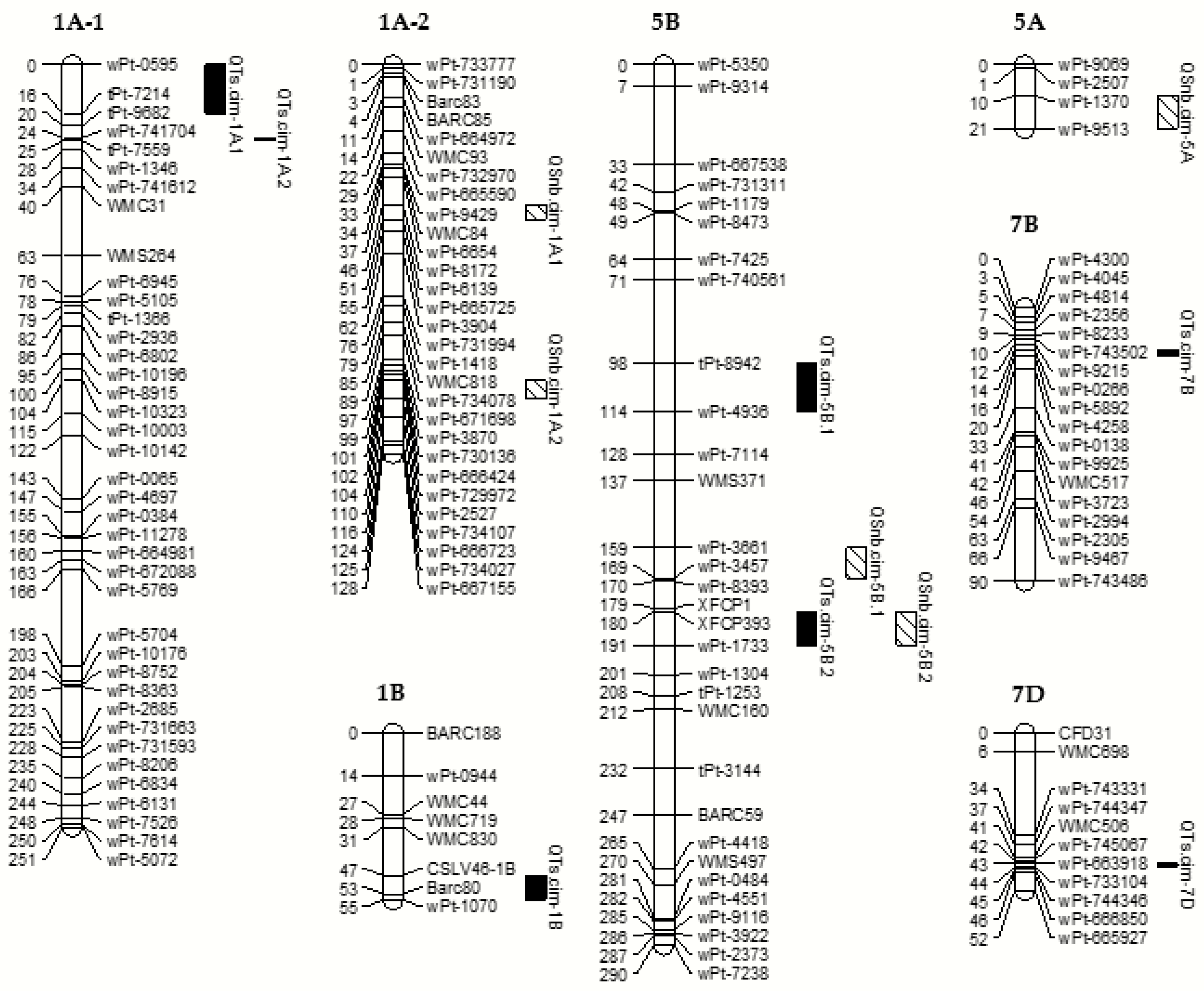
2.3. QTL Analysis
2.4. Interaction among Important QTLs
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Design
4.3. Disease Screening
4.4. Molecular Marker Analysis
4.5. Linkage Mapping and QTL Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Rees, R.; Platz, G.; Mayer, R. Susceptibility of Australian wheats to Pyrenophora tritici-repentis. Crop Pasture Sci. 1988, 39, 141–151. [Google Scholar] [CrossRef]
- Shabeer, A.; Bockus, W. Tan spot effects on yield and yield components relative to growth stage in winter wheat. Plant Dis. 1988, 72, 599–602. [Google Scholar] [CrossRef]
- Murray, G.; Brennan, J. Estimating disease losses to the Australian wheat industry. Australas. Plant Pathol. 2009, 38, 558–570. [Google Scholar] [CrossRef]
- De Wolf, E.D.; Effertz, R.J.; Ali, S.; Francl, L.J. Vistas of tan spot research. Can. J. Plant Pathol. 1998, 20, 349–370. [Google Scholar] [CrossRef]
- Fernandez, M.R.; DePauw, R.M.; Clarke, J.M.; Fox, S.L. Discoloration of wheat kernels by Pyrenophora tritici-repentis. Can. J. Plant Pathol. 1998, 20, 380–383. [Google Scholar] [CrossRef]
- Singh, P.K.; Singh, R.P.; Duveiller, E.; Mergoum, M.; Adhikari, T.B.; Elias, E.M. Genetics of wheat–Pyrenophora tritici-repentis interactions. Euphytica 2010, 171, 1–13. [Google Scholar] [CrossRef]
- Andrie, R.M.; Pandelova, I.; Ciuffetti, L.M. A combination of phenotypic and genotypic characterization strengthens Pyrenophora tritici-repentis race identification. Phytopathology 2007, 97, 694–701. [Google Scholar] [CrossRef]
- Ruud, A.K.; Windju, S.; Belova, T.; Friesen, T.L.; Lillemo, M. Mapping of SnTox3–Snn3 as a major determinant of field susceptibility to Septoria nodorum leaf blotch in the SHA3/CBRD × Naxos population. Theor. Appl. Genet. 2017, 130, 1361–1374. [Google Scholar] [CrossRef]
- Faris, J.D.; Anderson, J.A.; Francl, L.J.; Jordahl, J.G. Chromosomal location of a gene conditioning insensitivity in wheat to a necrosis-inducing culture filtrate from Pyrenophora tritici-repentis. Phytopathology 1996, 86, 459–463. [Google Scholar] [CrossRef]
- Anderson, J.A.; Effertz, R.J.; Faris, J.D.; Franel, L.J.; Meinhardt, S.W.; Gill, B.S. Genetic analysis of sensitivity to a Pyrenophora tritici-repentis necrosis-inducing toxin in durum and common wheat. Phytopathology 1999, 89, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Hughes, G.R. Inheritance of resistance to the chlorosis component of tan spot of wheat caused by Pyrenophora tritici-repentis, races 1 and 3. Euphytica 2006, 152, 413–420. [Google Scholar] [CrossRef]
- Tadesse, W.; Schmolke, M.; Hsam, S.L.K.; Mohler, V.; Wenzel, G.; Zeller, F.J. Molecular mapping of resistance genes to tan spot [Pyrenophora tritici-repentis race 1] in synthetic wheat lines. Theor. Appl. Genet. 2007, 114, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Mergoum, M.; Ali, S.; Adhikari, T.B.; Hughes, G.R. Genetic analysis of resistance to Pyrenophora tritici-repentis races 1 and 5 in tetraploid and hexaploid wheat. Phytopathology 2008, 98, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.; Wallwork, H.; Williams, K.J. Identification of a major QTL for yellow leaf spot resistance in the wheat varieties Brookton and Cranbrook. Aust. J. Agric. Res. 2004, 55, 315–319. [Google Scholar] [CrossRef]
- Friesen, T.L.; Faris, J.D. Molecular mapping of resistance to Pyrenophora tritici-repentis race 5 and sensitivity to Ptr ToxB in wheat. Theor. Appl. Genet. 2004, 109, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Faris, J.D.; Friesen, T.L. Identification of quantitative trait loci for race-nonspecific resistance to tan spot in wheat. Theor. Appl. Genet. 2005, 111, 386–392. [Google Scholar] [CrossRef]
- Singh, S.; Bockus, W.W.; Sharma, I.; Bowden, R.L. A novel source of resistance in wheat to Pyrenophora tritici-repentis race 1. Plant Dis. 2008, 92, 91–95. [Google Scholar] [CrossRef]
- Juliana, P.; Singh, R.P.; Singh, P.K.; Poland, J.A.; Bergstrom, G.C.; Huerta-Espino, J.; Bhavani, S.; Crossa, J.; Sorrells, M.E. Genome-wide association mapping for resistance to leaf rust, stripe rust and tan spot in wheat reveals potential candidate genes. Theor. Appl. Genet. 2018. [Google Scholar] [CrossRef]
- Hu, W.; He, X.; Dreisigacker, S.; Sansaloni, C.P.; Juliana, P.; Singh, P.K. A wheat chromosome 5AL region confers seedling resistance to both tan spot and Septoria nodorum blotch in two mapping populations. Crop J. 2019, in press. [Google Scholar] [CrossRef]
- Effertz, R.J.; Anderson, J.A.; Francl, L.J. Restriction fragment length polymorphism mapping of resistance to two races of Pyrenophora tritici-repentis in adult and seedling wheat. Phytopathology 2001, 91, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Ciuffetti, L.M.; Tuori, R.P.; Gaventa, J.M. A single gene encodes a selective toxin causal to the development of tan spot of wheat. Plant Cell 1997, 9, 135–144. [Google Scholar] [PubMed]
- Orolaza, N.P.; Lamari, L.; Ballance, G.M. Evidence of a host-specific chlorosis toxin from Pyrenophora tritici-repentis, the causal agent of tan spot of wheat. Phytopathology 1995, 85, 1282–1287. [Google Scholar] [CrossRef]
- Effertz, R.J.; Meinhardt, S.W.; Anderson, J.A.; Jordahl, J.G.; Francl, L.J. Identification of a chlorosis-inducing toxin from Pyrenophora tritici-repentis and the chromosomal location of an insensitivity locus in wheat. Phytopathology 2002, 92, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Mergoum, M.; Adhikari, T.B.; Shah, T.; Ghavami, F.; Kianian, S.F. Genetic and molecular analysis of wheat tan spot resistance effective against Pyrenophora tritici-repentis races 2 and 5. Mol. Breed. 2010, 25, 369–379. [Google Scholar] [CrossRef]
- Friesen, T.L.; Ali, S.; Kianian, S.; Francl, L.J.; Rasmussen, J.B. Role of host sensitivity to Ptr ToxA in development of tan spot of wheat. Phytopathology 2003, 93, 397–401. [Google Scholar] [CrossRef]
- Faris, J.D.; Anderson, J.A.; Francl, L.J.; Jordahl, J.G. RFLP mapping of resistance to chlorosis induction by Pyrenophora tritici-repentis in wheat. Theor. Appl. Genet. 1997, 94, 98–103. [Google Scholar] [CrossRef]
- Ma, H.; Hughes, G.R. Genetic control and chromosomal location of Triticum timopheevii-derived resistance to septoria nodorum blotch in durum wheat. Genome 1995, 38, 332–338. [Google Scholar] [CrossRef]
- Murphy, N.E.A.; Loughman, R.; Wilson, R.; Lagudah, E.S.; Appels, R.; Jones, M.G.K. Resistance to septoria nodorum blotch in the Aegilops tauschii accession RL5271 is controlled by a single gene. Euphytica 2000, 113, 227–233. [Google Scholar] [CrossRef]
- Nelson, L.; Gates, C. Genetics of host plant resistance of wheat to Septoria nodorum. Crop Sci. 1982, 22, 771–773. [Google Scholar] [CrossRef]
- Fried, P.; Meister, E. Inheritance of leaf and head resistance of winter wheat to Septoria nodorum in a diallel cross. Phytopathology 1987, 77, 1371–1375. [Google Scholar] [CrossRef]
- Bostwick, D.E.; Ohm, H.W.; Shaner, G. Inheritance of Septoria glume blotch resistance in wheat. Crop Sci. 1993, 33, 439–443. [Google Scholar] [CrossRef]
- Wicki, W.; Winzeler, M.; Schmid, J.E.; Stamp, P.; Messmer, M. Inheritance of resistance to leaf and glume blotch caused by Septoria nodorum Berk. in winter wheat. Theor. Appl. Genet. 1999, 99, 1265–1272. [Google Scholar] [CrossRef]
- Ruud, A.K.; Dieseth, J.A.; Lillemo, M. Effects of three necrotrophic effectors on spring wheat under Norwegian field conditions. Crop Sci. 2017. [Google Scholar] [CrossRef]
- Francki, M.G. Improving Stagonospora nodorum resistance in wheat: A review. Crop Sci. 2013, 53, 355–365. [Google Scholar] [CrossRef]
- Friesen, T.L.; Chu, C.; Xu, S.S.; Faris, J.D. SnTox5-Snn5: A novel Stagonospora nodorum effector-wheat gene interaction and its relationship with the SnToxA-Tsn1 and SnTox3-Snn3-B1 interactions. Mol. Plant Pathol. 2012, 13, 1101–1109. [Google Scholar] [CrossRef]
- Shi, G.; Friesen, T.L.; Saini, J.; Xu, S.S.; Rasmussen, J.B.; Faris, J.D. The wheat snn7 gene confers susceptibility on recognition of the Parastagonospora nodorum necrotrophic effector SnTox7. Plant Genome 2015, 8. [Google Scholar] [CrossRef]
- Liu, Z.H.; Friesen, T.L.; Rasmussen, J.B.; Ali, S.; Meinhardt, S.W.; Faris, J.D. Quantitative trait loci analysis and mapping of seedling resistance to Stagonospora nodorum leaf blotch in wheat. Phytopathology 2004, 94, 1061–1067. [Google Scholar] [CrossRef]
- Singh, P.K.; Mergoum, M.; Adhikari, T.B.; Kianian, S.F.; Elias, E.M. Chromosomal location of genes for seedling resistance to tan spot and Stagonospora nodorum blotch in tetraploid wheat. Euphytica 2007, 155, 27–34. [Google Scholar] [CrossRef]
- Shankar, M.; Walker, E.; Golzar, H.; Loughman, R.; Wilson, R.E.; Francki, M.G. Quantitative trait loci for seedling and adult plant resistance to Stagonospora nodorum in wheat. Phytopathology 2008, 98, 886–893. [Google Scholar] [CrossRef]
- Adhikari, T.B.; Jackson, E.W.; Gurung, S.; Hansen, J.M.; Bonman, J.M. Association mapping of quantitative resistance to Phaeosphaeria nodorum in spring wheat landraces from the USDA National Small Grains Collection. Phytopathology 2011, 101, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.T.T.; Rybak, K.; Bertazzoni, S.; Furuki, E.; Dinglasan, E.; Hickey, L.T.; Oliver, R.P.; Tan, K.C. Novel sources of resistance to Septoria nodorum blotch in the Vavilov wheat collection identified by genome-wide association studies. Theor. Appl. Genet. 2018, 131, 1223–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parlevliet, J.E. Durability of resistance against fungal, bacterial and viral pathogens; present situation. Euphytica 2002, 124, 147–156. [Google Scholar] [CrossRef]
- Lillemo, M.; Joshi, A.K.; Prasad, R.; Chand, R.; Singh, R.P. QTL for spot blotch resistance in bread wheat line Saar co-locate to the biotrophic disease resistance loci Lr34 and Lr46. Theor. Appl. Genet. 2013, 126, 711–719. [Google Scholar] [CrossRef]
- Singh, P.K.; Duveiller, E.; Singh, R.P. Resistance breeding for tan spot (Pyrenophora tritici-repentis) of wheat. In Disease Resistance in Wheat; Sharma, I., Ed.; CABI: Wallingford, UK, 2012; pp. 136–150. [Google Scholar]
- Faris, J.D.; Liu, Z.; Xu, S.S. Genetics of tan spot resistance in wheat. Theor. Appl. Genet. 2013, 126, 2197–2217. [Google Scholar] [CrossRef]
- Singh, P.K.; Crossa, J.; Duveiller, E.; Singh, R.P.; Djurle, A. Association mapping for resistance to tan spot induced by Pyrenophora tritici-repentis race 1 in CIMMYTs historical bread wheat set. Euphytica 2016, 207, 515–525. [Google Scholar] [CrossRef]
- Singh, S.; Singh, R.P.; Bhavani, S.; Huerta-Espino, J.; Eugenio, L.-V.E. QTL mapping of slow-rusting, adult plant resistance to race Ug99 of stem rust fungus in PBW343/Muu RIL population. Theor. Appl. Genet. 2013, 126, 1367–1375. [Google Scholar] [CrossRef]
- Chu, C.G.; Friesen, T.L.; Xu, S.S.; Faris, J.D. Identification of novel tan spot resistance loci beyond the known host-selective toxin insensitivity genes in wheat. Theor. Appl. Genet. 2008, 117, 873–881. [Google Scholar] [CrossRef]
- Lu, H.J.; Fellers, J.P.; Friesen, T.L.; Meinhardt, S.W.; Faris, J.D. Genomic analysis and marker development for the Tsn1 locus in wheat using bin-mapped ESTs and flanking BAC contigs. Theor. Appl. Genet. 2006, 112, 1132–1142. [Google Scholar] [CrossRef]
- Sun, X.C.; Bockus, W.; Bai, G. Quantitative trait loci for resistance to Pyrenophora tritici-repentis race 1 in a Chinese wheat. Phytopathology 2010, 100, 468–473. [Google Scholar] [CrossRef]
- Faris, J.D.; Abeysekara, N.S.; McClean, P.E.; Xu, S.S.; Friesen, T.L. Tan spot susceptibility governed by the Tsn1 locus and race-nonspecific resistance quantitative trait loci in a population derived from the wheat lines Salamouni and Katepwa. Mol. Breed. 2012, 30, 1669–1678. [Google Scholar] [CrossRef]
- Lagudah, E.S.; Krattinger, S.G.; Herrera-Foessel, S.; Singh, R.P.; Huerta-Espino, J.; Spielmeyer, W.; Brown-Guedira, G.; Selter, L.L.; Keller, B. Gene-specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor. Appl. Genet. 2009, 119, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J.; Burgueno, J.; Dreisigacker, S.; Vargas, M.; Herrera-Foessel, S.A.; Lillemo, M.; Singh, R.P.; Trethowan, R.; Warburton, M.; Franco, J.; et al. Association analysis of historical bread wheat germplasm using additive genetic covariance of relatives and population structure. Genetics 2007, 177, 1889–1913. [Google Scholar] [CrossRef] [PubMed]
- Lamari, L.; Bernier, C.C. Evaluation of wheat lines and cultivars to tan spot (Pyrenophora tritici-repentis) based on lesion type. Can. J. Plant Pathol. 1989, 11, 49–56. [Google Scholar] [CrossRef]
- Singh, S.; Bowden, R.L. Molecular mapping of adult-plant race-specific leaf rust resistance gene Lr12 in bread wheat. Mol. Breed. 2011, 28, 137–142. [Google Scholar] [CrossRef]
- Wenzl, P.; Li, H.; Carling, J.; Zhou, M.; Raman, H.; Paul, E.; Hearnden, P.; Maier, C.; Xia, L.; Caig, V.; et al. A high-density consensus map of barley linking DArT markers to SSR, RFLP and STS loci and agricultural traits. BMC Genom. 2006, 7, 206. [Google Scholar] [CrossRef]
- Neumann, K.; Kobiljski, B.; Denčić, S.; Varshney, R.; Börner, A. Genome-wide association mapping: A case study in bread wheat (Triticum aestivum L.). Mol. Breed. 2011, 27, 37–58. [Google Scholar] [CrossRef]
- Li, H.; Ribaut, J.-M.; Li, Z.; Wang, J. Inclusive composite interval mapping (ICIM) for digenic epistasis of quantitative traits in biparental populations. Theor. Appl. Genet. 2008, 116, 243–260. [Google Scholar] [CrossRef]
- Smith, A.; Cullis, B.R.; Thompson, R. The analysis of crop cultivar breeding and evaluation trials: An overview of current mixed model approaches. J. Agric. Sci. 2005, 143, 449–462. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Experiment | Source | Degree of Freedom | F Statistic | p Values |
|---|---|---|---|---|
| TS | Check | 3 | 107.14 | <0.001 |
| Exp | 1 | 0.53 | 0.634 | |
| SNB | Check | 4 | 73.59 | <0.001 |
| Exp | 2 | 0.52 | 0.642 |
| Experiment | Source | Variance Component | % Total Variance | Standard Error |
|---|---|---|---|---|
| TS | RILs | 0.390 | 68.720 | 0.042 |
| Exp.Rep | 0.001 | 0.123 | 0.001 | |
| RIL.Exp.Rep | 0.056 | 9.857 | 0.030 | |
| Residuals | 0.121 | 21.299 | 0.029 | |
| Total | 0.568 | 100 | ||
| SNB | RILs | 0.4737 | 63.618 | 0.0514 |
| Exp.Rep | 0.0035 | 0.47 | 0.0039 | |
| RIL.Exp.Rep | 0.1344 | 18.05 | 0.0335 | |
| Residuals | 0.133 | 17.862 | 0.0314 | |
| Total | 0.7446 | 100 |
| QTL | Chr. | Pos. a | Flanking Markers | LOD | PVE (%) b | Add c | |
|---|---|---|---|---|---|---|---|
| Left | Right | ||||||
| QTs.cim-5B.1 | 5B | 112 | wPt-8942 | wPt-4936 | 4.33 | 2.98 | −0.11 |
| QTs.cim-5B.2 | 5B | 180 | XFCP393 | wPt-1733 | 17.73 | 23.32 | −0.213 |
| QTs.cim-7B | 7B | 14 | wPt-0266 | wPt-5892 | 35.27 | 11.75 | 0.23 |
| QTs.cim-1B | 1B | 52.8 | CSLV46-1B | wPt-1070 | 4.83 | 3.00 | 0.11 |
| QTs.cim-7D | 7D | 43 | wPt-663918 | wPt-733104 | 8.86 | 5.78 | −0.15 |
| QTs.cim-1A.1 | 1A | 0 | wPt-0595 | tPt-7214 | 6.02 | 3.69 | −0.12 |
| QTs.cim-1A.2 | 1A | 25 | wPt-741704 | tPt-7559 | 23.18 | 19.00 | −0.29 |
| QTL | Chr. | Pos. a | Flanking Markers | LOD | PVE (%) b | Add c | |
|---|---|---|---|---|---|---|---|
| Left | Right | ||||||
| QSnb.cim-5B.1 | 5B | 168 | wPt-3661 | wPt-3457 | 4.88 | 8.00 | −0.20 |
| QSnb.cim-5B.2 | 5B | 180 | XFCP393 | wPt-1733 | 11.38 | 19.51 | −0.32 |
| QSnb.cim-1A.1 | 1A | 47 | wPt-8172 | wPt-6139 | 5.70 | 5.24 | 0.17 |
| QSnb.cim-1A.2 | 1A | 104 | wPt-729972 | wPt-2527 | 12.05 | 11.45 | −0.26 |
| QSnb.cim-5A | 5A | 10 | wPt-1370 | wPt-9513 | 11.24 | 10.69 | −0.25 |
| QTL | Absence | Presence | Difference | p Value | R2 |
|---|---|---|---|---|---|
| QTs.cim-5B.2 | 2.94 | 2.15 | 0.79 | <0.0001 | 0.31 |
| QTs.cim-7B | 2.75 | 2.49 | 0.26 | 0.0127 | 0.04 |
| QTs.cim-1B | 2.83 | 2.49 | 0.34 | 0.0045 | 0.06 |
| QTs.cim-1A.2 | 2.85 | 2.23 | 0.62 | <0.0001 | 0.20 |
| QTs.cim-5B.2 + QTs.cim-7B | 2.80 | 2.23 | 0.57 | <0.0001 | 0.33 |
| QTs.cim-7B + QTs.cim-1B | 3.14 | 2.44 | 0.70 | 0.0002 | 0.15 |
| QTs.cim-5B.2 + QTs.cim-1B | 2.80 | 2.26 | 0.55 | <0.0001 | 0.36 |
| QTs.cim-1B + QTs.cim-1A.2 | 2.81 | 2.57 | 0.25 | <0.0001 | 0.30 |
| QTs.cim-7B + QTs.cim-1A.2 | 2.70 | 2.27 | 0.43 | <0.0001 | 0.22 |
| QTs.cim-5B.2 + QTs.cim-1A.2 | 3.26 | 1.63 | 1.63 | <0.0001 | 0.59 |
| QTs.cim-5B.2 + QTs.cim-7B + QTs.cim-1B | 2.85 | 2.48 | 0.36 | <0.0001 | 0.41 |
| QTs.cim-5B.2 + QTs.cim-7B + QTs.cim-1A.2 | 3.18 | 1.58 | 1.60 | <0.0001 | 0.59 |
| QTs.cim-7B + QTs.cim-1B + QTs.cim-1A.2 | 2.95 | 2.71 | 0.24 | <0.0001 | 0.37 |
| QTs.cim-5B.2 + QTs.cim-7B + QTs.cim-1B + QTs.cim-1A.2 | 3.07 | 2.67 | 0.40 | <0.0001 | 0.66 |
| QTL | Absence | Presence | Difference | p Value | R2 |
|---|---|---|---|---|---|
| QSnb.cim-5B.2 | 3.00 | 2.01 | 1.00 | <0.0001 | 0.43 |
| QSnb.cim-1A.2 | 2.53 | 2.60 | 0.07 | 0.5562 | 0.00 |
| QSnb.cim-5A | 2.62 | 2.40 | 0.22 | 0.0630 | 0.02 |
| QSnb.cim-5B.2 + QSnb.cim-1A.2 | 2.98 | 1.99 | 0.99 | <0.0001 | 0.40 |
| QSnb.cim-5B.2 + QSnb.cim-5A | 3.15 | 1.86 | 1.28 | <0.0001 | 0.44 |
| QSnb.cim-1A.2 + QSnb.cim-5A | 2.58 | 2.25 | 0.33 | 0.1222 | 0.04 |
| QSnb.cim-5B.2 + QSnb.cim-1A.2 + QSnb.cim-5A | 3.11 | 1.70 | 1.41 | <0.0001 | 0.49 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.K.; Singh, S.; Deng, Z.; He, X.; Kehel, Z.; Singh, R.P. Characterization of QTLs for Seedling Resistance to Tan Spot and Septoria Nodorum Blotch in the PBW343/Kenya Nyangumi Wheat Recombinant Inbred Lines Population. Int. J. Mol. Sci. 2019, 20, 5432. https://doi.org/10.3390/ijms20215432
Singh PK, Singh S, Deng Z, He X, Kehel Z, Singh RP. Characterization of QTLs for Seedling Resistance to Tan Spot and Septoria Nodorum Blotch in the PBW343/Kenya Nyangumi Wheat Recombinant Inbred Lines Population. International Journal of Molecular Sciences. 2019; 20(21):5432. https://doi.org/10.3390/ijms20215432
Chicago/Turabian StyleSingh, Pawan Kumar, Sukhwinder Singh, Zhiying Deng, Xinyao He, Zakaria Kehel, and Ravi Prakash Singh. 2019. "Characterization of QTLs for Seedling Resistance to Tan Spot and Septoria Nodorum Blotch in the PBW343/Kenya Nyangumi Wheat Recombinant Inbred Lines Population" International Journal of Molecular Sciences 20, no. 21: 5432. https://doi.org/10.3390/ijms20215432
APA StyleSingh, P. K., Singh, S., Deng, Z., He, X., Kehel, Z., & Singh, R. P. (2019). Characterization of QTLs for Seedling Resistance to Tan Spot and Septoria Nodorum Blotch in the PBW343/Kenya Nyangumi Wheat Recombinant Inbred Lines Population. International Journal of Molecular Sciences, 20(21), 5432. https://doi.org/10.3390/ijms20215432