Current Understanding of Residual Force Enhancement: Cross-Bridge Component and Non-Cross-Bridge Component



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
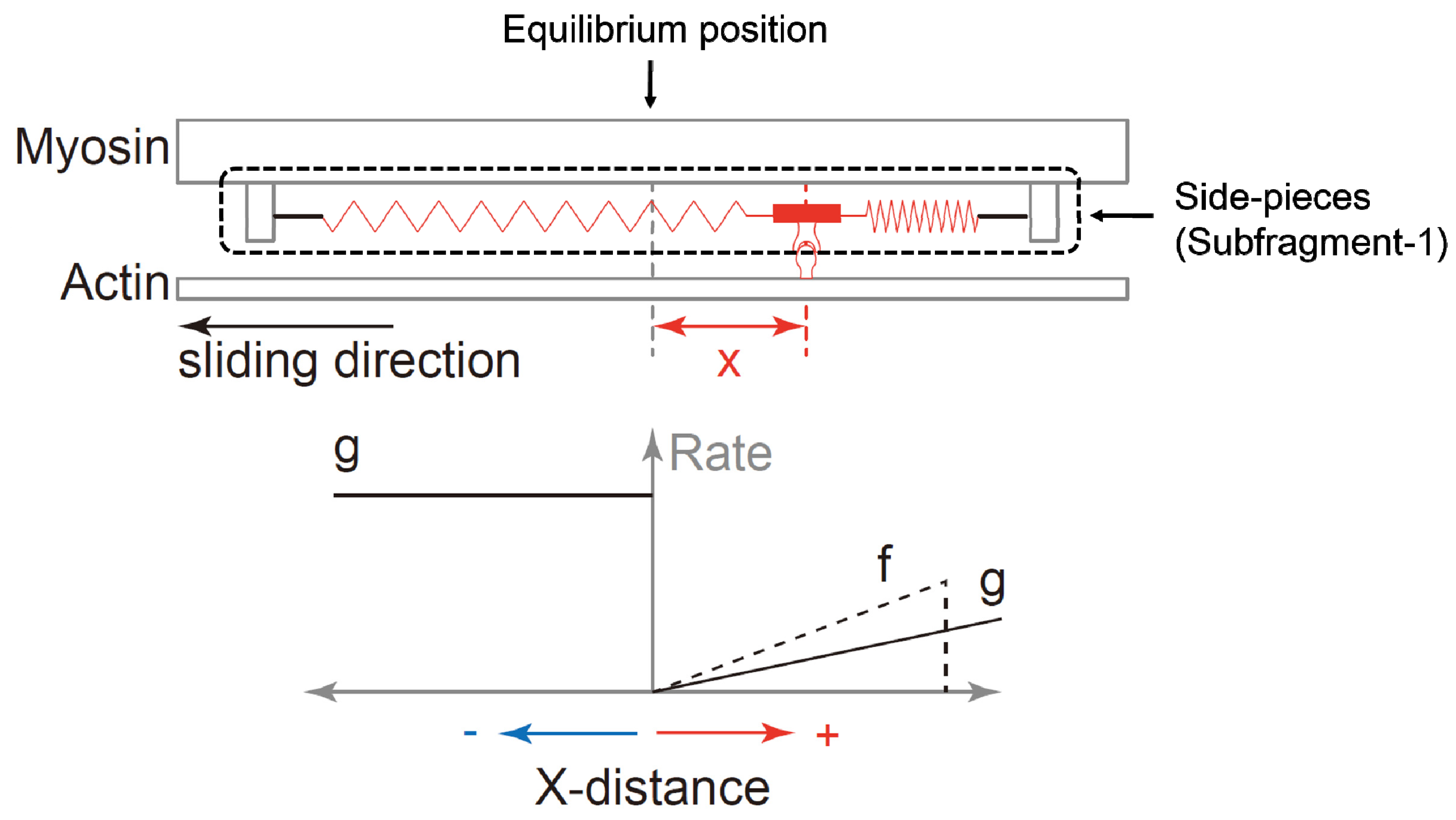
2. Cross-Bridge Theory
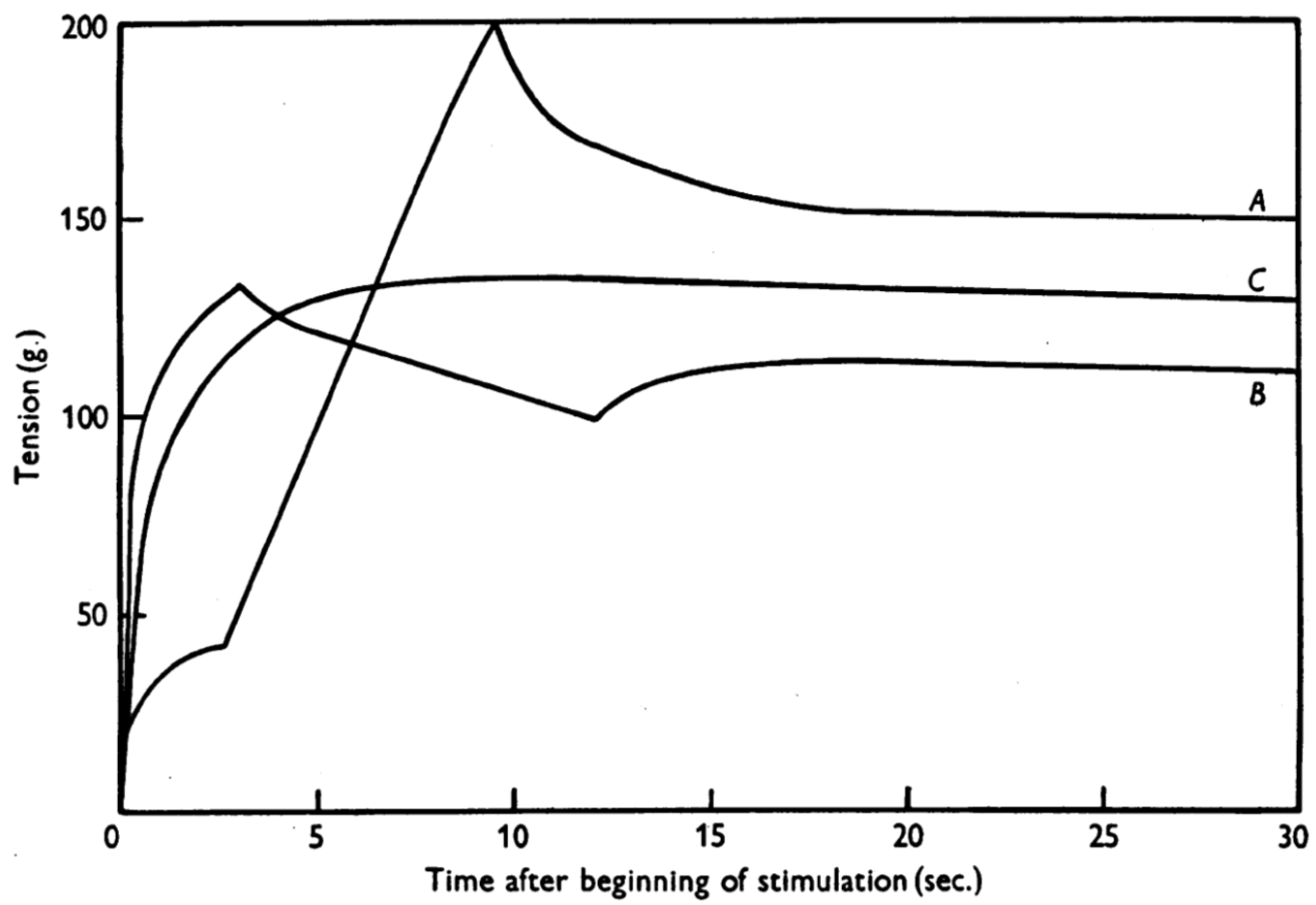
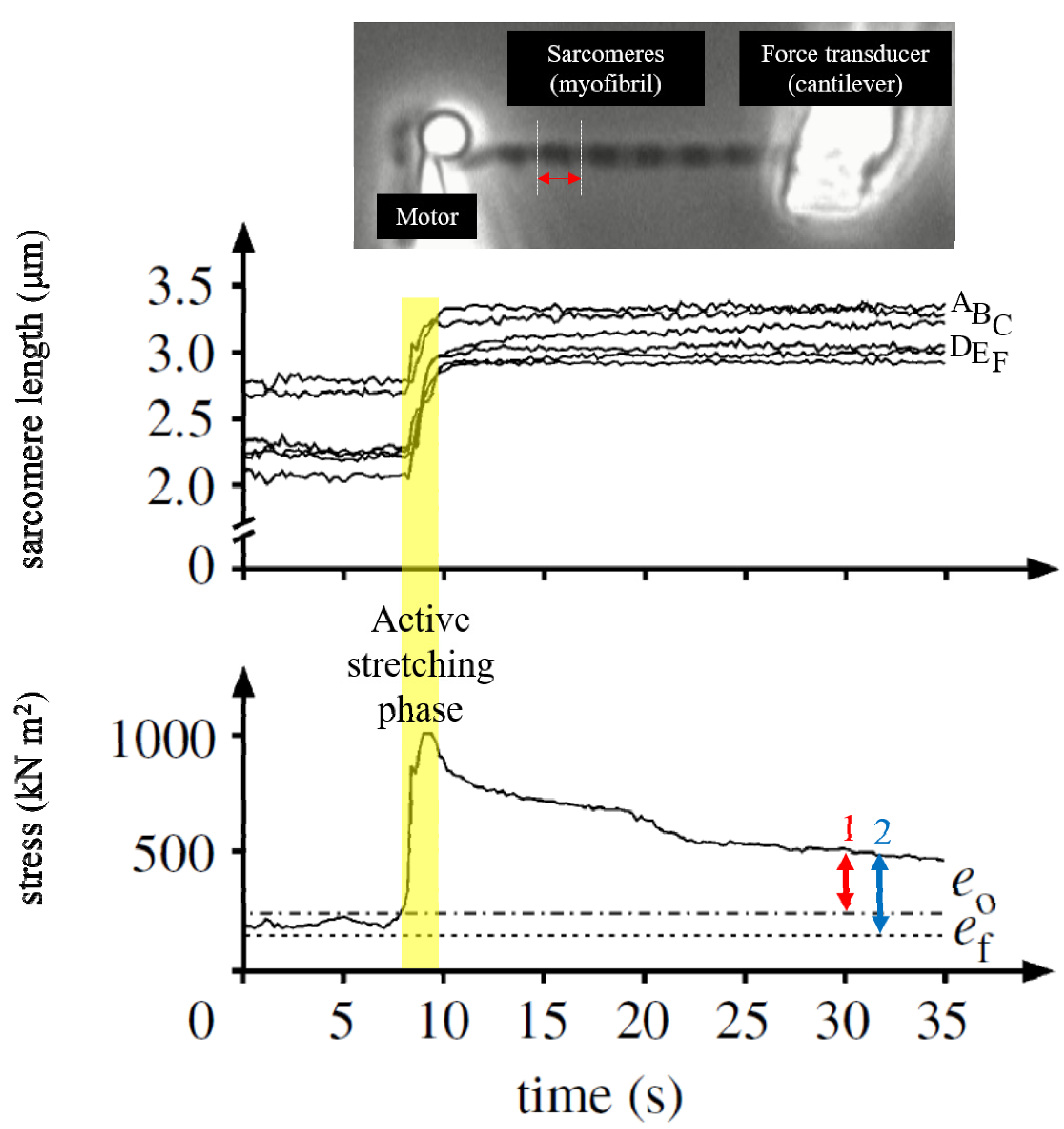
3. Residual Force Enhancement (rFE)
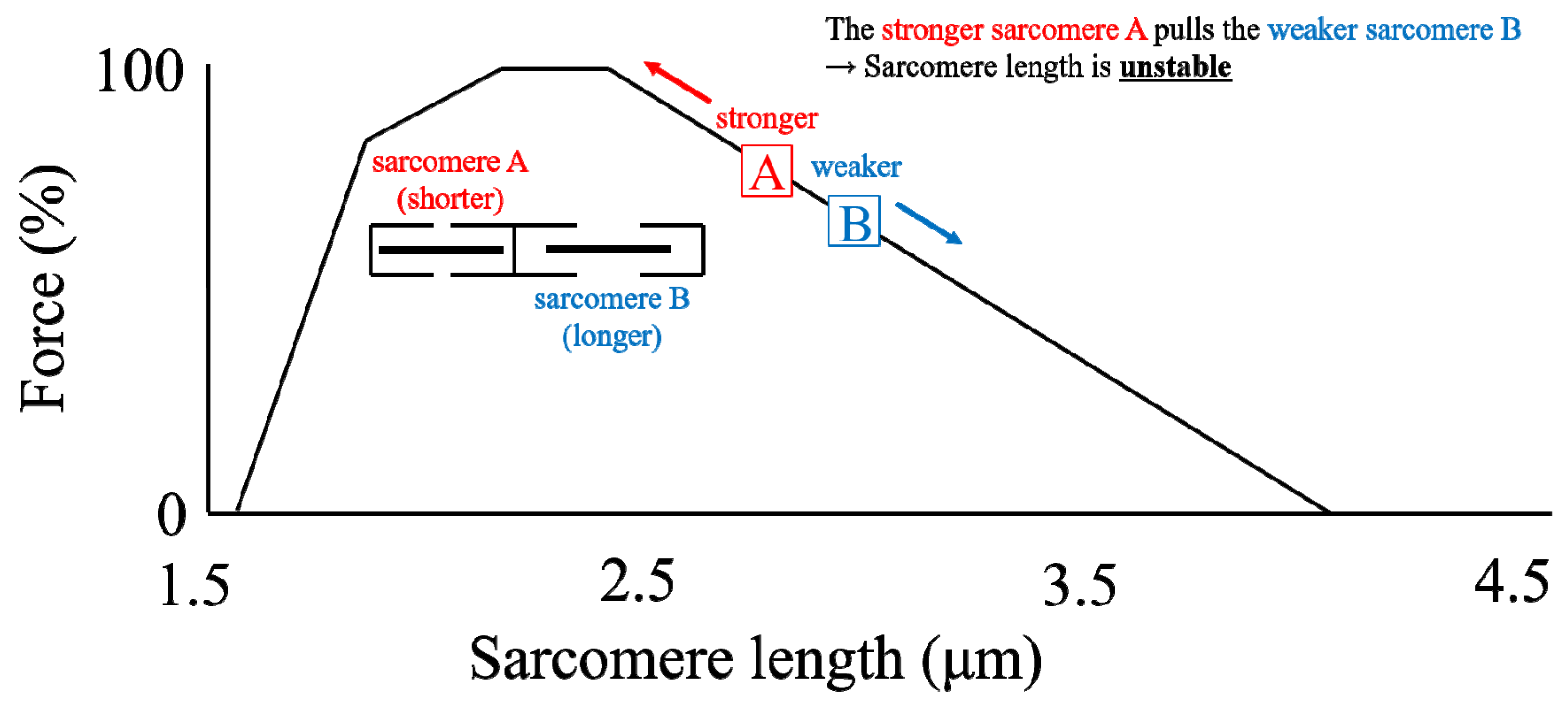
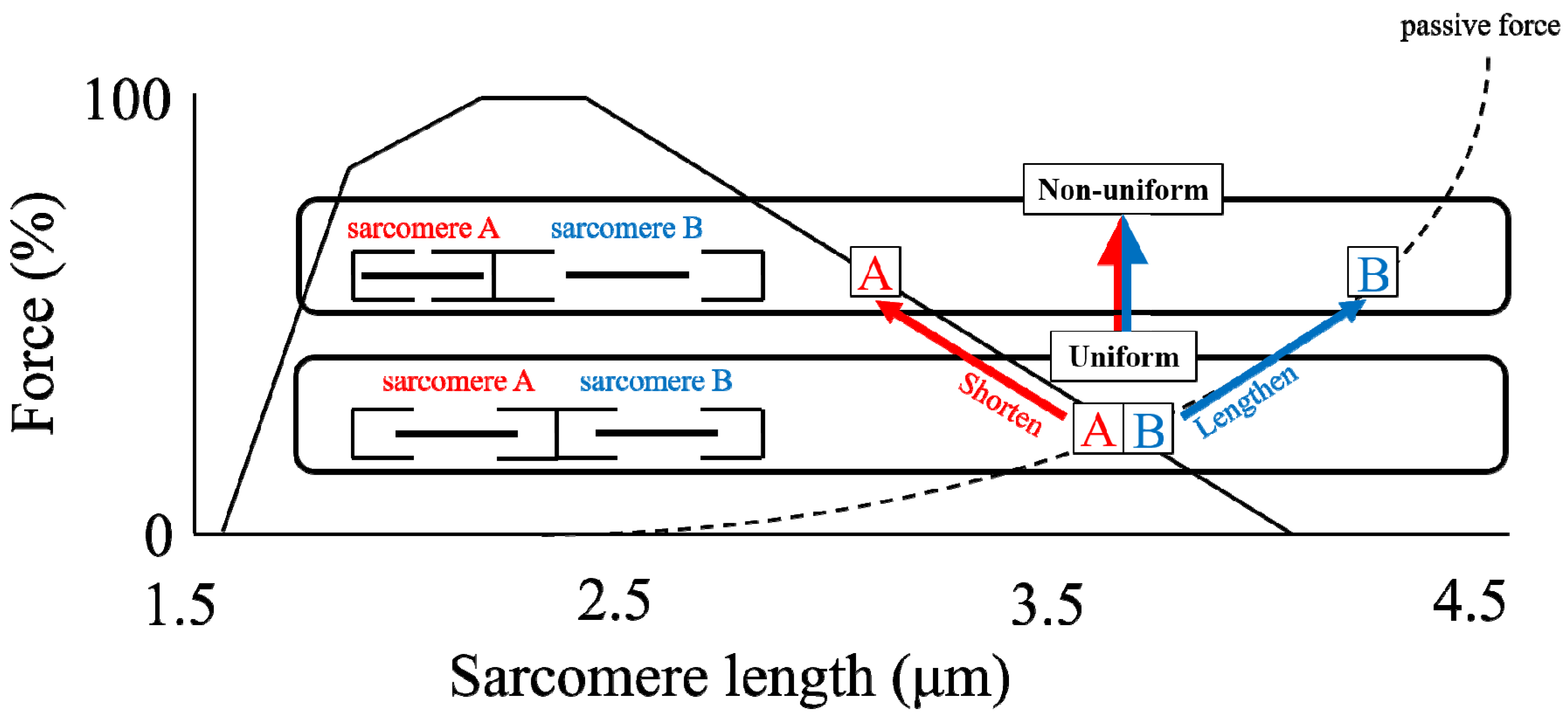
4. Sarcomere Length Non-Uniformity
4.1. Concept
4.2. Problems
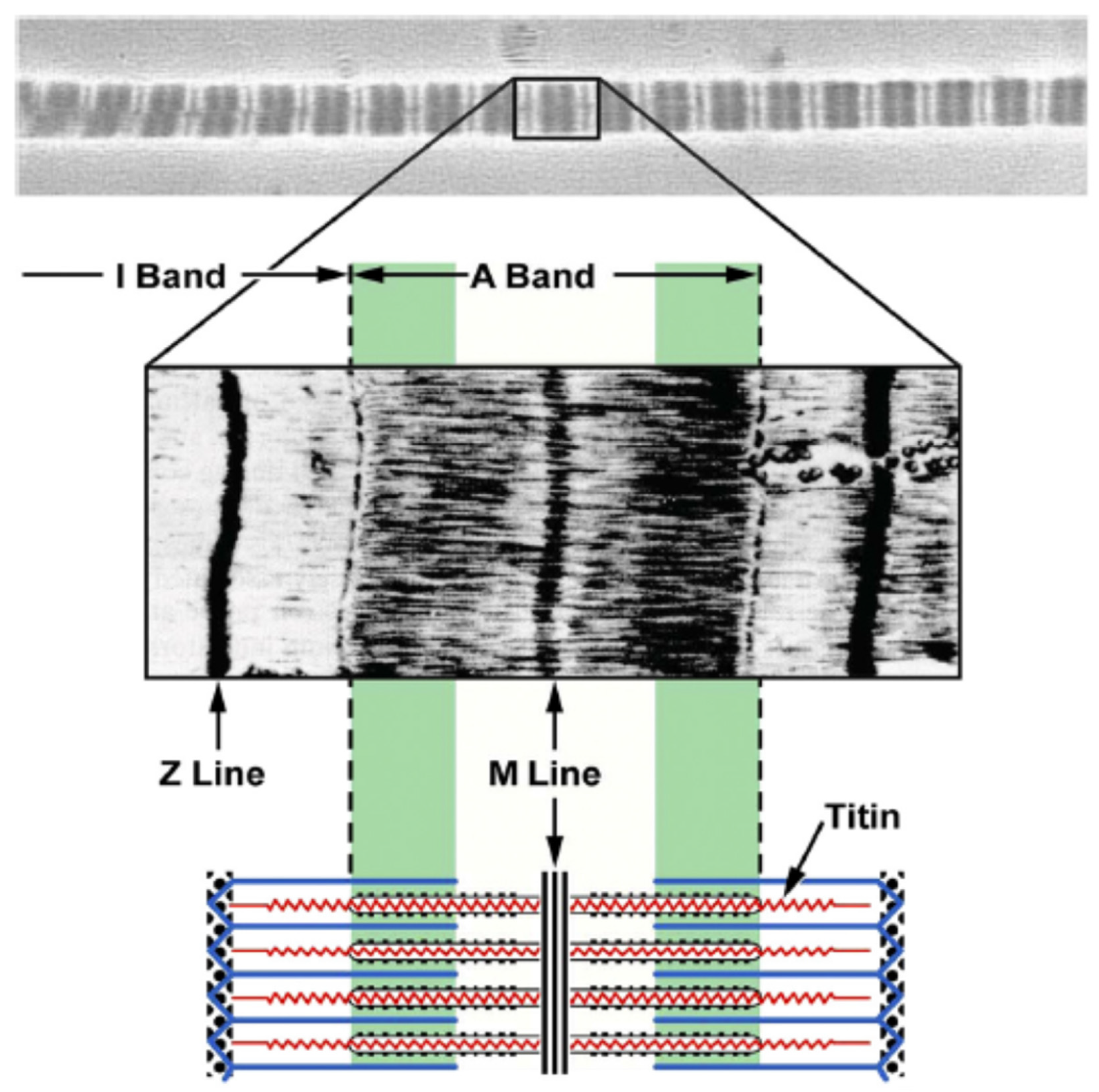
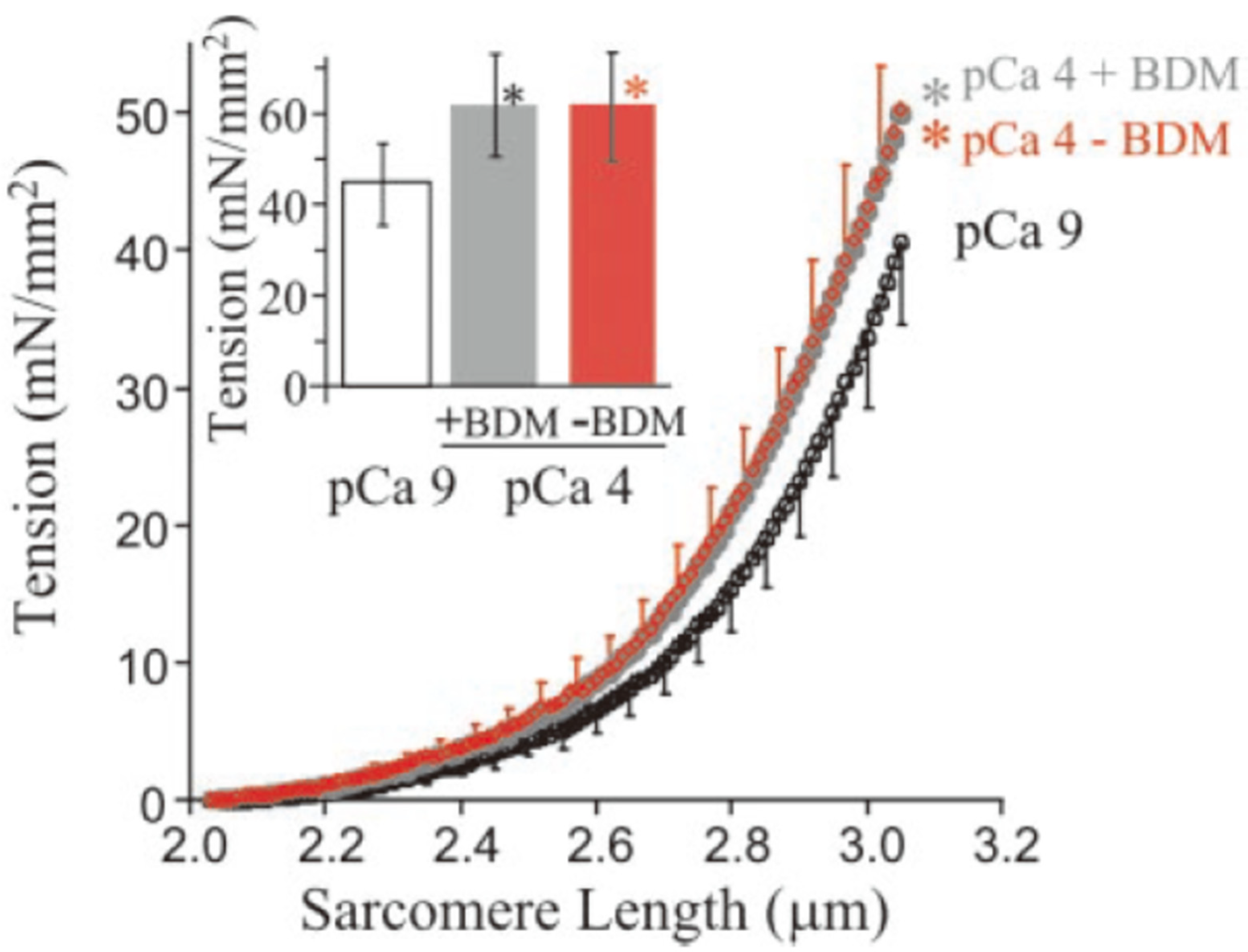
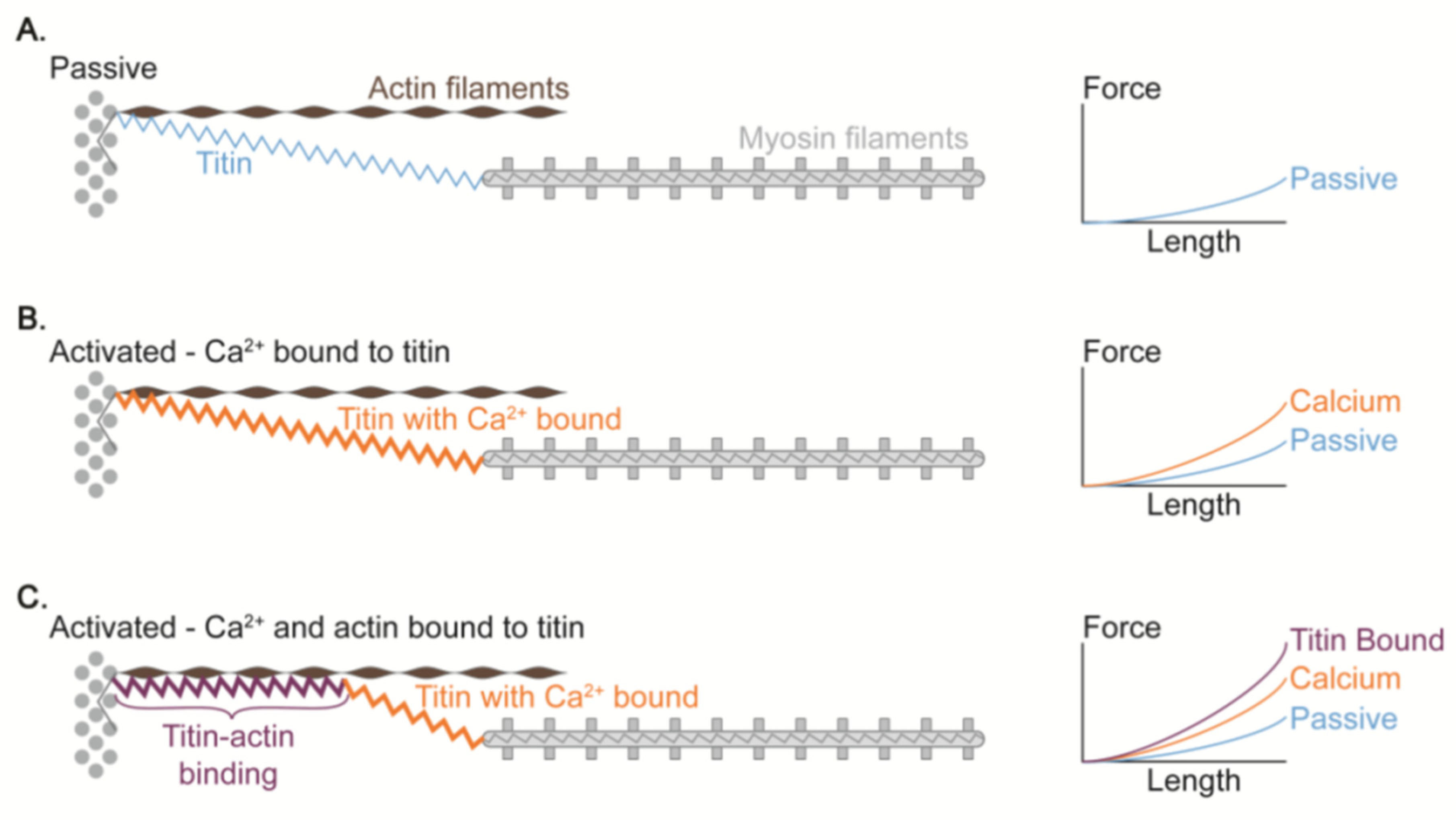
5. Titin Elasticity
5.1. Concept
5.2. Problems
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Huxley, A. Muscle structure and theories of contraction. Prog. Biophys. Biophys. Chem. 1957, 7, 255–318. [Google Scholar] [CrossRef]
- Rayment, I.; Holden, H.M.; Whittaker, M.; Yohn, C.B.; Lorenz, M.; Holmes, K.C.; Milligan, R.A. Structure of the actin-myosin complex and its implications for muscle contraction. Science 1993, 261, 58–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, A.V. The heat of shortening and the dynamic constants of muscle. Proc. R. Soc. Lond. 1938, 126, 136–195. [Google Scholar]
- Gordon, A.M.; Huxley, A.F.; Julian, F.J. The variation in isometric tension with sarcomere length in vertebrate muscle fibres. J. Physiol. 1966, 184, 170–192. [Google Scholar] [CrossRef] [PubMed]
- Abbott, B.C.; Aubert, X.M. The force exerted by active striated muscle during and after change of length. J. Physiol. 1952, 117, 77–86. [Google Scholar]
- Edman, K.A.; Elzinga, G.; Noble, M.I. Enhancement of mechanical performance by stretch during tetanic contractions of vertebrate skeletal muscle fibres. J. Physiol. 1978, 281, 139–155. [Google Scholar] [CrossRef]
- Edman, K.A.; Elzinga, G.; Noble, M.I. Residual force enhancement after stretch of contracting frog single muscle fibers. J. Gen. Physiol. 1982, 80, 769–784. [Google Scholar] [CrossRef]
- Lee, H.D.; Herzog, W. Force enhancement following muscle stretch of electrically stimulated and voluntarily activated human adductor pollicis. J. Physiol. 2002, 545, 321–330. [Google Scholar] [CrossRef]
- Lee, H.D.; Herzog, W. Force depression following muscle shortening of voluntarily activated and electrically stimulated human adductor pollicis. J. Physiol. 2003, 551, 993–1003. [Google Scholar] [CrossRef]
- Rassier, D.E.; Herzog, W.; Pollack, G.H. Stretch-induced force enhancement and stability of skeletal muscle myofibrils. In Molecular and Cellular Aspects of Muscle Contraction; Sugi, H., Ed.; Springer: Boston, MA, USA, 2003; Volume 538, pp. 501–515. [Google Scholar]
- Oskouei, A.E.; Herzog, W. Observations on force enhancement in submaximal voluntary contractions of human adductor pollicis muscle. J. Appl. Physiol. 2005, 98, 2087–2095. [Google Scholar] [CrossRef] [Green Version]
- Joumaa, V.; Leonard, T.R.; Herzog, W. Residual force enhancement in myofibrils and sarcomeres. Proc. Biol. Sci. 2008, 275, 1411–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hisey, B.; Leonard, T.R.; Herzog, W. Does residual force enhancement increase with increasing stretch magnitudes? J. Biomech. 2009, 42, 1488–1492. [Google Scholar] [CrossRef] [PubMed]
- Leonard, T.R.; DuVall, M.; Herzog, W. Force enhancement following stretch in a single sarcomere. Am. J. Physiol. 2010, 299, C1398–C1401. [Google Scholar] [CrossRef] [PubMed]
- Rassier, D.E.; Pavlov, I. Force produced by isolated sarcomeres and half-sarcomeres after an imposed stretch. Am. J. Physiol. Cell Physiol. 2012, 302, C240–C248. [Google Scholar] [CrossRef] [PubMed]
- Joumaa, V.; Herzog, W. Energy cost of force production is reduced after active stretch in skinned muscle fibres. J. Biomech. 2013, 46, 1135–1139. [Google Scholar] [CrossRef] [PubMed]
- Fukutani, A.; Herzog, W. Residual Force Enhancement Is Attenuated in a Shortening Magnitude-dependent Manner. Med. Sci. Sports Exerc. 2018, 50, 2007–2014. [Google Scholar] [CrossRef]
- Huxley, H.; Hanson, J. Changes in the cross-striations of muscle during contraction and stretch and their structural interpretation. Nature 1954, 173, 973–976. [Google Scholar] [CrossRef]
- Huxley, A.F.; Niedergerke, R. Structural changes in muscle during contraction; interference microscopy of living muscle fibres. Nature 1954, 173, 971–973. [Google Scholar] [CrossRef]
- Julian, F.J.; Morgan, D.L. The effect on tension of non-uniform distribution of length changes applied to frog muscle fibres. J. Physiol. 1979, 293, 379–392. [Google Scholar] [CrossRef]
- Morgan, D.L.; Whitehead, N.P.; Wise, A.K.; Gregory, J.E.; Proske, U. Tension changes in the cat soleus muscle following slow stretch or shortening of the contracting muscle. J. Physiol. 2000, 522, 503–513. [Google Scholar] [CrossRef]
- Peterson, D.; Rassier, D.; Herzog, W. Force enhancement in single skeletal muscle fibres on the ascending limb of the force-length relationship. J. Exp. Biol. 2004, 207, 2787–2791. [Google Scholar] [CrossRef] [PubMed]
- Herzog, W.; Leonard, T.R. The role of passive structures in force enhancement of skeletal muscles following active stretch. J. Biomech. 2005, 38, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Bullimore, S.R.; Leonard, T.R.; Rassier, D.E.; Herzog, W. History-dependence of isometric muscle force: Effect of prior stretch or shortening amplitude. J. Biomech. 2007, 40, 1518–1524. [Google Scholar] [CrossRef] [PubMed]
- Sugi, H.; Tsuchiya, T. Stiffness changes during enhancement and deficit of isometric force by slow length changes in frog skeletal muscle fibres. J. Physiol. 1988, 407, 215–229. [Google Scholar] [CrossRef]
- Morgan, D.L. An explanation for residual increased tension in striated muscle after stretch during contraction. Exp. Physiol. 1994, 79, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.V. The mechanics of active muscle. Proc. R. Soc. Lond. 1953, 141, 104–117. [Google Scholar]
- Allinger, T.L.; Herzog, W.; Epstein, M. Force-length properties in stable skeletal muscle fibers—theoretical considerations. J. Biomech. 1996, 29, 1235–1240. [Google Scholar] [CrossRef]
- Zahalak, G.I. Can muscle fibers be stable on the descending limbs of their sarcomere length-tension relations? J. Biomech. 1997, 30, 1179–1182. [Google Scholar] [CrossRef]
- Pun, C.; Syed, A.; Rassier, D.E. History-dependent properties of skeletal muscle myofibrils contracting along the ascending limb of the force-length relationship. Proc. Biol. Sci. 2010, 277, 475–484. [Google Scholar] [CrossRef]
- Rassier, D.; Herzog, W.; Wakeling, J.M.; Syme, D. Stretch-induced, steady-state force enhancement in single skeletal muscle fibers exceeds the isometric force at optimal fibre length. J. Biomech. 2003, 36, 1309–1316. [Google Scholar] [CrossRef]
- Schachar, R.; Herzog, W.; Leonard, T.R. The effects of muscle stretching and shortening on isometric forces on the descending limb of the force-length relationship. J. Biomech. 2004, 37, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, M.E.; Barretto, R.P.; Delp, S.L.; Schnitzer, M.J. Minimally invasive high-speed imaging of sarcomere contractile dynamics in mice and humans. Nature 2008, 454, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Moo, E.K.; Leonard, T.R.; Herzog, W. In Vivo Sarcomere Lengths Become More Non-uniform upon Activation in Intact Whole Muscle. Front. Physiol. 2017, 8, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moo, E.K.; Herzog, W. Single sarcomere contraction dynamics in a whole muscle. Sci. Rep. 2018, 8, 15235. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.; Jinha, A.; Herzog, W. The role of sarcomere length non-uniformities in residual force enhancement of skeletal muscle myofibrils. R. Soc. Open Sci. 2016, 3, 150657. [Google Scholar] [CrossRef]
- Chen, X.; Sanchez, G.N.; Schnitzer, M.J.; Delp, S.L. Changes in sarcomere lengths of the human vastus lateralis muscle with knee flexion measured using in vivo microendoscopy. J. Biomech. 2016, 49, 2989–2994. [Google Scholar] [CrossRef] [Green Version]
- Lichtwark, G.A.; Farris, D.J.; Chen, X.; Hodges, P.W.; Delp, S.L. Microendoscopy reveals positive correlation in multiscale length changes and variable sarcomere lengths across different regions of human muscle. J. Appl. Physiol. in press. [CrossRef]
- Noble, M.I. Enhancement of mechanical performance of striated muscle by stretch during contraction. Exp. Physiol. 1992, 77, 539–552. [Google Scholar] [CrossRef]
- Forcinito, M.; Epstein, M.; Herzog, W. Can a rheological muscle model predict force depression/enhancement? J. Biomech. 1998, 31, 1093–1099. [Google Scholar] [CrossRef]
- Rode, C.; Siebert, T.; Blickhan, R. Titin-induced force enhancement and force depression: A ‘sticky-spring’ mechanism in muscle contractions? J. Theor. Biol. 2009, 259, 350–360. [Google Scholar] [CrossRef]
- Nishikawa, K.C.; Monroy, J.A.; Uyeno, T.E.; Yeo, S.H.; Pai, D.K.; Lindstedt, S.L. Is titin a ‘winding filament’? A new twist on muscle contraction. Proc. Biol. Sci. 2012, 279, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Schappacher-Tilp, G.; Leonard, T.; Desch, G.; Herzog, W. A novel three-filament model of force generation in eccentric contraction of skeletal muscles. PLoS ONE 2015, 10, e0117634. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Natori, R.; Nonomura, Y. New elastic protein from muscle. Nature 1976, 262, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; McClure, J.; Tu, A. Titin: Major myofibrillar components of striated muscle. Proc. Natl. Acad. Sci. USA 1979, 76, 3698–3702. [Google Scholar] [CrossRef] [PubMed]
- Leonard, T.R.; Herzog, W. Regulation of muscle force in the absence of actin-myosin-based cross-bridge interaction. Am. J. Physiol. Cell Physiol. 2010, 299, C14–C20. [Google Scholar] [CrossRef] [Green Version]
- Freundt, J.K.; Linke, W.A. Titin as a force-generating muscle protein under regulatory control. J. Appl. Physiol. 2019, 126, 1474–1482. [Google Scholar] [CrossRef]
- Labeit, D.; Watanabe, K.; Witt, C.; Fujita, H.; Wu, Y.; Lahmers, S.; Funck, T.; Labeit, S.; Granzier, H. Calcium-dependent molecular spring elements in the giant protein titin. Proc. Natl. Acad. Sci. USA 2003, 100, 13716–13721. [Google Scholar] [CrossRef] [Green Version]
- DuVall, M.M.; Jinha, A.; Schappacher-Tilp, G.; Leonard, T.R.; Herzog, W. Differences in titin segmental elongation between passive and active stretch in skeletal muscle. J. Exp. Biol. 2017, 220, 4418–4425. [Google Scholar] [CrossRef] [Green Version]
- DuVall, M.M.; Gifford, J.L.; Amrein, M.; Herzog, W. Altered mechanical properties of titin immunoglobulin domain 27 in the presence of calcium. Eur. Biophys. J. 2013, 42, 301–307. [Google Scholar] [CrossRef]
- Krüger, M.; Kötter, S.; Grützner, A.; Lang, P.; Andresen, C.; Redfield, M.M.; Butt, E.; dos Remedios, C.G.; Linke, W.A. Protein kinase G modulates human myocardial passive stiffness by phosphorylation of the titin springs. Circ. Res. 2009, 104, 87–94. [Google Scholar] [CrossRef]
- Hamdani, N.; Krysiak, J.; Kreusser, M.M.; Neef, S.; Dos Remedios, C.G.; Maier, L.S.; Krüger, M.; Backs, J.; Linke, W.A. Crucial role for Ca2(+)/calmodulin-dependent protein kinase-II in regulating diastolic stress of normal and failing hearts via titin phosphorylation. Circ. Res. 2013, 112, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Herzog, W. The role of titin in eccentric muscle contraction. J. Exp. Biol. 2014, 217, 2825–2833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, K. Eccentric contraction: Unraveling mechanisms of force enhancement and energy conservation. J. Exp. Biol. 2016, 219, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Herzog, W.; Leonard, T.R. Force enhancement following stretching of skeletal muscle: A new mechanism. J. Exp. Biol. 2002, 205, 1275–1283. [Google Scholar]
- Rassier, D.E. Residual force enhancement in skeletal muscles: One sarcomere after the other. J. Muscle Res. Cell Motil. 2012, 33, 155–165. [Google Scholar] [CrossRef]
- Higuchi, H.; Takemori, S. Butanedione monoxime suppresses contraction and ATPase activity of rabbit skeletal muscle. J. Biochem. 1989, 105, 638–643. [Google Scholar] [CrossRef]
- Joumaa, V.; Rassier, D.E.; Leonard, T.R.; Herzog, W. Passive force enhancement in single myofibrils. Pflugers Arch. 2007, 455, 367–371. [Google Scholar] [CrossRef]
- Herzog, W. Why are muscles strong, and why do they require little energy in eccentric action? J. Sport Health Sci. 2018, 7, 255–264. [Google Scholar] [CrossRef]
- Linke, W.A.; Ivemeyer, M.; Labeit, S.; Hinssen, H.; Rüegg, J.C.; Gautel, M. Actin-titin interaction in cardiac myofibrils: Probing a physiological role. Biophys. J. 1997, 73, 905–919. [Google Scholar] [CrossRef]
- Kulke, M.; Fujita-Becker, S.; Rostkova, E.; Neagoe, C.; Labeit, D.; Manstein, D.J.; Gautel, M.; Linke, W.A. Interaction between PEVK-titin and actin filaments: Origin of a viscous force component in cardiac myofibrils. Circ. Res. 2001, 89, 874–881. [Google Scholar] [CrossRef]
- Bianco, P.; Nagy, A.; Kengyel, A.; Szatmári, D.; Mártonfalvi, Z.; Huber, T.; Kellermayer, M.S. Interaction forces between F-actin and titin PEVK domain measured with optical tweezers. Biophys. J. 2007, 93, 2102–2109. [Google Scholar] [CrossRef] [PubMed]
- Herzog, W.; Schappacher, G.; DuVall, M.; Leonard, T.R.; Herzog, J.A. Residual Force Enhancement Following Eccentric Contractions: A New Mechanism Involving Titin. Physiology 2016, 31, 300–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, W. The multiple roles of titin in muscle contraction and force production. Biophys. Rev. 2018, 10, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Cornachione, A.S.; Leite, F.; Bagni, M.A.; Rassier, D.E. The increase in non-cross-bridge forces after stretch of activated striated muscle is related to titin isoforms. Am. J. Physiol. Cell Physiol. 2016, 310, C19–C26. [Google Scholar] [CrossRef] [Green Version]
- Neagoe, C.; Opitz, C.A.; Makarenko, I.; Linke, W.A. Gigantic variety: Expression patterns of titin isoforms in striated muscles and consequences for myofibrillar passive stiffness. J. Muscle Res. Cell Motil. 2003, 24, 175–189. [Google Scholar] [CrossRef]
- Granzier, H.; Labeit, S. Structure-function relations of the giant elastic protein titin in striated and smooth muscle cells. Muscle Nerve 2007, 36, 740–755. [Google Scholar] [CrossRef]
- Fukutani, A.; Leonard, T.; Herzog, W. Does stretching velocity affect residual force enhancement? J. Biomech. 2019, 89, 143–147. [Google Scholar] [CrossRef]
- Nishikawa, K. Letter to the editor: “Titin-actin interaction: The report of its death was an exaggeration”. Am. J. Physiol. Cell Physiol. 2016, 310, C622. [Google Scholar] [CrossRef]
- Herzog, W. Letter to the editor: Comments on Cornachione et al. (2016): “The increase in non-cross-bridge forces after stretch of activated striated muscle is related to titin isoforms”. Am. J. Physiol. Cell Physiol. 2016, 311, C158–C159. [Google Scholar] [CrossRef]
- Rassier, D.E. Reply to “Letter to the editor: Comments on Cornachione et al. (2016): “The increase in non-cross-bridge forces after stretch of activated striated muscle is related to titin isoforms”. Am. J. Physiol. Cell Physiol. 2016, 311, C160–C161. [Google Scholar] [CrossRef]
- Rassier, D.E. Reply to “Letter to the editor: Titin-actin interaction: The report of its death was an exaggeration”. Am. J. Physiol. Cell Physiol. 2016, 310, C623. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, C.; Hudson, B.; Bogomolovas, J.; Zhu, Y.; Anderson, B.; Greaser, M.; Labeit, S.; Granzier, H. PKC phosphorylation of titin’s PEVK element: A novel and conserved pathway for modulating myocardial stiffness. Circ. Res. 2009, 105, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Giganti, D.; Yan, K.; Badilla, C.L.; Fernandez, J.M.; Alegre-Cebollada, J. Disulfide isomerization reactions in titin immunoglobulin domains enable a mode of protein elasticity. Nat. Commun. 2018, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, Y.; Iwamoto, H.; Kobayashi, T.; Sugi, H.; Tanaka, H.; Wakabayashi, K. Time-resolved X-ray diffraction studies on the effect of slow length changes on tetanized frog skeletal muscle. J. Physiol. 1988, 407, 231–241. [Google Scholar] [CrossRef]
- Herzog, W.; Leonard, T.R. The history dependence of force production in mammalian skeletal muscle following stretch-shortening and shortening-stretch cycles. J. Biomech. 2000, 33, 531–542. [Google Scholar] [CrossRef]
- Percario, V.; Boncompagni, S.; Protasi, F.; Pertici, I.; Pinzauti, F.; Caremani, M. Mechanical parameters of the molecular motor myosin II determined in permeabilised fibres from slow and fast skeletal muscles of the rabbit. J. Physiol. 2018, 596, 1243–1257. [Google Scholar] [CrossRef] [Green Version]
- Kellermayer, M.S.; Smith, S.B.; Granzier, H.L.; Bustamante, C. Folding-unfolding transitions in single titin molecules characterized with laser tweezers. Science 1997, 276, 1112–1126. [Google Scholar] [CrossRef]
- Ma, W.; Irving, T.C. X-ray Diffraction of Intact Murine Skeletal Muscle as a Tool for Studying the Structural Basis of Muscle Disease. J. Vis. Exp. 2019, 149. [Google Scholar] [CrossRef]
- Roig, M.; Macintyre, D.L.; Eng, J.J.; Narici, M.V.; Maganaris, C.N.; Reid, W.D. Preservation of eccentric strength in older adults: Evidence, mechanisms and implications for training and rehabilitation. Exp. Gerontol. 2010, 45, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Power, G.A.; Rice, C.L.; Vandervoort, A.A. Increased residual force enhancement in older adults is associated with a maintenance of eccentric strength. PLoS ONE 2012, 7, e48044. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukutani, A.; Herzog, W. Current Understanding of Residual Force Enhancement: Cross-Bridge Component and Non-Cross-Bridge Component. Int. J. Mol. Sci. 2019, 20, 5479. https://doi.org/10.3390/ijms20215479
Fukutani A, Herzog W. Current Understanding of Residual Force Enhancement: Cross-Bridge Component and Non-Cross-Bridge Component. International Journal of Molecular Sciences. 2019; 20(21):5479. https://doi.org/10.3390/ijms20215479
Chicago/Turabian StyleFukutani, Atsuki, and Walter Herzog. 2019. "Current Understanding of Residual Force Enhancement: Cross-Bridge Component and Non-Cross-Bridge Component" International Journal of Molecular Sciences 20, no. 21: 5479. https://doi.org/10.3390/ijms20215479
APA StyleFukutani, A., & Herzog, W. (2019). Current Understanding of Residual Force Enhancement: Cross-Bridge Component and Non-Cross-Bridge Component. International Journal of Molecular Sciences, 20(21), 5479. https://doi.org/10.3390/ijms20215479