p63 Is a Promising Marker in the Diagnosis of Unusual Skin Cancer

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
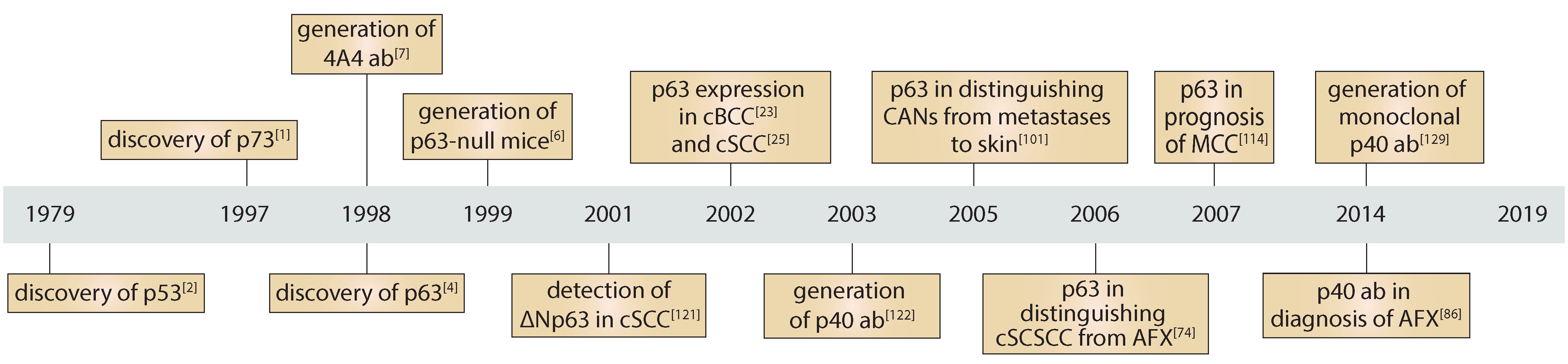
:1. Transcription Factor p63: Gene Structure and Function in Normal Skin and Skin Cancer
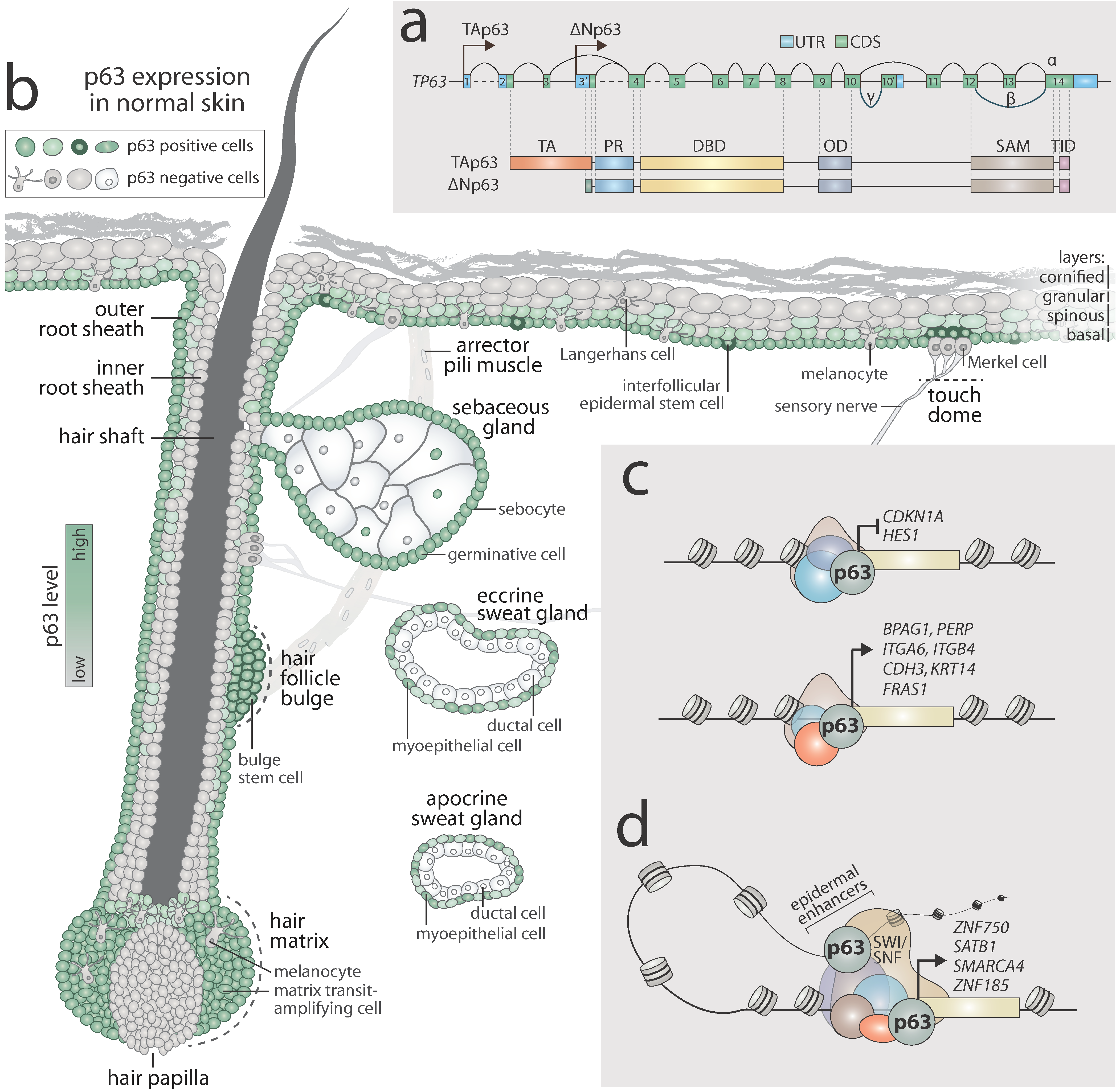
1.1. Gene Structure and Isoforms of p63
1.2. p63 Expression in Normal Skin
1.3. Transcription Regulation of Skin Development by p63
1.4. Transcription Regulation of Skin Cancer Mediated by p63
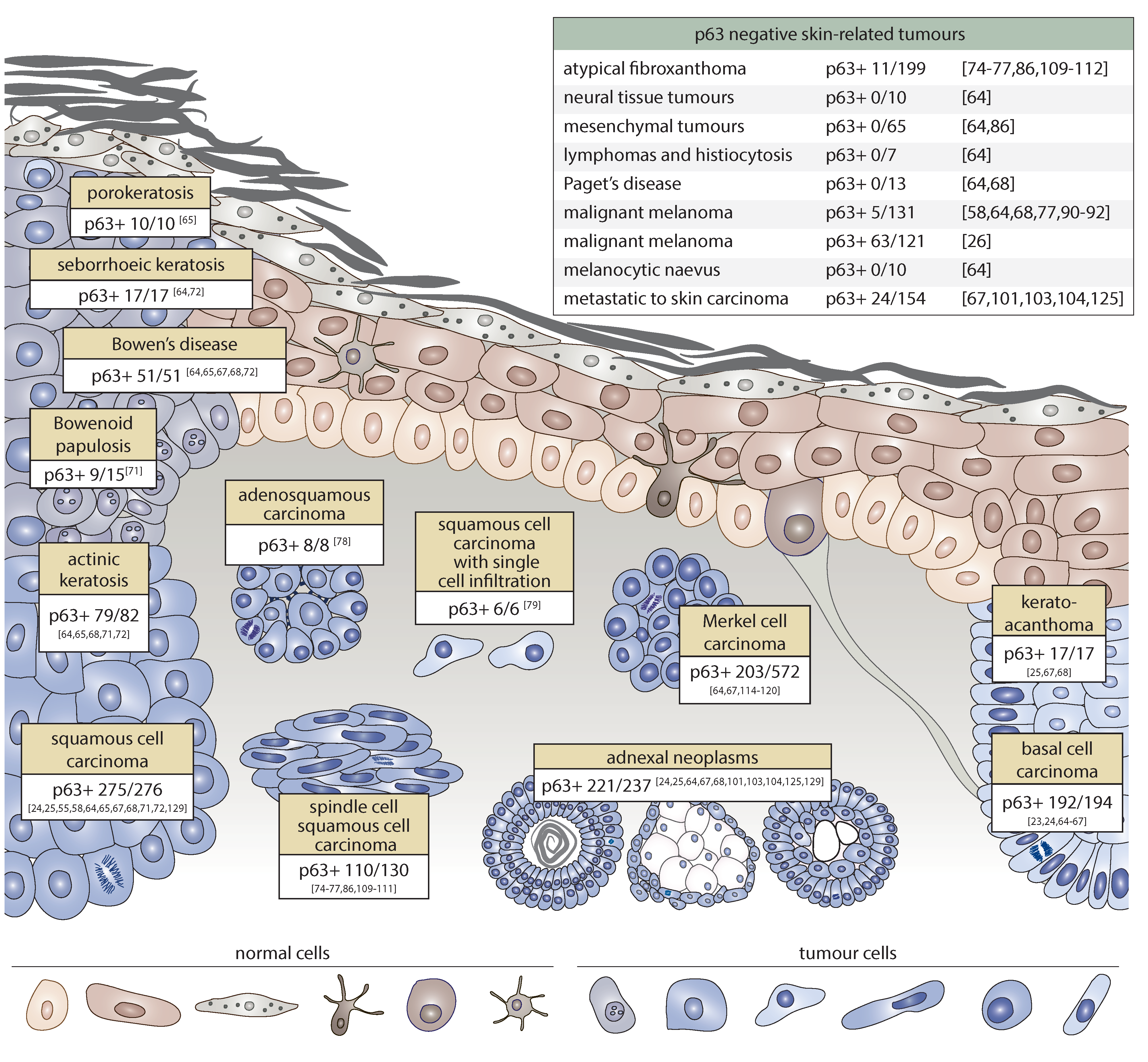
2. Expression Pattern of p63 in Skin Cancer
2.1. p63 Expression in Non-Melanoma Skin Cancer
2.2. p63 Expression in Melanoma
3. Diagnostic and Prognostic Value of p63 in Skin Cancer
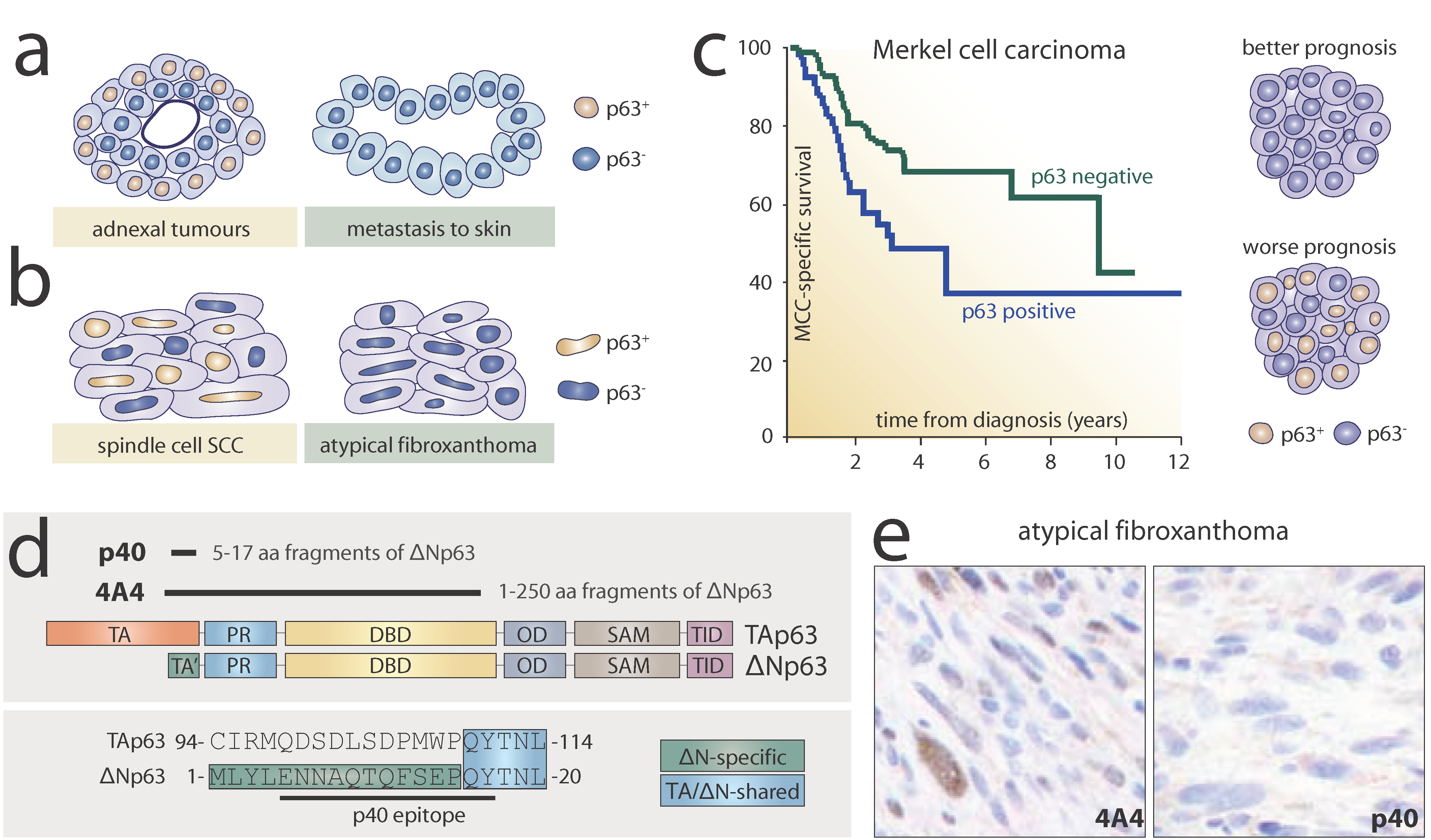
3.1. p63 in the Diagnosis of Metastases in Skin
3.2. p63 in the Diagnosis of Atypical Fibroxanthoma
3.3. p63 in the Prognosis of Merkel Cell Carcinoma
3.4. p40 as an Alternative to the Pan-p63 Antibody
4. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AFX | atypical fibroxanthoma |
| AK | actinic keratosis |
| BD | Bowen’s disease |
| BP | Bowenoid papulosis |
| CANs | cutaneous adnexal neoplasms |
| cBCC | cutaneous basal cell carcinoma |
| CK | cytokeratin |
| cSCC | cutaneous squamous cell carcinoma |
| cSCSCC | cutaneous spindle cell SCC |
| KA | keratoacanthoma |
| MCC | Merkel cell carcinoma |
| MM | malignant melanoma |
| PCR | polymerase chain reaction |
| PK | porokeratosis |
| SK | seborrhoeic keratosis |
| UV | ultraviolet |
References
- Kaghad, M.; Bonnet, H.; Yang, A.; Creancier, L.; Biscan, J.C.; Valent, A.; Minty, A.; Chalon, P.; Lelias, J.M.; Dumont, X.; et al. Monoallelically Expressed Gene Related to P53 at 1p36, a Region Frequently Deleted in Neuroblastoma and Other Human Cancers. Cell 1997, 90, 809–819. [Google Scholar] [CrossRef]
- Levine, A.J.; Oren, M. The First 30 Years of P53: Growing Ever More Complex. Nat. Rev. Cancer 2009, 9, 749. [Google Scholar] [CrossRef] [PubMed]
- Melino, G.; Lu, X.; Gasco, M.; Crook, T.; Knight, R.A. Functional Regulation of P73 and P63: Development and Cancer. Trends Biochem. Sci. 2003, 28, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Kaghad, M.; Wang, Y.; Gillett, E.; Fleming, M.D.; Dötsch, V.; Andrews, N.C.; Caput, D.; McKeon, F. P63, a P53 Homolog at 3Q27–29, Encodes Multiple Products with Transactivating, Death-Inducing, and Dominant-Negative Activities. Mol. Cell 1998, 2, 305–316. [Google Scholar] [CrossRef]
- Donehower, L.A. The P53-Deficient Mouse: A Model for Basic and Applied Cancer Studies. Semin. Cancer Biol. 1996, 7, 269–278. [Google Scholar] [CrossRef]
- Yang, A.; Schweitzer, R.; Sun, D.; Kaghad, M.; Walker, N.; Bronson, R.T.; Tabin, C.; Sharpe, A.; Caput, D.; Crum, C.; et al. P63 Is Essential for Regenerative Proliferation in Limb, Craniofacial and Epithelial Development. Nature 1999, 398, 714–718. [Google Scholar] [CrossRef]
- Yang, A.; Walker, N.; Bronson, R.; Kaghad, M.; Oosterwegel, M.; Bonnin, J.; Vagner, C.; Bonnet, H.; Dikkes, P.; Sharpe, A.; et al. P73-Deficient Mice Have Neurological, Pheromonal and Inflammatory Defects but Lack Spontaneous Tumours. Nature 2000, 404, 99. [Google Scholar] [CrossRef]
- Petitjean, A.; Ruptier, C.; Tribollet, V.; Hautefeuille, A.; Chardon, F.; Cavard, C.; Puisieux, A.; Hainaut, P.; De Fromentel, C.C. Properties of the Six Isoforms of P63: P53-like Regulation in Response to Genotoxic Stress and Cross Talk with ΔNp73. Carcinogenesis 2008, 29, 273–281. [Google Scholar] [CrossRef]
- Duijf, P.H.G. Gain-of-Function Mutation in ADULT Syndrome Reveals the Presence of a Second Transactivation Domain in P63. Hum. Mol. Genet. 2002, 11, 799–804. [Google Scholar] [CrossRef]
- Yang, A.; McKeon, F. P63 and P73: P53 Mimics, Menaces and More. Nat. Rev. Mol. Cell Biol. 2000, 1, 199. [Google Scholar] [CrossRef]
- McGrath, J.A. Hay-Wells Syndrome Is Caused by Heterozygous Missense Mutations in the SAM Domain of P63. Hum. Mol. Genet. 2001, 10, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Der Ou, H.; Löhr, F.; Vogel, V.; Mäntele, W.; Dötsch, V. Structural Evolution of C-Terminal Domains in the P53 Family. EMBO J. 2007, 26, 3463–3473. [Google Scholar]
- Serber, Z.; Lai, H.C.; Yang, A.; Ou, H.D.; Sigal, M.S.; Kelly, A.E.; Darimont, B.D.; Duijf, P.H.G.; van Bokhoven, H.; McKeon, F.; et al. A C-Terminal Inhibitory Domain Controls the Activity of P63 by an Intramolecular Mechanism. Mol. Cell. Biol. 2002, 22, 8601–8611. [Google Scholar] [CrossRef] [PubMed]
- Straub, W.E.; Weber, T.A.; Schäfer, B.; Candi, E.; Durst, F.; Ou, H.D.; Rajalingam, K.; Melino, G.; Dötsch, V. The C-Terminus of P63 Contains Multiple Regulatory Elements with Different Functions. Cell Death Dis. 2010, 1, e5. [Google Scholar] [CrossRef] [PubMed]
- Tuppi, M.; Kehrloesser, S.; Coutandin, D.W.; Rossi, V.; Luh, L.M.; Strubel, A.; Hötte, K.; Hoffmeister, M.; Schäfer, B.; De Oliveira, T.; et al. Oocyte DNA Damage Quality Control Requires Consecutive Interplay of CHK2 and CK1 to Activate P63. Nat. Struct. Mol. Biol. 2018, 25, 261. [Google Scholar] [CrossRef]
- Crum, C.P.; McKeon, F.D. P63 in Epithelial Survival, Germ Cell Surveillance, and Neoplasia. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 349–371. [Google Scholar] [CrossRef]
- Candi, E.; Rufini, A.; Terrinoni, A.; Giamboi-Miraglia, A.; Lena, A.M.; Mantovani, R.; Knight, R.; Melino, G. ΔNp63 Regulates Thymic Development through Enhanced Expression of FgfR2 and Jag2. Proc. Natl. Acad. Sci. USA 2007, 104, 11999–12004. [Google Scholar] [CrossRef]
- Candi, E.; Cipollone, R.; Di Val Cervo, P.R.; Gonfloni, S.; Melino, G.; Knight, R. P63 in Epithelial Development. Cell. Mol. Life Sci. 2008, 65, 3126. [Google Scholar] [CrossRef]
- Candi, E.; Rufini, A.; Terrinoni, A.; Dinsdale, D.; Ranalli, M.; Paradisi, A.; De Laurenzi, V.; Spagnoli, L.G.; Catani, M.V.; Ramadan, S.; et al. Differential Roles of P63 Isoforms in Epidermal Development: Selective Genetic Complementation in P63 Null Mice. Cell Death Differ. 2006, 13, 1037. [Google Scholar] [CrossRef]
- Romano, R.-A.; Smalley, K.; Magraw, C.; Serna, V.A.; Kurita, T.; Raghavan, S.; Sinha, S. ΔNp63 Knockout Mice Reveal Its Indispensable Role as a Master Regulator of Epithelial Development and Differentiation. J. Cell Sci. 2012, 139, 772–782. [Google Scholar]
- Vanbokhoven, H.; Melino, G.; Candi, E.; Declercq, W. P63, a Story of Mice and Men. J. Investig. Dermatol. 2011, 131, 1196–1207. [Google Scholar] [CrossRef] [PubMed]
- Rufini, A.; Weil, M.; McKeon, F.; Barlattani, A.; Melino, G.; Candi, E. P63 Protein Is Essential for the Embryonic Development of Vibrissae and Teeth. Biochem. Biophys. Res. Commun. 2006, 340, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Di Como, C.J.; Urist, M.J.; Babayan, I.; Drobnjak, M.; Hedvat, C.V.; Teruya-Feldstein, J.; Pohar, K.; Hoos, A.; Cordon-Cardo, C. P63 Expression Profiles in Human Normal and Tumor Tissues. Clin. Cancer Res. 2002, 8, 494–501. [Google Scholar] [PubMed]
- Tsujita-Kyutoku, M.; Kiuchi, K.; Danbara, N.; Yuri, T.; Senzaki, H.; Tsubura, A. P63 Expression in Normal Human Epidermis and Epidermal Appendages and Their Tumors. J. Cutan. Pathol. 2003, 30, 11–17. [Google Scholar] [CrossRef]
- Reis-Filho, J.S.; Torio, B.; Albergaria, A.; Schmitt, F.C. P63 Expression in Normal Skin and Usual Cutaneous Carcinomas. J. Cutan. Pathol. 2002, 29, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Matin, R.N.; Chikh, A.; Law Pak Chong, S.; Mesher, D.; Graf, M.; Sanza, P.; Senatore, V.; Scatolini, M.; Moretti, F.; Leigh, I.M.; et al. P63 Is an Alternative P53 Repressor in Melanoma That Confers Chemoresistance and a Poor Prognosis. J. Exp. Med. 2013, 210, 581–603. [Google Scholar] [CrossRef] [PubMed]
- ENCODE Project Consortium. An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Dohata, A.; Chambers, J.K.; Uchida, K.; Nakazono, S.; Kinoshita, Y.; Nibe, K.; Nakayama, H. Clinical and Pathologic Study of Feline Merkel Cell Carcinoma with Immunohistochemical Characterization of Normal and Neoplastic Merkel Cells. Vet. Pathol. 2015, 52, 1012–1018. [Google Scholar] [CrossRef]
- Truong, A.B.; Kretz, M.; Ridky, T.W.; Kimmel, R.; Khavari, P.A. P63 Regulates Proliferation and Differentiation of Developmentally Mature Keratinocytes. Genes Dev. 2006, 20, 3185–3197. [Google Scholar] [CrossRef]
- Nguyen, B.C.; Lefort, K.; Mandinova, A.; Antonini, D.; Devgan, V.; Della Gatta, G.; Koster, M.I.; Zhang, Z.; Wang, J.; Di Vignano, A.T.; et al. Cross-Regulation between Notch and P63 in Keratinocyte Commitment to Differentiation. Genes Dev. 2006, 20, 1028–1042. [Google Scholar] [CrossRef]
- Osada, M.; Nagakawa, Y.; Park, H.L.; Yamashita, K.; Wu, G.; Kim, M.S.; Fomenkov, A.; Trink, B.; Sidransky, D. P63-Specific Activation of the BPAG-1e Promoter. J. Investig. Dermatol. 2005, 125, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Ihrie, R.A.; Marques, M.R.; Nguyen, B.T.; Horner, J.S.; Papazoglu, C.; Bronson, R.T.; Mills, A.A.; Attardi, L.D. Perp Is a P63-Regulated Gene Essential for Epithelial Integrity. Cell 2005, 120, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Carroll, D.K.; Carroll, J.S.; Leong, C.-O.; Cheng, F.; Brown, M.; Mills, A.A.; Brugge, J.S.; Ellisen, L.W. P63 Regulates an Adhesion Programme and Cell Survival in Epithelial Cells. Nat. Cell Biol. 2006, 8, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Wajid, M.; Shapiro, L.; Christiano, A.M. P-Cadherin Is a P63 Target Gene with a Crucial Role in the Developing Human Limb Bud and Hair Follicle. Development 2008, 135, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Koster, M.I.; Dai, D.; Marinari, B.; Sano, Y.; Costanzo, A.; Karin, M.; Roop, D.R. P63 Induces Key Target Genes Required for Epidermal Morphogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 3255–3260. [Google Scholar] [CrossRef] [Green Version]
- Amelio, I.; Melino, G.; Candi, E. P63 Adjusts Sugar Taste of Epidermal Layers. J. Investig. Dermatol. 2017, 137, 1204–1206. [Google Scholar] [CrossRef] [Green Version]
- Viticchiè, G.; Agostini, M.; Lena, A.M.; Mancini, M.; Zhou, H.; Zolla, L.; Dinsdale, D.; Saintigny, G.; Melino, G.; Candi, E. P63 Supports Aerobic Respiration through Hexokinase II. Proc. Natl. Acad. Sci. USA 2015, 112, 11577–11582. [Google Scholar] [CrossRef] [Green Version]
- Latina, A.; Viticchiè, G.; Lena, A.M.; Piro, M.C.; Annicchiarico-Petruzzelli, M.; Melino, G.; Candi, E. ΔNp63 Targets Cytoglobin to Inhibit Oxidative Stress-Induced Apoptosis in Keratinocytes and Lung Cancer. Oncogene 2016, 35, 1493–1503. [Google Scholar] [CrossRef]
- Ramsey, M.R.; He, L.; Forster, N.; Ory, B.; Ellisen, L.W. Physical Association of HDAC1 and HDAC2 with P63 Mediates Transcriptional Repression and Tumor Maintenance in Squamous Cell Carcinoma. Cancer Res. 2011, 71, 4373–4379. [Google Scholar] [CrossRef] [Green Version]
- LeBoeuf, M.; Terrell, A.; Trivedi, S.; Sinha, S.; Epstein, J.A.; Olson, E.N.; Morrisey, E.E.; Millar, S.E. Hdac1 and Hdac2 Act Redundantly to Control P63 and P53 Functions in Epidermal Progenitor Cells. Dev. Cell 2010, 19, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Lin-Shiao, E.; Lan, Y.; Coradin, M.; Anderson, A.; Donahue, G.; Simpson, C.L.; Sen, P.; Saffie, R.; Busino, L.; Garcia, B.A.; et al. KMT2D Regulates P63 Target Enhancers to Coordinate Epithelial Homeostasis. Genes Dev. 2018, 32, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Sen, G.L.; Boxer, L.D.; Webster, D.E.; Bussat, R.T.; Qu, K.; Zarnegar, B.J.; Johnston, D.; Siprashvili, Z.; Khavari, P.A. ZNF750 Is a P63 Target Gene That Induces KLF4 to Drive Terminal Epidermal Differentiation. Dev. Cell 2012, 22, 669–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boxer, L.D.; Barajas, B.; Tao, S.; Zhang, J.; Khavari, P.A. Znf750 Interacts with KLF4 and RCOR1, KDM1A, And CTBP1/2 Chromatin Regulators to Repress Epidermal Progenitor Genes and Induce Differentiation Genes. Genes Dev. 2014, 28, 2013–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fessing, M.Y.; Mardaryev, A.N.; Gdula, M.R.; Sharov, A.A.; Sharova, T.Y.; Rapisarda, V.; Gordon, K.B.; Smorodchenko, A.D.; Poterlowicz, K.; Ferone, G.; et al. P63 Regulates Satb1 to Control Tissue-Specific Chromatin Remodeling during Development of the Epidermis. J. Cell Biol. 2011, 194, 825–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardaryev, A.N.; Gdula, M.R.; Yarker, J.L.; Emelianov, V.N.; Poterlowicz, K.; Sharov, A.A.; Sharova, T.Y.; Scarpa, J.A.; Chambon, P.; Botchkarev, V.A.; et al. P63 and Brg1 Control Developmentally Regulated Higher-Order Chromatin Remodelling at the Epidermal Differentiation Complex Locus in Epidermal Progenitor Cells. Development 2014, 141, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Rubin, A.J.; Barajas, B.C.; Furlan-Magaril, M.; Lopez-Pajares, V.; Mumbach, M.R.; Howard, I.; Kim, D.S.; Boxer, L.D.; Cairns, J.; Spivakov, M.; et al. Lineage-Specific Dynamic and Pre-Established Enhancer–Promoter Contacts Cooperate in Terminal Differentiation. Nat. Genet. 2017, 49, 1522. [Google Scholar] [CrossRef]
- Bao, X.; Rubin, A.J.; Qu, K.; Zhang, J.; Giresi, P.G.; Chang, H.Y.; Khavari, P.A. A Novel ATAC-Seq Approach Reveals Lineage-Specific Reinforcement of the Open Chromatin Landscape via Cooperation between BAF and P63. Genome Biol. 2015, 16, 284. [Google Scholar] [CrossRef] [Green Version]
- Kouwenhoven, E.N.; Oti, M.; Niehues, H.; van Heeringen, S.J.; Schalkwijk, J.; Stunnenberg, H.G.; van Bokhoven, H.; Zhou, H. Transcription Factor P63 Bookmarks and Regulates Dynamic Enhancers during Epidermal Differentiation. EMBO Rep. 2015, 16, 863–878. [Google Scholar] [CrossRef]
- Smirnov, A.; Lena, A.M.; Cappello, A.; Panatta, E.; Anemona, L.; Bischetti, S.; Annicchiarico-Petruzzelli, M.; Mauriello, A.; Melino, G.; Candi, E. ZNF185 Is a P63 Target Gene Critical for Epidermal Differentiation and Squamous Cell Carcinoma Development. Oncogene 2018, 38, 1625. [Google Scholar] [CrossRef] [Green Version]
- Gatti, V.; Fierro, C.; Annicchiarico-Petruzzelli, M.; Melino, G.; Peschiaroli, A. ΔNp63 in Squamous Cell Carcinoma: Defining the Oncogenic Routes Affecting Epigenetic Landscape and Tumour Microenvironment. Mol. Oncol. 2019, 13, 981–1001. [Google Scholar] [CrossRef] [Green Version]
- Moses, M.A.; George, A.L.; Sakakibara, N.; Mahmood, K.; Ponnamperuma, R.M.; King, K.E.; Weinberg, W.C. Molecular Mechanisms of P63-Mediated Squamous Cancer Pathogenesis. Int. J. Mol. Sci. 2019, 20, 3590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, L.; Ponnamperuma, R.M.; Jay, S.; Ricci, M.S.; Weinberg, W.C. Dysregulated ΔNp63α Inhibits Expression of Ink4a/Arf, Blocks Senescence, and Promotes Malignant Conversion of Keratinocytes. PLoS ONE 2011, 6, e21877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyes, W.M.; Pecoraro, M.; Aranda, V.; Vernersson-Lindahl, E.; Li, W.; Vogel, H.; Guo, X.; Garcia, E.L.; Michurina, T.V.; Enikolopov, G.; et al. Δnp63α Is an Oncogene That Targets Chromatin Remodeler Lsh to Drive Skin Stem Cell Proliferation and Tumorigenesis. Cell Stem Cell 2011, 8, 164–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikh, A.; Matin, R.N.H.; Senatore, V.; Hufbauer, M.; Lavery, D.; Raimondi, C.; Ostano, P.; Mello-Grand, M.; Ghimenti, C.; Bahta, A.; et al. IASPP/P63 Autoregulatory Feedback Loop Is Required for the Homeostasis of Stratified Epithelia. EMBO J. 2011, 30, 4261–4273. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.J.; Patel, A.; Purdie, K.J.; Wang, J.; Rizvi, H.; Hufbauer, M.; Ostano, P.; Akgül, B.; Chiorino, G.; Harwood, C.A.; et al. Epigenetic Regulation of IASPP-P63 Feedback Loop in Cutaneous Squamous Cell Carcinoma. J. Investig. Dermatol. 2019, 139, 1658–1671. [Google Scholar] [CrossRef]
- Hazawa, M.; Lin, D.; Kobayashi, A.; Jiang, Y.; Xu, L.; Dewi, F.R.P.; Mohamed, M.S.; Hartono; Nakada, M.; Meguro-Horike, M.; et al. ROCK-dependent Phosphorylation of NUP 62 Regulates P63 Nuclear Transport and Squamous Cell Carcinoma Proliferation. EMBO Rep. 2018, 19, 73–88. [Google Scholar] [CrossRef] [Green Version]
- DeYoung, M.P.; Johannessen, C.M.; Leong, C.O.; Faquin, W.; Rocco, J.W.; Ellisen, L.W. Tumor-Specific P73 up-Regulation Mediates P63 Dependence in Squamous Cell Carcinoma. Cancer Res. 2006, 66, 9362–9368. [Google Scholar] [CrossRef] [Green Version]
- Cordani, N.; Pozzi, S.; Martynova, E.; Fanoni, D.; Borrelli, S.; Alotto, D.; Castagnoli, C.; Berti, E.; Viganò, M.A.; Mantovani, R. Mutant P53 Subverts P63 Control over KLF4 Expression in Keratinocytes. Oncogene 2011, 30, 922. [Google Scholar] [CrossRef] [Green Version]
- Bornachea, O.; López-Calderón, F.F.; Dueñas, M.; Segrelles, C.; Lorz, C.; Suárez-Cabrera, C.; Marañón, M.; Paradela-Dobarro, B.; Santos, M.; Paramio, J.M. The Downregulation of ΔNp63 in P53-Deficient Mouse Epidermal Tumors Favors Metastatic Behavior. Oncotarget 2015, 6, 24230. [Google Scholar] [CrossRef]
- Anna, B.; Blazej, Z.; Jacqueline, G.; Andrew, C.J.; Jeffrey, R.; Andrzej, S. Mechanism of UV-Related Carcinogenesis and Its Contribution to Nevi/Melanoma. Expert Rev. Dermatol. 2007, 2, 451–469. [Google Scholar]
- Gordon, R. Skin Cancer: An Overview of Epidemiology and Risk Factors. Semin. Oncol. Nurs. 2013, 29, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E.H. Basal Cell Carcinomas: Attack of the Hedgehog. Nat. Rev. Cancer 2008, 8, 743. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.C.; Eberl, M.; Vagnozzi, A.N.; Belkadi, A.; Veniaminova, N.A.; Verhaegen, M.E.; Bichakjian, C.K.; Ward, N.L.; Dlugosz, A.A.; Wong, S.Y. Basal Cell Carcinoma Preferentially Arises from Stem Cells within Hair Follicle and Mechanosensory Niches. Cell Stem Cell 2015, 16, 400–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, Y.; Tamura, A.; Kamiya, M.; Fukuda, T.; Ishikawa, O. Immunohistochemical Analyses of P63 Expression in Cutaneous Tumours. Br. J. Dermatol. 2005, 153, 1230–1232. [Google Scholar] [CrossRef] [PubMed]
- Park, H.R.; Soo, K.M.; Hyun, D.C.; Kwang, H.K.; Hyung, S.S.; Young, E.P. Expression Profiles of P63, P53, Survivin, and HTERT in Skin Tumors. J. Cutan. Pathol. 2004, 31, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Bircan, S.; Candir, O.; Kapucoglu, N.; Baspinar, S. The Expression of P63 in Basal Cell Carcinomas and Association with Histological Differentiation. J. Cutan. Pathol. 2006, 33, 293–298. [Google Scholar] [CrossRef]
- Kanitakis, J.; Chouvet, B. Expression of P63 in Cutaneous Metastases. Am. J. Clin. Pathol. 2007, 128, 753–758. [Google Scholar] [CrossRef]
- Sakiz, D.; Turkmenoglu, T.T.; Kabukcuoglu, F. The Expression of P63 and P53 in Keratoacanthoma and Intraepidermal and Invasive Neoplasms of the Skin. Pathol. Res. Pract. 2009, 205, 589–594. [Google Scholar] [CrossRef]
- Burton, K.A.; Ashack, K.A.; Khachemoune, A. Cutaneous Squamous Cell Carcinoma: A Review of High-Risk and Metastatic Disease. Am. J. Clin. Dermatol. 2016, 17, 491–508. [Google Scholar] [CrossRef]
- Lapouge, G.; Youssef, K.K.; Vokaer, B.; Achouri, Y.; Michaux, C.; Sotiropoulou, P.A.; Blanpain, C. Identifying the Cellular Origin of Squamous Skin Tumors. Proc. Natl. Acad. Sci. USA 2011, 108, 7431–7436. [Google Scholar] [CrossRef] [Green Version]
- Abbas, O.; Richards, J.E.; Yaar, R.; Mahalingam, M. Stem Cell Markers (Cytokeratin 15, Cytokeratin 19 and P63) in in Situ and Invasive Cutaneous Epithelial Lesions. Mod. Pathol. 2011, 24, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrone, D.A.; Yoo, S.; Chipps, L.K.; Moy, R.L. The Expression of P63 in Actinic Keratosis, Seborrheic Keratosis, and Cutaneous Squamous Cell Carcinomas. Dermatol. Surg. 2004, 30, 1299–1302. [Google Scholar] [PubMed]
- Cassarino, D.S.; DeRienzo, D.P.; Barr, R.J. Cutaneous Squamous Cell Carcinoma: A Comprehensive Clinicopathologic Classification—Part Two. J. Cutan. Pathol. 2006, 33, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Dotto, J.E.; Glusac, E.J. P63 Is a Useful Marker for Cutaneous Spindle Cell Squamous Cell Carcinoma. J. Cutan. Pathol. 2006, 33, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.M.; Saenger, J.S.; Fadare, O. Diagnostic Utility of P63 and CD10 in Distinguishing Cutaneous Spindle Cell/Sarcomatoid Squamous Cell Carcinomas and Atypical Fibroxanthomas. Int. J. Clin. Exp. Pathol. 2008, 1, 524. [Google Scholar] [PubMed]
- Gleason, B.C.; Calder, K.B.; Cibull, T.L.; Thomas, A.B.; Billings, S.D.; Morgan, M.B.; Hiatt, K.M.; Smoller, B.R. Utility of P63 in the Differential Diagnosis of Atypical Fibroxanthoma and Spindle Cell Squamous Cell Carcinoma. J. Cutan. Pathol. 2009, 36, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Kanner, W.A.; Brill, L.B.; Patterson, J.W.; Wick, M.R. CD10, P63 and CD99 Expression in the Differential Diagnosis of Atypical Fibroxanthoma, Spindle Cell Squamous Cell Carcinoma and Desmoplastic Melanoma. J. Cutan. Pathol. 2010, 37, 744–750. [Google Scholar] [CrossRef]
- Ko, C.J.; Leffell, D.J.; McNiff, J.M. Adenosquamous Carcinoma: A Report of Nine Cases with P63 and Cytokeratin 5/6 Staining. J. Cutan. Pathol. 2009, 36, 448–452. [Google Scholar] [CrossRef]
- Ko, C.J.; McNiff, J.M.; Glusac, E.J. Squamous Cell Carcinomas with Single Cell Infiltration: A Potential Diagnostic Pitfall and the Utility of MNF116 and P63. J. Cutan. Pathol. 2008, 35, 353–357. [Google Scholar] [CrossRef]
- Roh, N.K.; Hahn, H.J.; Lee, Y.W.; Choe, Y.B.; Ahn, K.J. Clinical and Histopathological Investigation of Seborrheic Keratosis. Ann. Dermatol. 2016, 28, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Zargaran, M.; Baghaei, F. A Clinical, Histopathological and Immunohistochemical Approach to the Bewildering Diagnosis of Keratoacanthoma. J. Dent. 2014, 15, 91. [Google Scholar]
- Sertznig, P.; von Felbert, V.; Megahed, M. Porokeratosis: Present Concepts. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Roewert-Huber, J.; Stockfleth, E.; Kerl, H. Pathology and Pathobiology of Actinic (Solar) Keratosis—An Update. Br. J. Dermatol. 2007, 157, 18–20. [Google Scholar] [CrossRef] [PubMed]
- Neagu, T.P.; Ţigliş, M.; Botezatu, D.; Enache, V.; Cobilinschi, C.O.; Vâlcea-Precup, M.S.; Grinţescu, I.M. Clinical, Histological and Therapeutic Features of Bowen’s Disease. Rom. J. Morphol. Embryol. 2017, 58, 33–40. [Google Scholar] [PubMed]
- Dubina, M.; Goldenberg, G. Viral-Associated Nonmelanoma Skin Cancers: A Review. Am. J. Dermatopathol. 2009, 31, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Ha Lan, T.T.; Chen, S.J.T.; Arps, D.P.; Fullen, D.R.; Patel, R.M.; Siddiqui, J.; Carskadon, S.; Palanisamy, N.; Harms, P.W. Expression of the P40 Isoform of P63 Has High Specificity for Cutaneous Sarcomatoid Squamous Cell Carcinoma. J. Cutan. Pathol. 2014, 41, 831–838. [Google Scholar] [CrossRef]
- Mai, R.; Zhou, S.; Zhou, S.; Zhong, W.; Hong, L.; Wang, Y.; Lu, S.; Pan, J.; Huang, Y.; Su, M.; et al. Transcriptome Analyses Reveal FOXA1 Dysregulation in Mammary and Extramammary Paget’s Disease. Hum. Pathol. 2018, 77, 152–158. [Google Scholar] [CrossRef]
- Leonardi, G.C.; Falzone, L.; Salemi, R.; Zanghì, A.; Spandidos, D.A.; Mccubrey, J.A.; Candido, S.; Libra, M. Cutaneous Melanoma: From Pathogenesis to Therapy. Int. J. Oncol. 2018, 52, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Shain, A.H.; Bastian, B.C. From Melanocytes to Melanomas. Nat. Rev. Cancer 2016, 16, 345. [Google Scholar] [CrossRef]
- Brinck, U.; Ruschenburg, I.; Di Como, C.J.; Buschmann, N.; Betke, H.; Stachura, J.; Cordon-Cardo, C.; Korabiowska, M. Comparative Study of P63 and P53 Expression in Tissue Microarrays of Malignant Melanomas. Int. J. Mol. Med. 2002, 10, 707–711. [Google Scholar] [CrossRef]
- Reis-Filho, J.S.; Simpson, P.T.; Martins, A.; Preto, A.; Gärtner, F.; Schmitt, F.C. Distribution of P63, Cytokeratins 5/6 and Cytokeratin 14 in 51 Normal and 400 Neoplastic Human Tissue Samples Using TARP-4 Multi-Tumor Tissue Microarray. Virchows Arch. 2003, 443, 122–132. [Google Scholar] [PubMed] [Green Version]
- Bourne, T.D.; Bellizzi, A.M.; Stelow, E.B.; Loy, A.H.; Levine, P.A.; Wick, M.R.; Mills, S.E. P63 Expression in Olfactory Neuroblastoma and Other Small Cell Tumors of the Sinonasal Tract. Am. J. Clin. Pathol. 2008, 130, 213–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monti, P.; Ghiorzo, P.; Menichini, P.; Foggetti, G.; Queirolo, P.; Izzotti, A.; Fronza, G. TP63 Mutations are Frequent in Cutaneous Melanoma, Support UV Etiology, but Their Role in Melanomagenesis is Unclear. Oncol. Rep. 2017, 38, 1985–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, G.; Cipollone, R.; Lena, A.M.; Serra, V.; Zhou, H.; van Bokhoven, H.; Dötsch, V.; Merico, D.; Mantovani, R.; Terrinoni, A.; et al. Differential Altered Stability and Transcriptional Activity of ΔNp63 Mutants in Distinct Ectodermal Dysplasias. J. Cell Sci. 2011, 124, 2200–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, J.I.; Egevad, L.; Humphrey, P.A.; Montironi, R.; Amin, M.B.; Ulbright, T.M.; Grignon, D.; Trpkov, K.; Lopez-Beltran, A.; Zhou, M.; et al. Best Practices Recommendations in the Application of Immunohistochemistry in the Prostate: Report from the International Society of Urologic Pathology Consensus Conference. Am. J. Surg. Pathol. 2014, 38, e6–e19. [Google Scholar] [CrossRef]
- Amin, M.B.; Epstein, J.I.; Ulbright, T.M.; Humphrey, P.A.; Egevad, L.; Montironi, R.; Grignon, D.; Trpkov, K.; Lopez-Beltran, A.; Zhou, M.; et al. Best Practices Recommendations in the Application of Immunohistochemistry in Urologic Pathology: Report from the International Society of Urological Pathology Consensus Conference. Am. J. Surg. Pathol. 2014, 38, 1017–1022. [Google Scholar] [CrossRef]
- Travis, W.D.; Brambilla, E.; Noguchi, M.; Nicholson, A.G.; Geisinger, K.; Yatabe, Y.; Powell, C.A.; Beer, D.; Riely, G.; Garg, K.; et al. International Association for the Study of Lung Cancer/American Thoracic Society/European Respiratory Society: International Multidisciplinary Classification of Lung Adenocarcinoma—An Executive Summary. Proc. Am. Thorac. Soc. 2011, 6, 244–285. [Google Scholar] [CrossRef] [Green Version]
- Mehregan, A.H. The Origin of the Adnexal Tumors of the Skin: A Viewpoint. J. Cutan. Pathol. 1985, 12, 459–467. [Google Scholar] [CrossRef]
- Agrawal, S.; Jain, R.; Panchonia, A.; Kulkarni, C.; Mehar, R. “Troublesome Tumors” of the Skin: Spectrum of Skin Adnexal Tumors at a Tertiary Care Center in Malwa Region. Int. J. Med. Sci. Public Health 2018, 7, 714–719. [Google Scholar] [CrossRef]
- Fulton, E.H.; Kaley, J.R.; Gardner, J.M. Skin Adnexal Tumors in Plain Language a Practical Approach for the General Surgical Pathologist. Arch. Pathol. Lab. Med. 2019, 143, 832–851. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, H.S.; Ormsby, A.H.; Lee, M.W.; Zarbo, R.J.; Ma, C.K. The Diagnostic Utility of P63, CK5/6, CK 7, and CK 20 in Distinguishing Primary Cutaneous Adnexal Neoplasms from Metastatic Carcinomas. J. Cutan. Pathol. 2004, 31, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, H.S.; Salama, M.E.; Chitale, D.; Bansal, I.; Ma, C.K.; Raju, U.; Ormsby, A.; Lee, M.W. Primary Cutaneous Mucinous Carcinoma: Presence of Myoepithelial Cells as a Clue to the Cutaneous Origin. Am. J. Dermatopathol. 2004, 26, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Ivan, D.; Diwan, A.H.; Prieto, V.G. Expression of P63 in Primary Cutaneous Adnexal Neoplasms and Adenocarcinoma Metastatic to the Skin. J. Cutan. Pathol. 2005, 32, 94. [Google Scholar] [CrossRef]
- Ivan, D.; Nash, J.W.; Prieto, V.G.; Calonje, E.; Lyle, S.; Diwan, A.H.; Lazar, A.J.F. Use of P63 Expression in Distinguishing Primary and Metastatic Cutaneous Adnexal Neoplasms from Metastatic Adenocarcinoma to Skin. J. Cutan. Pathol. 2007, 34, 474–480. [Google Scholar] [CrossRef]
- Valencia-Guerrero, A.; Dresser, K.; Cornejo, K.M. Utility of Immunohistochemistry in Distinguishing Primary Adnexal Carcinoma from Metastatic Breast Carcinoma to Skin and Squamous Cell Carcinoma. Am. J. Dermatopathol. 2018, 40, 389–396. [Google Scholar] [CrossRef]
- Panse, G.; Cowper, S.E.; Leffell, D.J.; Pulitzer, M.; Ko, C.J. Well-Differentiated Neuroendocrine Tumors in Skin: Terminology and Diagnostic Utility of Cytokeratin 5/6 and P63. J. Cutan. Pathol. 2017, 44, 557–562. [Google Scholar] [CrossRef]
- Chapman, L.W.; Yu, S.S.; Arron, S.T. Atypical Fibroxanthoma. Semin. Cutan. Med. Surg. 2019, 38, E65–E66. [Google Scholar] [CrossRef]
- Koch, M.; Freundl, A.J.; Agaimy, A.; Kiesewetter, F.; Künzel, J.; Cicha, I.; Alexiou, C. Atypical Fibroxanthoma-Histological Diagnosis, Immunohistochemical Markers and Concepts of Therapy. Anticancer Res. 2015, 35, 5717–5735. [Google Scholar]
- Morgan, M.B.; Purohit, C.; Anglin, T.R. Immunohistochemical Distinction of Cutaneous Spindle Cell Carcinoma. Am. J. Dermatopathol. 2008, 30, 228–232. [Google Scholar] [CrossRef]
- Buonaccorsi, J.N.; Plaza, J.A. Role of CD10, Wide-Spectrum Keratin, P63, and Podoplanin in the Distinction of Epithelioid and Spindle Cell Tumors of the Skin: An Immunohistochemical Study of 81 Cases. Am. J. Dermatopathol. 2012, 34, 404–411. [Google Scholar] [CrossRef]
- Henderson, S.A.; Torres-Cabala, C.A.; Curry, J.L.; Bassett, R.L.; Ivan, D.; Prieto, V.G.; Tetzlaff, M.T. P40 Is More Specific than P63 for the Distinction of Atypical Fibroxanthoma from Other Cutaneous Spindle Cell Malignancies. Am. J. Surg. Pathol. 2014, 38, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Alomari, A.K.; Glusac, E.J.; McNiff, J.M. P40 Is a More Specific Marker than P63 for Cutaneous Poorly Differentiated Squamous Cell Carcinoma. J. Cutan. Pathol. 2014, 41, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.C.; Stang, A.; DeCaprio, J.A.; Cerroni, L.; Lebbé, C.; Veness, M.; Nghiem, P. Merkel Cell Carcinoma. Nat. Rev. Dis. Primer 2017, 3, 17077. [Google Scholar] [CrossRef] [PubMed]
- Asioli, S.; Righi, A.; Volante, M.; Eusebi, V.; Bussolati, G. P63 Expression as a New Prognostic Marker in Mrkel Cell Carcinoma. Cancer 2007, 110, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Asioli, S.; Righi, A.; De Biase, D.; Morandi, L.; Caliendo, V.; Picciotto, F.; MacRipò, G.; Maletta, F.; Di Cantogno, L.V.; Chiusa, L.; et al. Expression of P63 Is the Sole Independent Marker of Aggressiveness in Localised (Stage I–II) Merkel Cell Carcinomas. Mod. Pathol. 2011, 24, 1451. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.J.; Pincus, L.B.; Yu, S.S.; Oh, D.H.; Wilson, A.R.; McCalmont, T.H. Immunohistochemical Prognostication of Merkel Cell Carcinoma: P63 Expression but Not Polyomavirus Status Correlates with Outcome. J. Cutan. Pathol. 2012, 39, 911–917. [Google Scholar] [CrossRef]
- Stetsenko, G.Y.; Malekirad, J.; Paulson, K.G.; Iyer, J.G.; Thibodeau, R.M.; Nagase, K.; Schmidt, M.; Storer, B.E.; Argenyi, Z.B.; Nghiem, P. P63 Expression in Merkel Cell Carcinoma Predicts Poorer Survival yet May Have Limited Clinical Utility. Am. J. Clin. Pathol. 2013, 140, 838–844. [Google Scholar] [CrossRef] [Green Version]
- Fleming, K.E.; Ly, T.Y.; Pasternak, S.; Godlewski, M.; Doucette, S.; Walsh, N.M. Support for P63 Expression as an Adverse Prognostic Marker in Merkel Cell Carcinoma: Report on a Canadian Cohort. Hum. Pathol. 2014, 45, 952–960. [Google Scholar] [CrossRef]
- Lim, C.S.; Whalley, D.; Haydu, L.E.; Murali, R.; Tippett, J.; Thompson, J.F.; Hruby, G.; Scolyer, R.A. Increasing Tumor Thickness Is Associated with Recurrence and Poorer Survival in Patients with Merkel Cell Carcinoma. Ann. Surg. Oncol. 2012, 19, 3325–3334. [Google Scholar] [CrossRef]
- Dabner, M.; McClure, R.J.; Harvey, N.T.; Budgeon, C.A.; Beer, T.W.; Amanuel, B.; Wood, B.A. Merkel Cell Polyomavirus and P63 Status in Merkel Cell Carcinoma by Immunohistochemistry: Merkel Cell Polyomavirus Positivity Is Inversely Correlated with Sun Damage, but Neither Is Correlated with Outcome. Pathology 2014, 46, 205–210. [Google Scholar] [CrossRef]
- Senoo, M.; Tsuchiya, I.; Matsumura, Y.; Mori, T.; Saito, Y.; Kato, H.; Okamoto, T.; Habu, S. Transcriptional Dysregulation of the P73L/P63/P51/P40/KET Gene in Human Squamous Cell Carcinomas: Expression of ΔNp73L, a Novel Dominant-Negative Isoform, and Loss of Expression of the Potential Tumour Suppressor P51. Br. J. Cancer 2001, 84, 1235. [Google Scholar] [CrossRef] [PubMed]
- Geddert, H.; Kiel, S.; Heep, H.J.; Gabbert, H.E.; Sarbia, M. The Role of P63 and ΔNp63 (P40) Protein Expression and Gene Amplification in Esophageal Carcinogenesis. Hum. Pathol. 2003, 34, 850–856. [Google Scholar] [CrossRef]
- Bishop, J.A.; Teruya-Feldstein, J.; Westra, W.H.; Pelosi, G.; Travis, W.D.; Rekhtman, N. P40 (ΔNp63) Is Superior to P63 for the Diagnosis of Pulmonary Squamous Cell Carcinoma. Mod. Pathol. 2012, 25, 405. [Google Scholar] [CrossRef] [PubMed]
- Nekulova, M.; Holcakova, J.; Nenutil, R.; Stratmann, R.; Bouchalova, P.; Müller, P.; Mouková, L.; Coates, P.J.; Vojtesek, B. Characterization of Specific P63 and P63-N-Terminal Isoform Antibodies and Their Application for Immunohistochemistry. Virchows Arch. 2013, 463, 415–425. [Google Scholar] [CrossRef]
- Lee, J.J.; Mochel, M.C.; Piris, A.; Boussahmain, C.; Mahalingam, M.; Hoang, M.P. P40 Exhibits Better Specificity than P63 in Distinguishing Primary Skin Adnexal Carcinomas from Cutaneous Metastases. Hum. Pathol. 2014, 45, 1078–1083. [Google Scholar] [CrossRef]
- Andruszkow, J.H.K.; Oll, M.; Förster, S.; Knüchel, R.; Jäkel, J. P40 in Conjunction with CK20 and E-Cadherin Distinguishes Primary Adnexal Neoplasms of the Skin. Appl. Immunohistochem. Mol. Morphol. 2016, 24, 414–421. [Google Scholar] [CrossRef]
- Jain, D.; Mathur, S.R.; Sharma, M.C.; Iyer, V.K. Cytomorphology of Sebaceous Carcinoma with Analysis of P40 Antibody Expression. Diagn. Cytopathol. 2015, 43, 456–461. [Google Scholar] [CrossRef]
- Kriegsmann, K.; Cremer, M.; Zgorzelski, C.; Harms, A.; Muley, T.; Winter, H.; Kazdal, D.; Warth, A.; Kriegsmann, M. Agreement of CK5/6, P40, and P63 Immunoreactivity in Non-Small Cell Lung Cancer. Pathology 2019, 51, 240–245. [Google Scholar] [CrossRef]
- Tacha, D.; Bremer, R.; Haas, T.; Qi, W. An Immunohistochemical Analysis of a Newly Developed, Mouse Monoclonal P40 (BC28) Antibody in Lung, Bladder, Skin, Breast, Prostate, and Head and Neck Cancers. Arch. Pathol. Lab. Med. 2014, 138, 1358–1364. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smirnov, A.; Anemona, L.; Novelli, F.; Piro, C.M.; Annicchiarico-Petruzzelli, M.; Melino, G.; Candi, E. p63 Is a Promising Marker in the Diagnosis of Unusual Skin Cancer. Int. J. Mol. Sci. 2019, 20, 5781. https://doi.org/10.3390/ijms20225781
Smirnov A, Anemona L, Novelli F, Piro CM, Annicchiarico-Petruzzelli M, Melino G, Candi E. p63 Is a Promising Marker in the Diagnosis of Unusual Skin Cancer. International Journal of Molecular Sciences. 2019; 20(22):5781. https://doi.org/10.3390/ijms20225781
Chicago/Turabian StyleSmirnov, Artem, Lucia Anemona, Flavia Novelli, Cristina M. Piro, Margherita Annicchiarico-Petruzzelli, Gerry Melino, and Eleonora Candi. 2019. "p63 Is a Promising Marker in the Diagnosis of Unusual Skin Cancer" International Journal of Molecular Sciences 20, no. 22: 5781. https://doi.org/10.3390/ijms20225781
APA StyleSmirnov, A., Anemona, L., Novelli, F., Piro, C. M., Annicchiarico-Petruzzelli, M., Melino, G., & Candi, E. (2019). p63 Is a Promising Marker in the Diagnosis of Unusual Skin Cancer. International Journal of Molecular Sciences, 20(22), 5781. https://doi.org/10.3390/ijms20225781