Puf6 and Loc1 Are the Dedicated Chaperones of Ribosomal Protein Rpl43 in Saccharomyces cerevisiae


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Reagents
2.2. The Analysis of Protein Stability and Western Blotting
2.3. Immunoprecipitation
2.4. Polysome Profile Analysis
2.5. Northern Blotting
2.6. In Vitro Interaction Experiments
3. Results
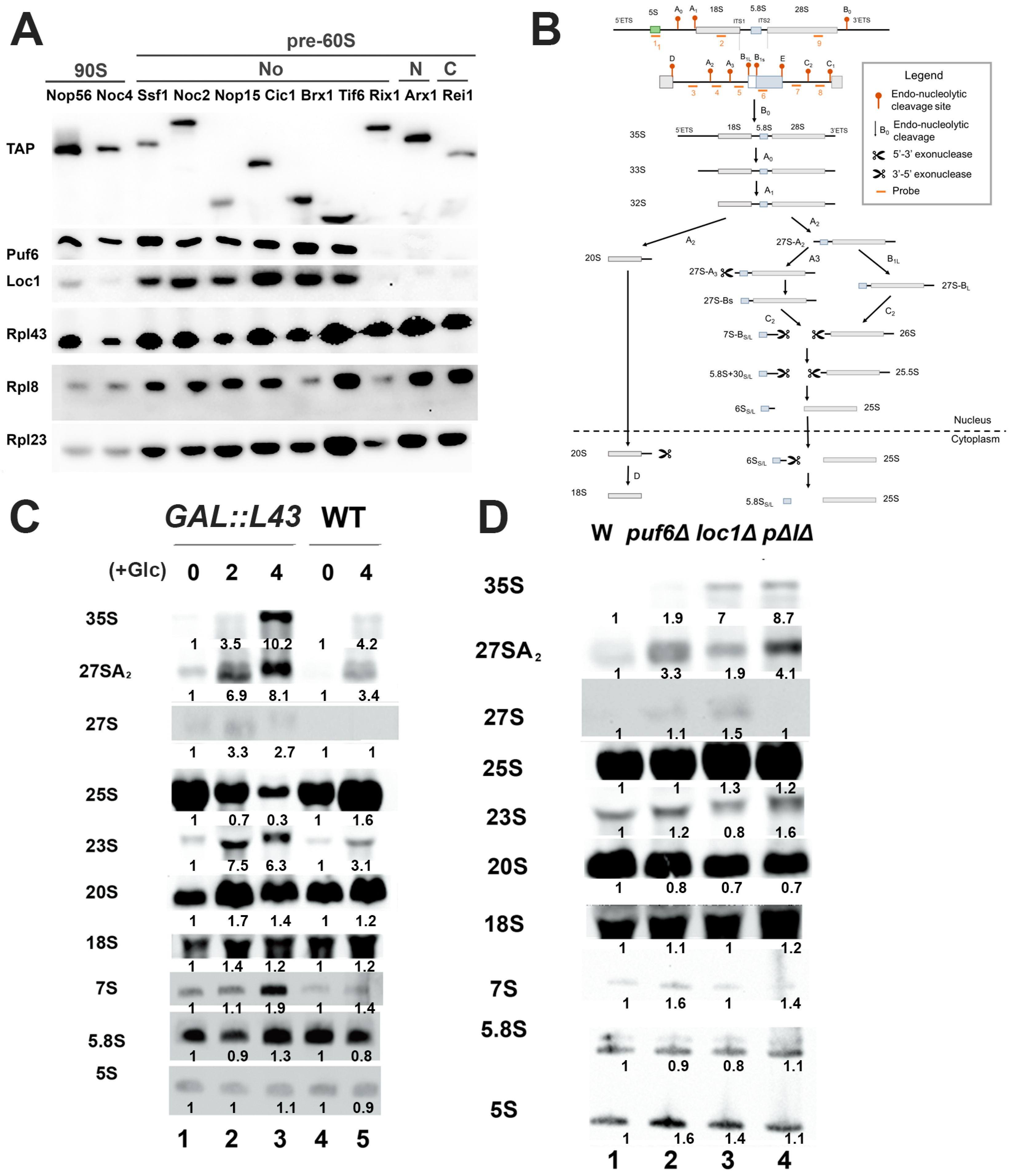
3.1. Mutations of Puf6, Loc1, and Rpl43 Show Similar rRNA Processing Defects
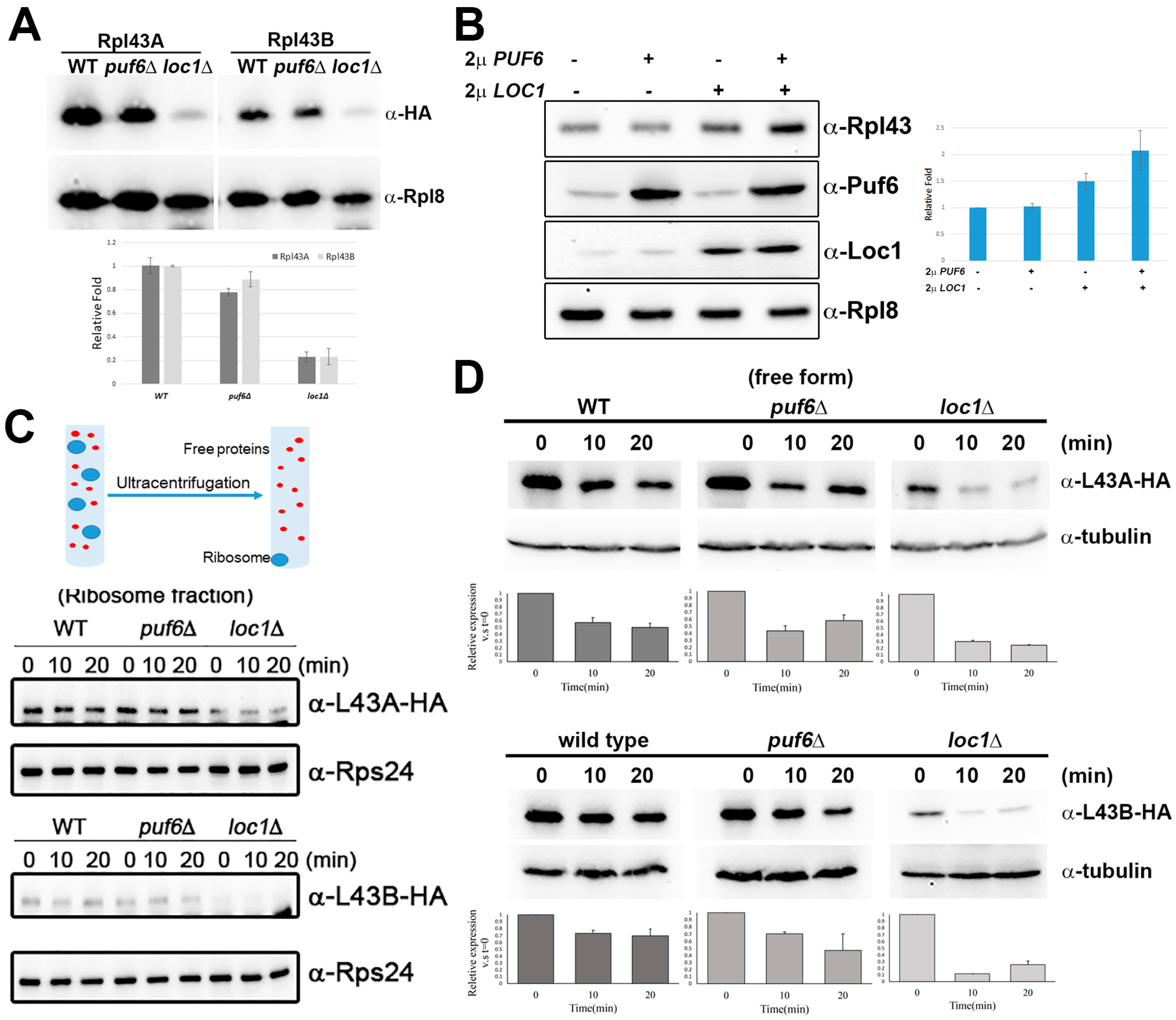
3.2. Puf6 and Loc1 Are Important for the Protein Stability of Rpl43
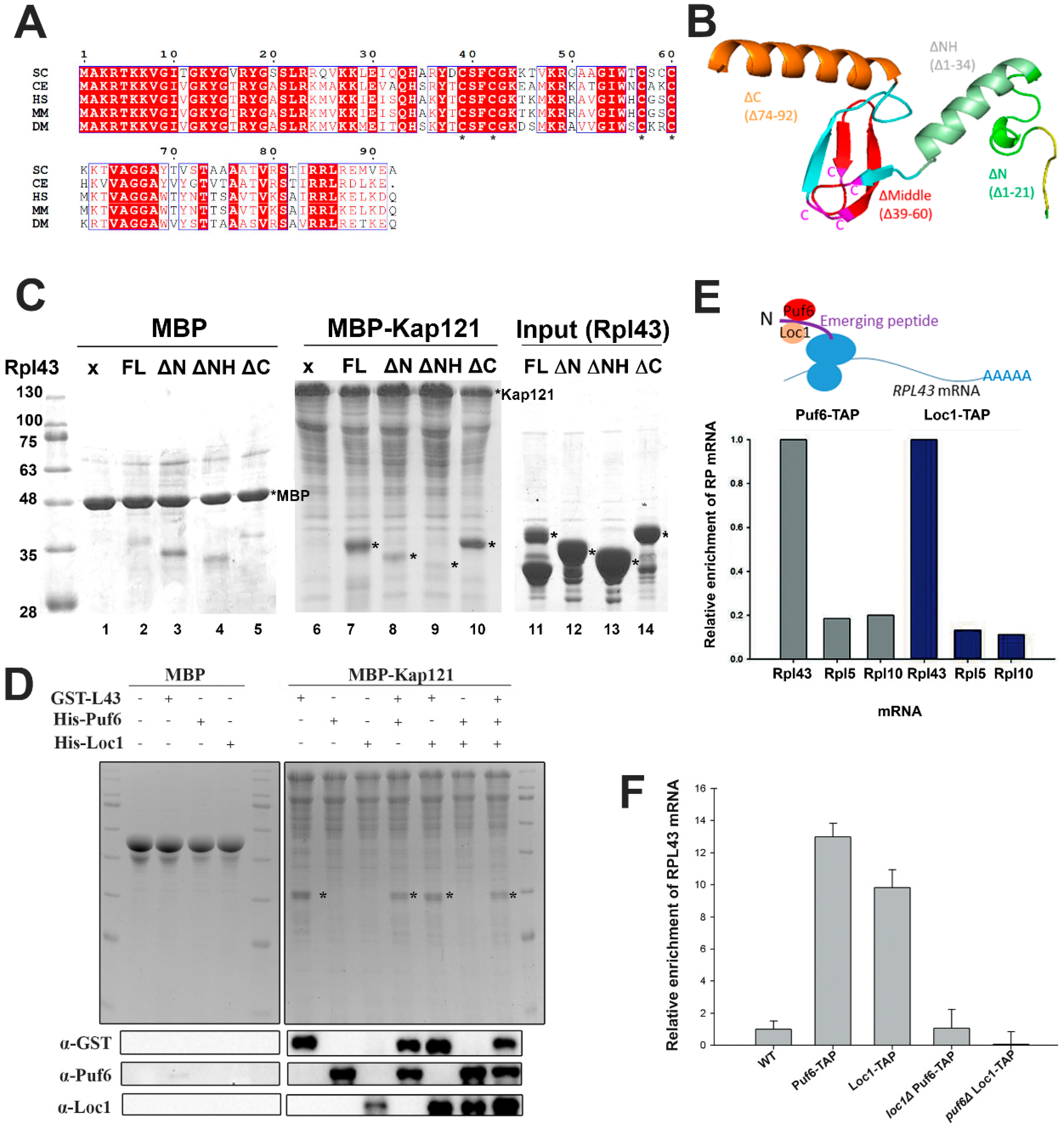
3.3. Puf6 and Loc1 Can Associate with Nascent Rpl43 but Are Not Required for Kap Interaction or Release
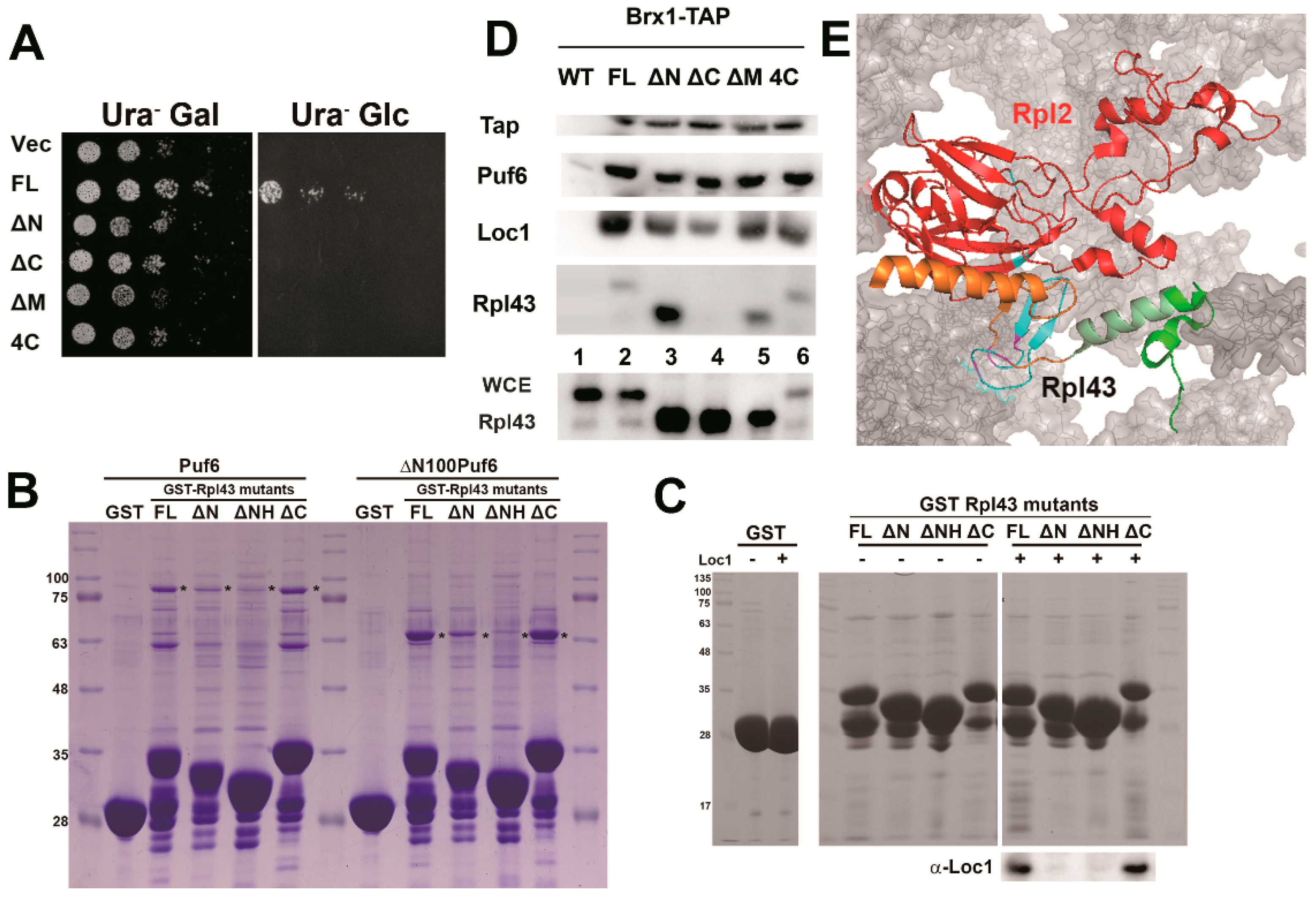
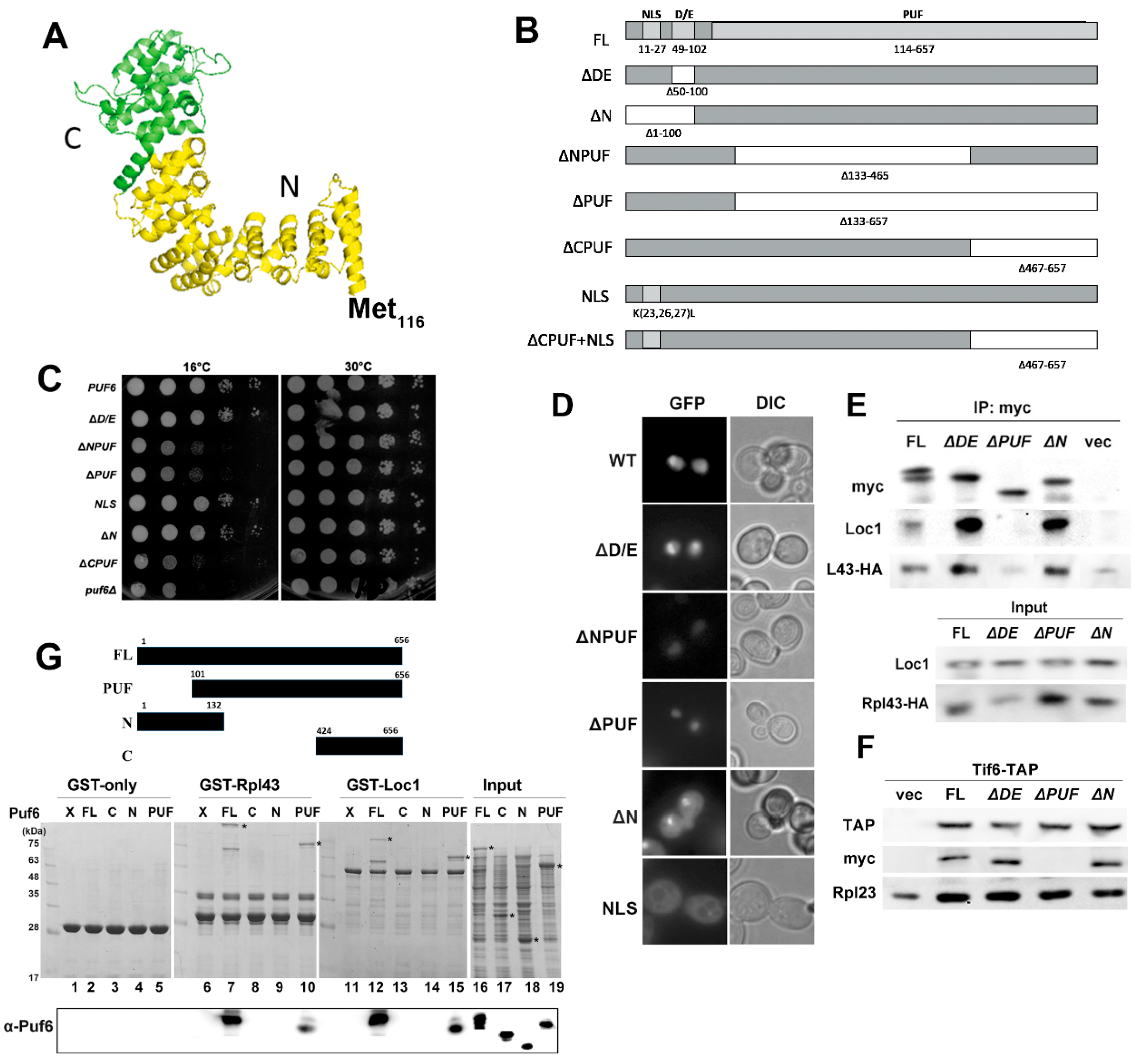
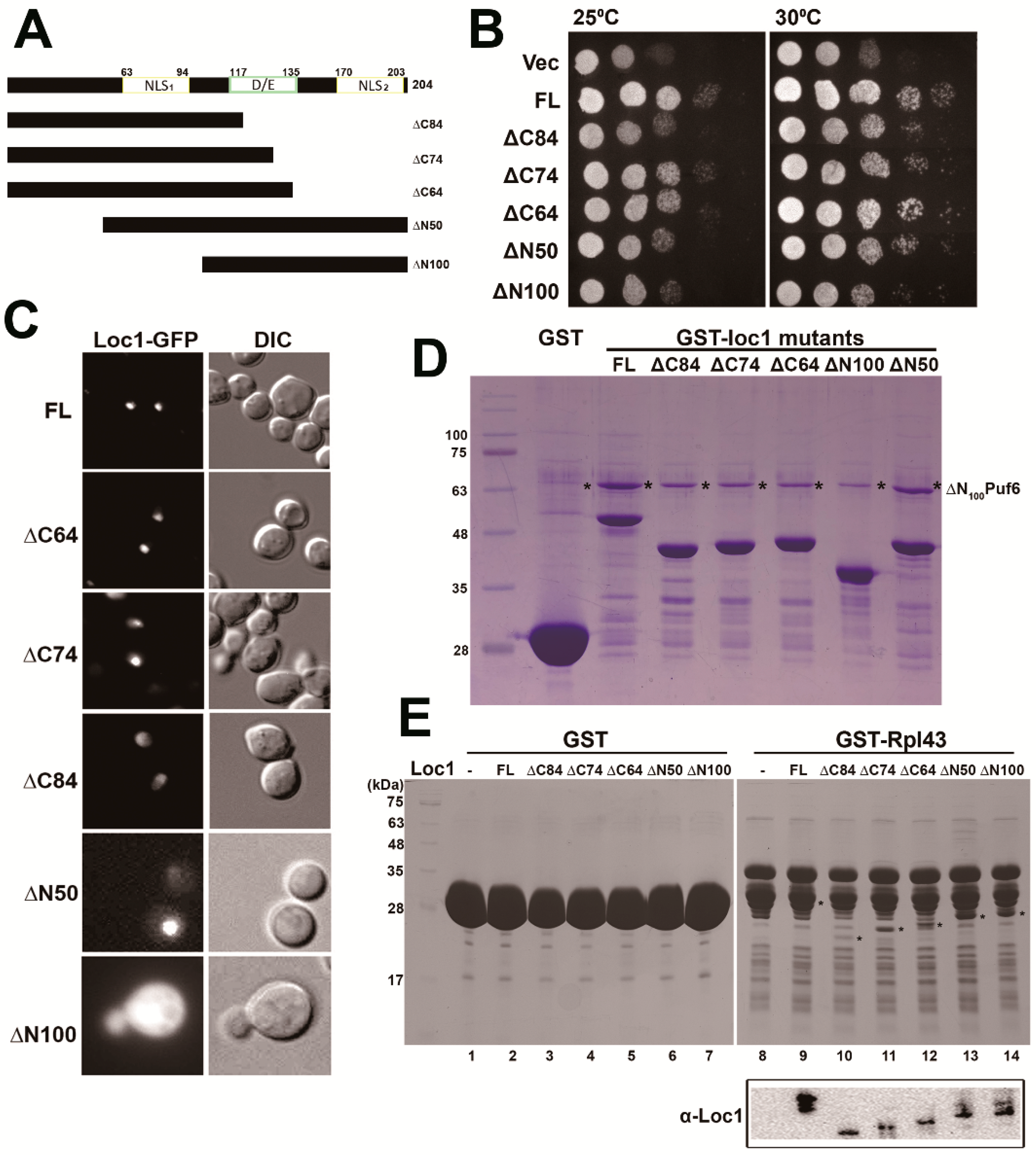
3.4. Determination of the Potential Interaction Sites among the Three Proteins
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Warner, J.R. The economics of ribosome biosynthesis in yeast. Trends. Biochem. Sci. 1999, 24, 437–440. [Google Scholar] [CrossRef]
- Dufner, A.; Thomas, G. Ribosomal S6 kinase signaling and the control of translation. Exp. Cell Res. 1999, 253, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M. Regulation of ribosome biosynthesis in Escherichia coli and Saccharomyces cerevisiae: Diversity and common principles. J. Bacteriol. 1999, 181, 6857–6864. [Google Scholar] [PubMed]
- Leary, D.J.; Huang, S. Regulation of ribosome biogenesis within the nucleolus. Febs Lett. 2001, 509, 145–150. [Google Scholar] [CrossRef]
- Tschochner, H.; Hurt, E. Pre-ribosomes on the road from the nucleolus to the cytoplasm. Trends. Cell. Biol. 2003, 13, 255–263. [Google Scholar] [CrossRef]
- Venema, J.; Tollervey, D. Ribosome synthesis in Saccharomyces cerevisiae. Annu. Rev. Genet. 1999, 33, 261–311. [Google Scholar] [CrossRef] [PubMed]
- Woolford, J.L., Jr.; Baserga, S.J. Ribosome biogenesis in the yeast Saccharomyces cerevisiae. Genetics 2013, 195, 643–681. [Google Scholar] [CrossRef]
- Panse, V.G.; Johnson, A.W. Maturation of eukaryotic ribosomes: Acquisition of functionality. Trends Biochem. Sci. 2010, 35, 260–266. [Google Scholar] [CrossRef]
- Gerhardy, S.; Menet, A.M.; Pena, C.; Petkowski, J.J.; Panse, V.G. Assembly and nuclear export of pre-ribosomal particles in budding yeast. Chromosoma 2014, 123, 327–344. [Google Scholar] [CrossRef]
- Zemp, I.; Kutay, U. Nuclear export and cytoplasmic maturation of ribosomal subunits. Febs Lett. 2007, 581, 2783–2793. [Google Scholar] [CrossRef]
- Kraushar, M.L.; Popovitchenko, T.; Volk, N.L.; Rasin, M.R. The frontier of RNA metamorphosis and ribosome signature in neocortical development. Int. J. Dev. Neurosci. 2016, 55, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Wool, I.G. The structure and function of eukaryotic ribosomes. Annu. Rev. Biochem. 1979, 48, 719–754. [Google Scholar] [CrossRef] [PubMed]
- Velculescu, V.E.; Zhang, L.; Zhou, W.; Vogelstein, J.; Basrai, M.A.; Bassett, D.E., Jr.; Hieter, P.; Vogelstein, B.; Kinzler, K.W. Characterization of the yeast transcriptome. Cell 1997, 88, 243–251. [Google Scholar] [CrossRef]
- Pillet, B.; Mitterer, V.; Kressler, D.; Pertschy, B. Hold on to your friends: Dedicated chaperones of ribosomal proteins: Dedicated chaperones mediate the safe transfer of ribosomal proteins to their site of pre-ribosome incorporation. Bioessays: News Rev. Mol. Cell. Dev. Biol. 2017, 39, 1–12. [Google Scholar] [CrossRef]
- Tsay, Y.F.; Thompson, J.R.; Rotenberg, M.O.; Larkin, J.C.; Woolford, J.L. Ribosomal-Protein Synthesis Is Not Regulated at the Translational Level in Saccharomyces-Cerevisiae - Balanced Accumulation of Ribosomal Protein-L16 and Protein-Rp59 Is Mediated by Turnover of Excess Protein. Gene. Dev. 1988, 2, 664–676. [Google Scholar] [CrossRef]
- Eisinger, D.P.; Dick, F.A.; Denke, E.; Trumpower, B.L. SQT1, which encodes an essential WD domain protein of Saccharomyces cerevisiae, suppresses dominant-negative mutations of the ribosomal protein gene QSR1. Mol. Cell. Biol. 1997, 17, 5146–5155. [Google Scholar] [CrossRef]
- West, M.; Hedges, J.B.; Chen, A.; Johnson, A.W. Defining the order in which Nmd3p and Rpl10p load onto nascent 60S ribosomal subunits. Mol. Cell. Biol. 2005, 25, 3802–3813. [Google Scholar] [CrossRef]
- Pillet, B.; Garcia-Gomez, J.J.; Pausch, P.; Falquet, L.; Bange, G.; de la Cruz, J.; Kressler, D. The Dedicated Chaperone Acl4 Escorts Ribosomal Protein Rpl4 to Its Nuclear Pre-60S Assembly Site. PLoS Genet. 2015, 11, e1005565. [Google Scholar] [CrossRef]
- Stelter, P.; Huber, F.M.; Kunze, R.; Flemming, D.; Hoelz, A.; Hurt, E. Coordinated Ribosomal L4 Protein Assembly into the Pre-Ribosome Is Regulated by Its Eukaryote-Specific Extension. Mol. Cell 2015, 58, 854–862. [Google Scholar] [CrossRef]
- Holzer, S.; Ban, N.; Klinge, S. Crystal Structure of the Yeast Ribosomal Protein rpS3 in Complex with Its Chaperone Yar1. J. Mol. Biol. 2013. [Google Scholar] [CrossRef]
- Koch, B.; Mitterer, V.; Niederhauser, J.; Stanborough, T.; Murat, G.; Rechberger, G.; Bergler, H.; Kressler, D.; Pertschy, B. Yar1 protects the ribosomal protein Rps3 from aggregation. J. Biol. Chem. 2012, 287, 21806–21815. [Google Scholar] [CrossRef] [PubMed]
- Iouk, T.L.; Aitchison, J.D.; Maguire, S.; Wozniak, R.W. Rrb1p, a yeast nuclear WD-repeat protein involved in the regulation of ribosome biosynthesis. Mol. Cell. Biol. 2001, 21, 1260–1271. [Google Scholar] [CrossRef] [PubMed]
- Schaper, S.; Fromont-Racine, M.; Linder, P.; de la Cruz, J.; Namane, A.; Yaniv, M. A yeast homolog of chromatin assembly factor 1 is involved in early ribosome assembly. Curr. Biol. 2001, 11, 1885–1890. [Google Scholar] [CrossRef]
- Schutz, S.; Fischer, U.; Altvater, M.; Nerurkar, P.; Pena, C.; Gerber, M.; Chang, Y.; Caesar, S.; Schubert, O.T.; Schlenstedt, G.; et al. A RanGTP-independent mechanism allows ribosomal protein nuclear import for ribosome assembly. eLife 2014, 3, e03473. [Google Scholar] [CrossRef] [PubMed]
- Schutz, S.; Michel, E.; Damberger, F.F.; Oplova, M.; Pena, C.; Leitner, A.; Aebersold, R.; Allain, F.H.T.; Panse, V.G. Molecular basis for disassembly of an importin: Ribosomal protein complex by the escortin Tsr2. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Bange, G.; Murat, G.; Sinning, I.; Hurt, E.; Kressler, D. New twist to nuclear import: When two travel together. Commun. Integr. Biol. 2013, 6, e24792. [Google Scholar] [CrossRef]
- Calvino, F.R.; Kharde, S.; Ori, A.; Hendricks, A.; Wild, K.; Kressler, D.; Bange, G.; Hurt, E.; Beck, M.; Sinning, I. Symportin 1 chaperones 5S RNP assembly during ribosome biogenesis by occupying an essential rRNA-binding site. Nat. Commun. 2015, 6, 6510. [Google Scholar] [CrossRef]
- Kressler, D.; Bange, G.; Ogawa, Y.; Stjepanovic, G.; Bradatsch, B.; Pratte, D.; Amlacher, S.; Strauss, D.; Yoneda, Y.; Katahira, J.; et al. Synchronizing nuclear import of ribosomal proteins with ribosome assembly. Science 2012, 338, 666–671. [Google Scholar] [CrossRef]
- Ting, Y.H.; Lu, T.J.; Johnson, A.W.; Shie, J.T.; Chen, B.R.; Kumar, S.S.; Lo, K.Y. Bcp1 is the Nuclear Chaperone of the 60S ribosomal protein Rpl23 in Saccharomyces cerevisiae. J. Biol. Chem. 2017, 292, 585–596. [Google Scholar] [CrossRef]
- Rossler, I.; Embacher, J.; Pillet, B.; Murat, G.; Liesinger, L.; Hafner, J.; Unterluggauer, J.J.; Birner-Gruenberger, R.; Kressler, D.; Pertschy, B. Tsr4 and Nap1, two novel members of the ribosomal protein chaperOME. Nucleic Acids Res. 2019, 47, 6984–7002. [Google Scholar] [CrossRef]
- Black, J.J.; Musalgaonkar, S.; Johnson, A.W. Tsr4 Is a Cytoplasmic Chaperone for the Ribosomal Protein Rps2 in Saccharomyces cerevisiae. Mol. Cell. Biol. 2019, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pausch, P.; Singh, U.; Ahmed, Y.L.; Pillet, B.; Murat, G.; Altegoer, F.; Stier, G.; Thoms, M.; Hurt, E.; Sinning, I.; et al. Co-translational capturing of nascent ribosomal proteins by their dedicated chaperones. Nat. Commun. 2015, 6, 7494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pena, C.; Hurt, E.; Panse, V.G. Eukaryotic ribosome assembly, transport and quality control. Nat. Struct. Mol. Biol. 2017, 24, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Jenner, L.; Melnikov, S.; Garreau de Loubresse, N.; Ben-Shem, A.; Iskakova, M.; Urzhumtsev, A.; Meskauskas, A.; Dinman, J.; Yusupova, G.; Yusupov, M. Crystal structure of the 80S yeast ribosome. Curr. Opin. Struct. Biol. 2012, 22, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Ban, N.; Nissen, P.; Hansen, J.; Moore, P.B.; Steitz, T.A. The complete atomic structure of the large ribosomal subunit at 2.4 A resolution. Science 2000, 289, 905–920. [Google Scholar] [CrossRef]
- Poll, G.; Braun, T.; Jakovljevic, J.; Neueder, A.; Jakob, S.; Woolford, J.L., Jr.; Tschochner, H.; Milkereit, P. rRNA maturation in yeast cells depleted of large ribosomal subunit proteins. PLoS ONE 2009, 4, e8249. [Google Scholar] [CrossRef] [Green Version]
- Long, R.M.; Gu, W.; Meng, X.; Gonsalvez, G.; Singer, R.H.; Chartrand, P. An exclusively nuclear RNA-binding protein affects asymmetric localization of ASH1 mRNA and Ash1p in yeast. J. Cell. Biol. 2001, 153, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.T.; Ting, Y.H.; Liang, K.J.; Lo, K.Y. The Roles of Puf6 and Loc1 in 60S Biogenesis Are Interdependent, and Both Are Required for Efficient Accommodation of Rpl43. J. Biol. Chem. 2016, 291, 19312–19323. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Lee, I.; Moradi, E.; Hung, N.J.; Johnson, A.W.; Marcotte, E.M. Rational extension of the ribosome biogenesis pathway using network-guided genetics. PLoS Biol. 2009, 7, e1000213. [Google Scholar] [CrossRef] [Green Version]
- Fromont-Racine, M.; Senger, B.; Saveanu, C.; Fasiolo, F. Ribosome assembly in eukaryotes. Gene 2003, 313, 17–42. [Google Scholar] [CrossRef]
- Konikkat, S.; Woolford, J.L., Jr. Principles of 60S ribosomal subunit assembly emerging from recent studies in yeast. Biochem. J. 2017, 474, 195–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, K.Y.; Li, Z.; Bussiere, C.; Bresson, S.; Marcotte, E.M.; Johnson, A.W. Defining the pathway of cytoplasmic maturation of the 60S ribosomal subunit. Mol. Cell 2010, 39, 196–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamalinda, M.; Ohmayer, U.; Jakovljevic, J.; Kumcuoglu, B.; Woolford, J.; Mbom, B.; Lin, L.; Woolford, J.L., Jr. A hierarchical model for assembly of eukaryotic 60S ribosomal subunit domains. Genes Dev. 2014, 28, 198–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohmayer, U.; Gamalinda, M.; Sauert, M.; Ossowski, J.; Poll, G.; Linnemann, J.; Hierlmeier, T.; Perez-Fernandez, J.; Kumcuoglu, B.; Leger-Silvestre, I.; et al. Studies on the assembly characteristics of large subunit ribosomal proteins in S. cerevisae. PLoS ONE 2013, 8, e68412. [Google Scholar] [CrossRef]
- Waskiewicz-Staniorowska, B.; Skala, J.; Jasinski, M.; Grenson, M.; Goffeau, A.; Ulaszewski, S. Functional analysis of three adjacent open reading frames from the right arm of yeast chromosome XVI. Yeast 1998, 14, 1027–1039. [Google Scholar] [CrossRef]
- Leidig, C.; Thoms, M.; Holdermann, I.; Bradatsch, B.; Berninghausen, O.; Bange, G.; Sinning, I.; Hurt, E.; Beckmann, R. 60S ribosome biogenesis requires rotation of the 5S ribonucleoprotein particle. Nat. Commun. 2014, 5, 3491. [Google Scholar] [CrossRef]
- Chook, Y.M.; Blobel, G. Karyopherins and nuclear import. Curr. Opin. Struct. Biol. 2001, 11, 703–715. [Google Scholar] [CrossRef]
- Rout, M.P.; Blobel, G.; Aitchison, J.D. A distinct nuclear import pathway used by ribosomal proteins. Cell 1997, 89, 715–725. [Google Scholar] [CrossRef] [Green Version]
- Edwards, T.A.; Pyle, S.E.; Wharton, R.P.; Aggarwal, A.K. Structure of Pumilio reveals similarity between RNA and peptide binding motifs. Cell 2001, 105, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Gavis, E.R. Over the rainbow to translational control. Nat. Struct. Biol. 2001, 8, 387–389. [Google Scholar] [CrossRef]
- Wang, X.; Zamore, P.D.; Hall, T.M. Crystal structure of a Pumilio homology domain. Mol. Cell. 2001, 7, 855–865. [Google Scholar] [CrossRef]
- Qiu, C.; McCann, K.L.; Wine, R.N.; Baserga, S.J.; Hall, T.M. A divergent Pumilio repeat protein family for pre-rRNA processing and mRNA localization. Proc. Natl. Acad. Sci. USA 2014, 111, 18554–18559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, A.; Mills, R.E.; Lange, C.J.; Stewart, M.; Devine, S.E.; Corbett, A.H. Classical nuclear localization signals: Definition, function, and interaction with importin alpha. J. Biol. Chem. 2007, 282, 5101–5105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraushar, M.L.; Thompson, K.; Wijeratne, H.R.; Viljetic, B.; Sakers, K.; Marson, J.W.; Kontoyiannis, D.L.; Buyske, S.; Hart, R.P.; Rasin, M.R. Temporally defined neocortical translation and polysome assembly are determined by the RNA-binding protein Hu antigen R. Proc. Natl. Acad. Sci. USA 2014, 111, E3815–E3824. [Google Scholar] [CrossRef] [Green Version]
- Kraushar, M.L.; Viljetic, B.; Wijeratne, H.R.; Thompson, K.; Jiao, X.; Pike, J.W.; Medvedeva, V.; Groszer, M.; Kiledjian, M.; Hart, R.P.; et al. Thalamic WNT3 Secretion Spatiotemporally Regulates the Neocortical Ribosome Signature and mRNA Translation to Specify Neocortical Cell Subtypes. J. Neurosci. 2015, 35, 10911–10926. [Google Scholar] [CrossRef] [Green Version]
- Andrade, J.M.; dos Santos, R.F.; Chelysheva, I.; Ignatova, Z.; Arraiano, C.M. The RNA-binding protein Hfq is important for ribosome biogenesis and affects translation fidelity. Embo J. 2018, 37. [Google Scholar] [CrossRef]
- Fischer, A.D.; Olivas, W.M. Multiple Puf proteins regulate the stability of ribosome biogenesis transcripts. RNA Biol. 2018, 15, 1228–1243. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| BY4741 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | |
| KLY67 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 puf6Δ::KanMX | [38] |
| KLY218 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 loc1Δ::KanMX | [38] |
| KLY312 | his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 puf6∆::KanMX loc1∆::KanMX | [38] |
| KLY134 | MATa PUF6-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| KLY317 | MATa ARX1-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | [38] |
| KLY415 | MATa loc1Δ::KanMX PUF6-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| KLY471 | MATa BRX1-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | [38] |
| KLY475 | MATa LOC1-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | [38] |
| KLY583 | MATa TIF6-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | [38] |
| KLY596 | MATa RIX1-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | [38] |
| KLY598 | MATa SSF1-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | [38] |
| KLY628 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 rpl43a∆::KanMX rpl43b∆::CloNAT GAL::RPL43B-HA (PKL381 CEN HIS3) | [38] |
| KLY672 | MATa REI1-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| KLY816 | MATa NOC2-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| KLY822 | MATa CIC1-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| KLY842 | MATa puf6Δ::KanMX LOC-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| KLY875 | MATa NOP15-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| KLY876 | MATa NOP56-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| KLY877 | MATa NOC4-TAP::HIS3MX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| Plasmid | Gene | Relevant Marker | Source |
|---|---|---|---|
| PKL52 | PUF6-GFP | CEN LEU2 | This study |
| PKL53 | puf6ΔDE-GFP | CEN LEU2 | This study |
| PKL54 | puf6ΔPUF-GFP | CEN LEU2 | This study |
| PKL55 | puf6ΔNPUF-GFP | CEN LEU2 | [38] |
| PKL56 | pET28a-PUF6 | Kan | [38] |
| PKL85 | PUF6-myc | CEN LEU2 | [38] |
| PKL86 | puf6ΔD/E-myc | CEN LEU2 | This study |
| PKL87 | puf6ΔPUF-myc | CEN LEU2 | This study |
| PKL88 | puf6ΔNPUF -myc | CEN LEU2 | This study |
| PKL188 | puf6ΔN-GFP | CEN LEU2 | This study |
| PKL189 | puf6ΔN-myc | CEN LEU2 | This study |
| PKL198 | puf6(NLS)-myc | CEN LEU2 | This study |
| PKL199 | puf6(NLS)-GFP | CEN LEU2 | This study |
| PKL288 | pET28a-LOC1 | Kan | [38] |
| PKL306 | puf6ΔCPUF-myc | CEN LEU2 | This study |
| PKL333 | PUF6 | 2μ URA3 | This study |
| PKL337 | LOC1-GFP | CEN LEU2 | [38] |
| PKL349 | RPL43A-HA | CEN URA3 | This study |
| PKL350 | RPL43B-HA | CEN URA3 | [38] |
| PKL400 | GST-LOC1 | CEN LEU2 | This study |
| PKL449 | loc1ΔC84-GFP | CEN LEU2 | This study |
| PKL450 | loc1ΔC74-GFP | CEN LEU2 | This study |
| PKL451 | loc1ΔC64-GFP | CEN LEU2 | This study |
| PKL474 | GST-RPL43 | Amp | [38] |
| PKL512 | pET28a-puf6(PUF) | Kan | This study |
| PKL518 | loc1ΔN100-GFP | CEN LEU2 | This study |
| PKL563 | RPL43B-HA | CEN HIS3 | This study |
| PKL580 | GST-loc1ΔC84 | Amp | This study |
| PKL581 | GST-loc1ΔC74 | Amp | This study |
| PKL582 | GST-loc1ΔN100 | Amp | This study |
| PKL583 | GST-loc1ΔN50 | Amp | This study |
| PKL584 | GST-L43ΔC helix | Amp | This study |
| PKL585 | GST-L43ΔN coil | Amp | This study |
| PKL586 | pET28a-loc1ΔC84 | Kan | This study |
| PKL588 | pET28a-loc1ΔC74 | Kan | This study |
| PKL590 | pET28a-loc1ΔC64 | Kan | This study |
| PKL592 | pET28a-loc1ΔN100 | Kan | This study |
| PKL650 | pET28a-loc1ΔN50 | Kan | This study |
| PKL652 | GST-L43ΔN helix | Amp | This study |
| PKL653 | GST-loc1ΔC64 | Amp | This study |
| PKL655 | loc1ΔN50-GFP | CEN LEU2 | This study |
| PKL657 | rpl43BΔN-HA | CEN URA3 | This study |
| PKL658 | rpl43BΔC-HA | CEN URA3 | This study |
| PKL659 | rpl43BΔM-HA | CEN URA3 | This study |
| PKL660 | rpl43B-4C-HA | CEN URA3 | This study |
| PKL661 | MBP-KAP121 | Amp | This study |
| PKL662 | pET28a-puf6-Nter | Kan | This study |
| PKL663 | pET28a-puf6-Cter | Kan | This study |
| PKL746 | LOC1 | 2μ LEU2 | This study |
| Oligo# | Sequence | Source | |
|---|---|---|---|
| probes | |||
| #1 (KLO48) | 5S | TCTGGTAGATATGGCCGCAACC | [39] |
| #2 (KLO49) | 18S | GTCTGGACCTGGTGAGTTTCCC | [39] |
| #3 (KLO50) | 20S | TCTTGCCCAGTAAAAGCTCTCATGC | [39] |
| #4 (KLO51) | 23S | TGTTACCTCTGGGCCCCGATTG | [39] |
| #5 (KLO52) | 35S, 27SA | TCCAGTTACGAAAATTCTTGTTTTTGACAA | [39] |
| #6 (KLO53) | 5.8S | CGCTGCGTTCTTCATCGATGCG | [39] |
| #7 (KLO54_ | 7S | GGCCAGCAATTTCAAGTTA | [39] |
| #9 (KLO56) | 25S | CCCGCCGTTTACCCGCGCTTGG | [39] |
| qPCR Primers | |||
| KLO323 | RPL5 | TAGCTGCTGCCTACTCCCACGA | [32] |
| KLO324 | RPL5 | GCAGCAGCCCAGTTGGTCAAA | [32] |
| KLO325 | RPL10 | TGTCTTGTGCCGGTGCGGAT | [32] |
| KLO326 | RPL10 | TGTCGACACGAGCGGCCAAA | [32] |
| KLO327 | RPL43 | CAAGAGAGGTGCTGCTGGTA | This study |
| KLO328 | RPL43 | GCAGTGGAGACAGTGTAAGCA | This study |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, K.-J.; Yueh, L.-Y.; Hsu, N.-H.; Lai, J.-S.; Lo, K.-Y. Puf6 and Loc1 Are the Dedicated Chaperones of Ribosomal Protein Rpl43 in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2019, 20, 5941. https://doi.org/10.3390/ijms20235941
Liang K-J, Yueh L-Y, Hsu N-H, Lai J-S, Lo K-Y. Puf6 and Loc1 Are the Dedicated Chaperones of Ribosomal Protein Rpl43 in Saccharomyces cerevisiae. International Journal of Molecular Sciences. 2019; 20(23):5941. https://doi.org/10.3390/ijms20235941
Chicago/Turabian StyleLiang, Kai-Jen, Le-Yun Yueh, Ning-Hsiang Hsu, Jui-Sheng Lai, and Kai-Yin Lo. 2019. "Puf6 and Loc1 Are the Dedicated Chaperones of Ribosomal Protein Rpl43 in Saccharomyces cerevisiae" International Journal of Molecular Sciences 20, no. 23: 5941. https://doi.org/10.3390/ijms20235941
APA StyleLiang, K. -J., Yueh, L. -Y., Hsu, N. -H., Lai, J. -S., & Lo, K. -Y. (2019). Puf6 and Loc1 Are the Dedicated Chaperones of Ribosomal Protein Rpl43 in Saccharomyces cerevisiae. International Journal of Molecular Sciences, 20(23), 5941. https://doi.org/10.3390/ijms20235941