1. Introduction
The leucine-rich repeats (LRR)-containing domain is noted to be evolutionarily conserved among many proteins correlated with innate immunity in an array of organisms, including invertebrates, vertebrates, and plants [
1]. Innate immunity is a conserved host response that serves as the host’s first line of defense by sensing pathogen-associated molecular patterns through germline-encoded pattern recognition receptors [
2]. For example, in mammals, activation of antimicrobial defenses and stimulation of the adaptive immune response is mediated by Toll-like receptors (TLRs) and nucleotide-binding oligomerization domain (NOD)-like receptors through the LRR domain in response to pathogens [
3].
Mosquitoes lack an adaptive immune system [
4] and so they are fully dependent on mounting an innate immune response to fight infection [
5,
6,
7,
8]. Research indicates that mosquito vectors and mosquito cell lines produce both humoral and cellular immune responses against invading pathogens [
9,
10]. Leucine-rich repeat immune proteins (LRIMs) are a mosquito-specific family of putative innate receptors [
11,
12,
13]. Two LRR proteins, LRIM1 (leucine-rich repeat immune protein 1) and APL1 (Anopheles Plasmodium-responsive leucine-rich repeat 1), have been recognized as major mosquito factors that regulate parasite infection and parasite loads [
14]. In
Anopheles mosquitoes,
LRIM1,
APL1C, and LRR-containing proteins activate complement-like defense responses against malaria by forming a disulphide-bridge that interacts with thioester-containing protein 1 (TEP1), a complement C3-like protein [
15,
16].
APL1 of
Anopheles gambiae is a family of variable LRR proteins required for cell-mediated protection as shown using a rodent model malaria parasite,
Plasmodium berghei [
17]. The
An. gambiae APL1 genomic locus circumscribes three distinct genes (
APL1A,
APL1B, and
APL1C), however, only the product of
APL1C acts as a
P. berghei antagonist [
17].
LRIM1 and
APL1C both play a significant role in the anti-
Plasmodium response. Accordingly, silencing of these genes results in an altered response against
Plasmodium infection [
13]. A genome-wide study found that variations or polymorphisms in the
LRIM1 and
APL1C proteins were correlated with resistance and susceptibility to
Plasmodium infection [
18]. Additional studies offered mechanisms for controlling
Plasmodium early during the infection process by targeting the ookinete or oocyst stages of oocyst [
19,
20]. Nonetheless, no detailed information has been generated for pathogen response by the LRR-containing proteins of immune factors in
Aedes mosquitoes, responsible for transmitting multiple emerging arboviruses.
Aedes aegypti (L.) is a vector for transmitting emergent arboviruses, including chikungunya, dengue, yellow fever, and Zika viruses. Chikungunya fever is primarily transmitted to humans through mosquito vectors,
Ae. aegypti and
Ae. albopictus. It is a viral disease belonging to the family, Togaviridae, and genus,
Alphavirus. Recent outbreaks of chikungunya fever occurred from 2004 to 2006 spanning Kenya in 2004 (Eastern/Central/Southern African, ECSA, CHIKV lineage) and the island of La Réunion in 2005-2006 (Indian Ocean CHIKV lineage). It later emerged in the New World in 2013 on St. Martin Island (Asian CHIKV lineage), eventually spreading throughout the Americas [
21,
22,
23,
24]. Over the span of the past 12 years, it is estimated that more than four million human cases of chikungunya infection have occurred worldwide [
25]. Symptoms of infection include a rash, fever, headache, joint pain, and muscle pain [
26] along with the chance of developing chronic musculoskeletal diseases [
27]. There is currently no vaccine available for CHIKV. Accordingly, controlling the mosquito vectors is the primary method utilized to reduce the risk of disease transmission.
Zika virus (ZIKV) was first discovered in 1947 and belongs to the family, Flaviviridae, genus,
Flavivirus (CDC 2016) [
28]. Spreading to the Oceania region, ZIKV caused outbreaks on Yap Island in Micronesia and French Polynesia in 2007 and 2013, respectively. In 2015, ZIKV was found to have reached Brazil, spreading throughout the Americas [
28,
29]. It is estimated that 1.5 million people have been infected by ZIKV in Brazil [
30]. ZIKV continues to spread to new areas. Transmission in the U.S. is a major public health risk, notably for the Gulf states, such as Florida and Texas, where ecological conditions are favorable for the primary vector,
Ae. aegypti, as well as an increasing likelihood for virus introduction by imported cases. Manifestations of ZIKV take on different forms, the most serious of which include birth defects in humans [
31] along with neurological complications that may result in Guillain-Barré syndrome, both of which are significant public health threats [
32].
Mosquitoes respond to infection using an array of molecular signaling pathways and immune effector proteins. A focus on the immune system response of
Ae. aegypti has unveiled a transcriptome analysis of genome-wide mechanisms that are implicated in defense against arbovirus infections [
33,
34,
35,
36,
37]. No sequence-structure-function relationships of mosquito leucine-rich repeat immune proteins in
Ae. aegypti in response to arboviruses are available, though the LRR-proteins have been compared with
An. gambiae and
Culex quinquefasciatus [
16]. Many studies of
LRIM1 and
APL1C have been reported as playing an important role in defense against
Plasmodium in
Anopheles [
13,
14,
15]. However, there is no study of
LRIM1 and
APL1C in the
Aedes in response to arboviruses. Therefore, we re-examined previous RNAseq data analysis and obtained 60 leucine-rich repeat related transcriptions of
Ae. aegypti (Accession number: GSE118858, accessed on: August 22, 2018,
https://www.ncbi.nlm.nih.gov/gds/?term=GSE118858) [
34]. We examined the
AaeLRIM1 and
AaeAPL1 gene expressions in response to both ZIKV and CHIKV infection using a time course study. Additionally, we investigated the developmental expressions of these genes in the eggs, larvae, pupae, and adults. The current study aims to improve our understanding of the transcription profiles of
Ae. aegypti LRR proteins during development and in response to arbovirus infection.
3. Discussion
Identification and characterization of genes related to
LRIM1 and
APL1C revealed novel innate immune factors and furthered our understanding of their presumed molecular functions. Waterhouse et al. (2010) used comparative sequenced genomes:
An. gambiae,
Ae. aegypti, and
Cx. quinquefasciatus revealed that mosquito LRIM proteins can be classified into four distinct subfamilies by a variable number of LRRs [
16]. Our phylogenetic tree of paralogue showed that
AaeAPL1 (or
AaeLRIM2) sequence-structure-function was most closely related to
AaeLRIM4.
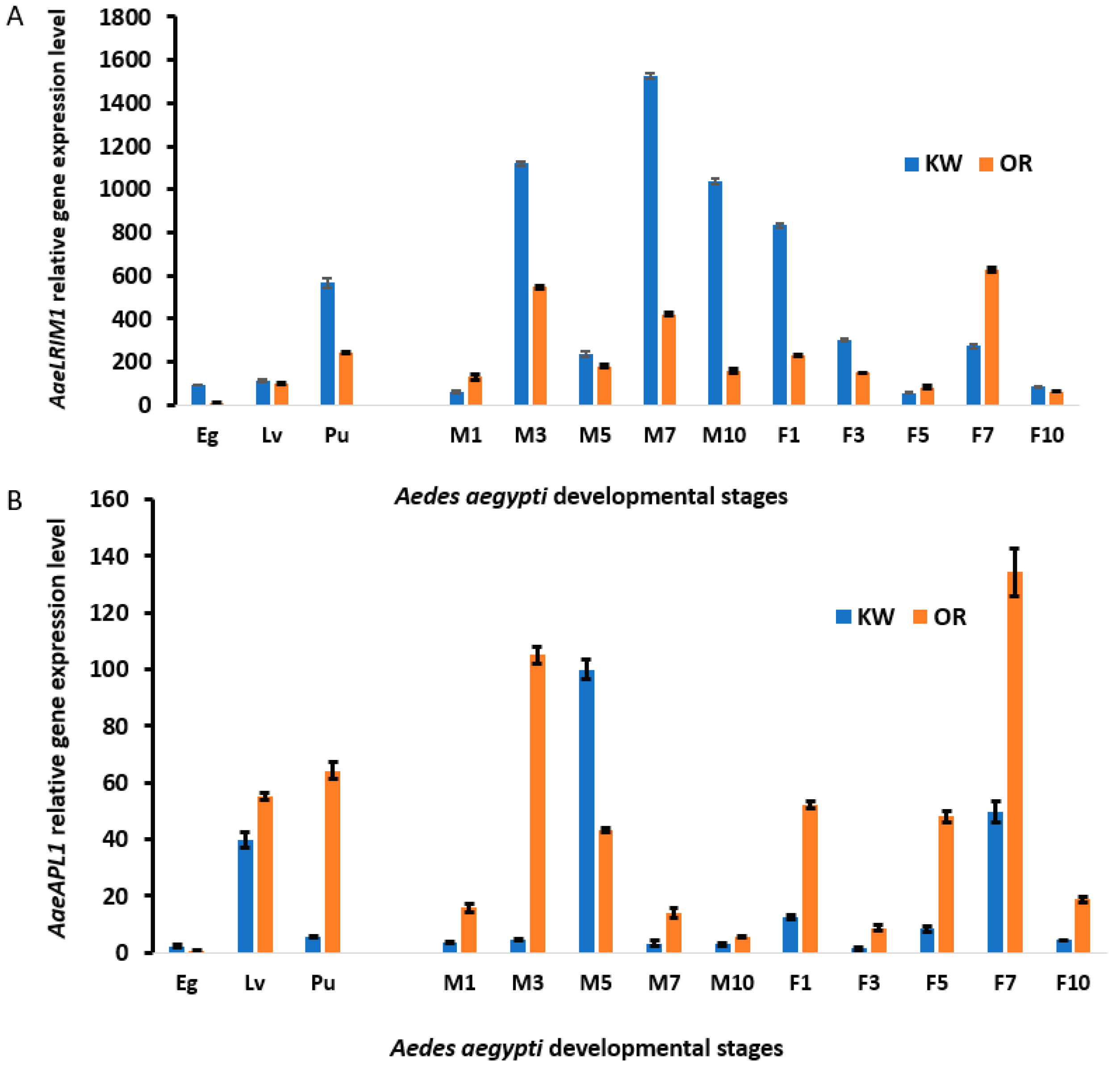
We analyzed developmental changes in the gene expression of LRR-containing proteins in
Ae. aegypti eggs, larvae, pupae, and adults. The nucleus gene LRR-containing proteins,
AaeLRIM1, AaeAPL1, and the other five paralogues are expressed <100-fold lower than the other nuclear genes, such as defensin, during all developmental stages examined [
40]. Our data show that the expression of
AaeLRIM1 (AAEL012086),
AaeAPL1 (AAEL009520-RA), and the other five paralogues is not only regulated by development, but also by the varying environmental origin (or permethrin resistant selected strain) of mosquito strains in
Ae. aegypti. For both immature and adult stages, we observed higher expression of
AaeLRIM1 than
AaeAPL1 in Key West
Ae. aegypti and higher expression of
AaeAPL1 than
AaeLRIM1 in Orlando
Ae. aegypti. These differences in responses may be attributable to differences in insecticide resistance among the Orlando (permethrin susceptible) and Key West (permethrin resistant) strains of
Ae. aegypti. However, we are unable to rule out that other differences between these two strains of
Ae. aegypti (e.g., geographic origin and founder effects) may contribute to the observed differences in gene expressions. For the immature, but not adult stages, expression of
AaeAPL1 and
AaeLRIM1 increased with the developmental stage. The strain effect for the analyses were modified by interactions with other factors, suggesting complex interactions between gene expression and immature stage, adult age, and sex. To our knowledge, this is the first exploration of
AaeLRIM gene expression during
Ae.
aegypti development.
In
Anopheles, two leucine-rich repeat (LRR) protein related genes,
LRIM1 and
APL1, have been shown to strongly affect
P. berghei development in the mosquito midgut and have been identified as major mosquito factors that regulate parasite loads [
11,
13,
14,
15,
41,
42,
43,
44]. In
Ae. aegypti, the likely LRIM1 orthologue is upregulated with other immune genes following infection with
Wolbachia bacteria, resulting in immune activation and shortened mosquito life spans [
45]. However, there is no information available for revealing
LRIM1 and
APL1 affected by arboviruses. Our data demonstrates that gene expression of LRR-containing proteins,
AaeLRIM1 and
AaeAPL1, not only affect regulated parasites in the
Anopheles, but are also altered by arbovirus infection in
Ae. aegypti. Biophysical analysis of
An. gambiae LRR proteins,
APL1A1,
APL1B, and
APL1C, can arrange an extended, flexible heterodimer with
LRIM1, providing a repertoire of functional innate immune complexes to protect
An. gambiae from a diverse array of pathogens [
14]. Future studies are needed to identify how proteins,
AaeLRIM1 and
AaeAPL1, may influence progression of infection of arboviruses in
Ae. aegypti. Some of the highest changes in expression of
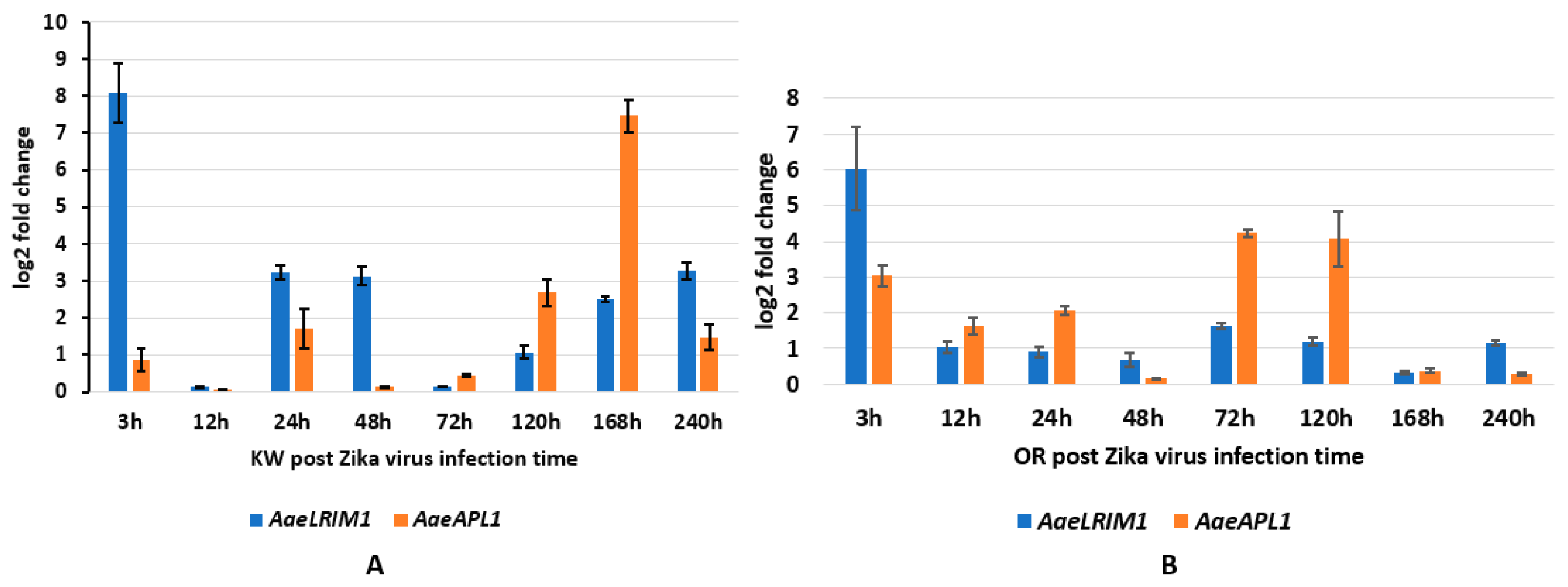
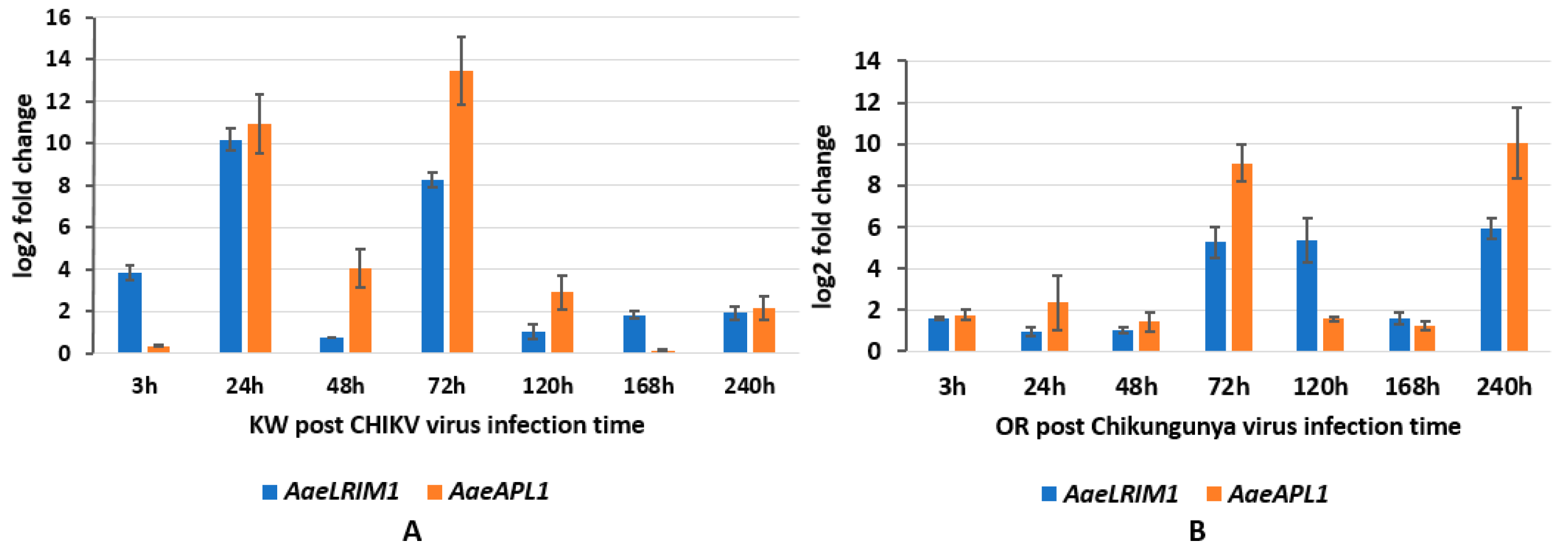
AaeAPL1 and
AaeLRIM1 for both strains of
Ae. aegypti occurred 24–72 h post infection with CHIKV, which approximates the time when
Ae. aegypti acquire disseminated infections [
24]. However, this response appears to be earlier for the Key West strain (approximately 24 h), which may suggest alterations in immune responses between permethrin susceptible and resistant
Ae. aegypti (Shin et al. unpublished data). Changes in the expression of
AaeLRIM1 and
AaeAPL1 followed a different pattern for ZIKV infected mosquitoes, suggesting gene expression changes depend on the particular arbovirus. For both strains of
Ae. aegypti,
AaeLRIM1 tended to be highest early during infection and declined at later points. In contrast, expression of
AaeAPL1 was low early during infection and higher at later measured times.
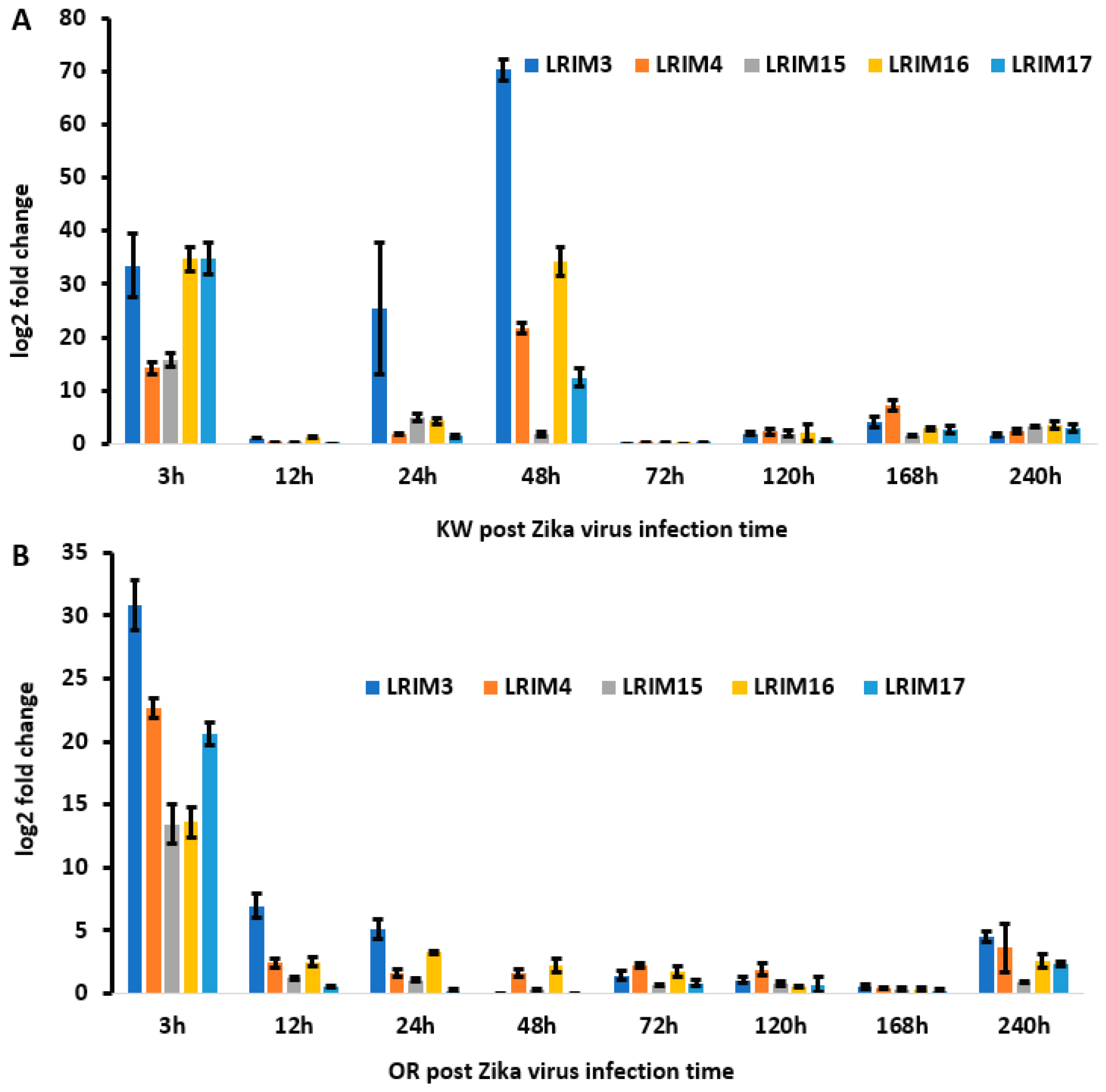
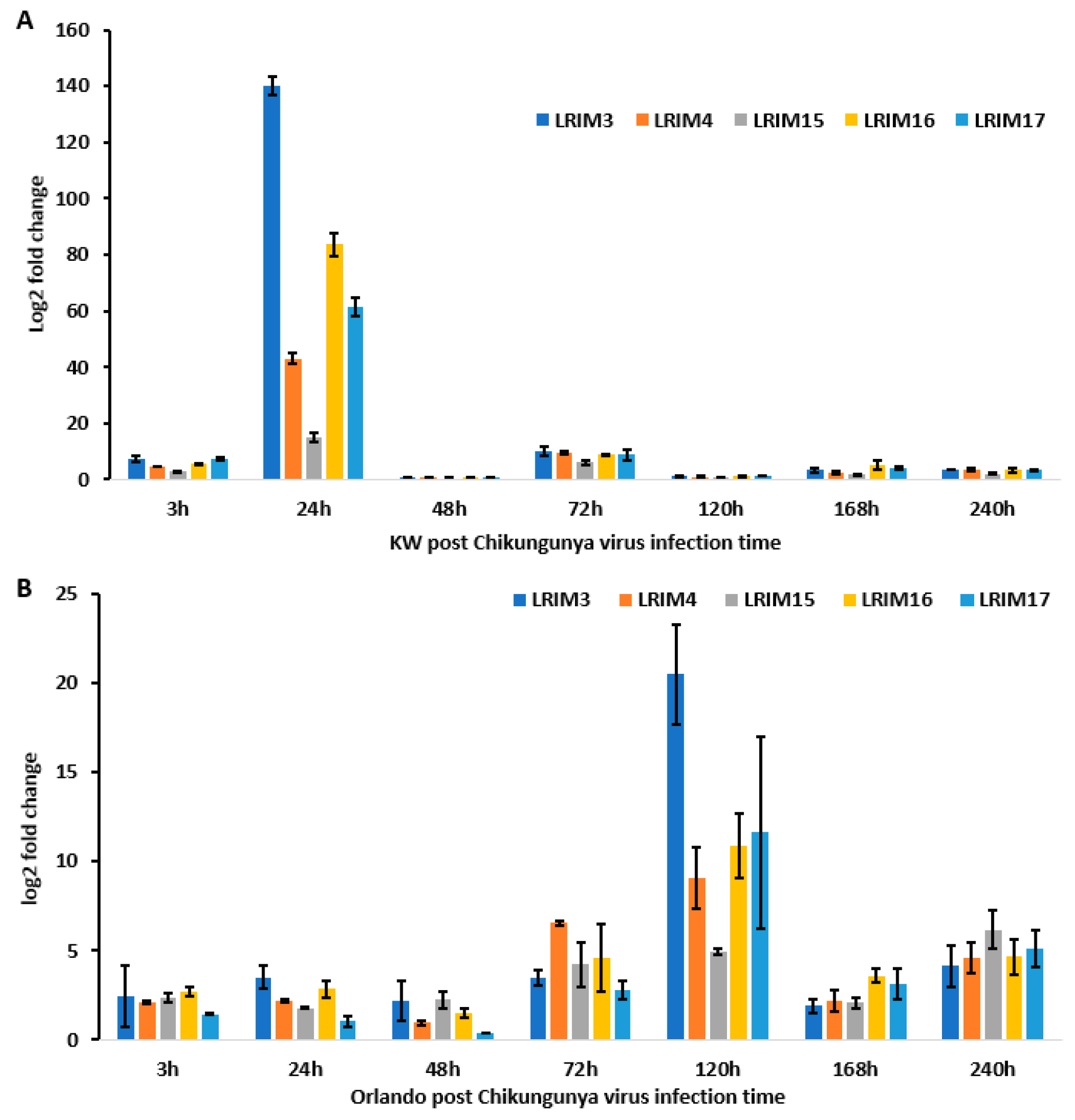
We also described the gene expression of five paralogues of AaeAPL1 (AaeLRIM3, AaeLRIM4, AaeLRIM15, AaeLRIM16, and AaeLRIM17) in Ae. aegypti following ingestion of arbovirus infected blood. The relative contributions of the five paralogues of AaeAPL1 differed in response to infection with either ZIKV or CHIKV, which may suggest differences in mounting an immune response between viruses of different families. Gene expression of AeaLRIM16 and AeaLRIM17 made the largest contributions to significant treatment effects for mosquitoes orally exposed to ZIKV. Along the same lines, AeaLRIM3 and AeaLRIM16 made the largest contributions to significant treatment effects for mosquitoes following oral ingestion of CHIKV infected blood. However, there were some common features observed in the gene expression of the paralogues of AaeAPL1. Gene expression of all five paralogues of AaeAPL1 were usually the highest within the first 48-hours following ingestion of ZIKV or CHIKV infected blood, with lower rates of expression later during the infection process.
Studying the immune system of mosquitoes provides insights into significant opportunities to bridge tissue damage, immune invasions mechanisms, and the immune response against pathogens [
4]. More importantly, unveiling a newfound understanding of mosquito immunity will shed light on the fight against disease-spreading pathogens, including ZIKV and CHIKV. Understanding the mechanisms that allow pathogens to grow and replicate in mosquitoes will provide insights into the mechanisms of mosquito-pathogen interactions. Finding exact immune evasion strategies of pathogens will help produce novel strategies that are effective at controlling them.
4. Materials and Methods
4.1. Mosquito Strains and Developmental Stages of Aedes aegypti
Ae. aegypti larvae were collected from Key West (24.55°N, 81.78°W), Florida, USA since 2011 and were initially tested for permethrin resistance, then subjected to permethrin selection for 15 generations and again assayed for resistance strain [
34]. Key West strain
Ae. aegypti, referred to as the resistant strain, was maintained at the Florida Medical Entomology Laboratory (FMEL) in Vero Beach, FL, USA. The Orlando population of
Ae. aegypti was collected from Orlando, FL, USA and reared in the Mosquito and Fly Research Unit, Center for Medical, Agricultural and Veterinary Entomology, ARS-USDA in Gainesville, FL since 1952. The Orlando strain is recognized as a permethrin susceptible strain of
Ae. aegypti [
46]. For the experiments, mosquito eggs were hatched and reared in a rearing chamber at 27 °C [
40]. We sampled eggs, larvae, pupae, and adults to measure developmentally regulated gene expression of Leucine-Rich Repeat Proteins in
Ae. aegypti. We collected 100 µg of eggs, 20 larvae at the third instar stage, 20 pupae, 10 adult mosquitoes, 10 males and 10 females at 1-day-old (teneral), 3-day-old, 5-day-old, 7-day-old, and 10-day-old adults. The experiments were repeated three times.
4.2. Chikungunya Virus and Zika Virus Infection
Four-day-old female adults were fed blood containing either CHIKV, ZIKV, or blood without virus as the control [
34,
40]. Isolates of the Indian Ocean lineage of CHIKV (LR2006-OPY1, GenBank accession: KT449801) from Réunion and the Asian lineage of ZIKV (strain PRVABC59, GenBank accession # KU501215.1) from Puerto Rico were cultured in African green monkey (Vero) cells and used in the mosquito infection study. Ten mosquitos were used in each sample and three independent experiments were conducted for each time point. The detailed procedures were described in the previous publication [
34,
40].
Arboviruses were propagated on monolayers of African green monkey kidney (Vero) cells at a multiplicity of infection of 0.1 and environmental conditions of 37 °C and 5% CO2. Monolayers of cells were covered with 24 mL media (M199 medium supplemented with 10% fetal bovine serum, penicillin/streptomycin, and mycostatin). For CHIKV and ZIKV, we collected the media with virus on days three and six, respectively, and mixed it with defibrinated bovine blood containing 0.005 M ATP. Mosquitoes were allowed to feed on infected blood for one hour feeding trials using a Hemotek membrane feeding system. Samples of blood were taken during feeding trials to allow for quantification of the amount of virus presented to mosquitoes. We also allowed mosquitoes to feed on a blood and media mixture without the presence of arboviruses (controls). Mosquitoes were fed 8.0-8.3 log10 pfu/mL of CHIKV for each strain and 6.4 log10 pfu/mL of ZIKV.
Following feeding trials, fully engorged mosquitoes were sorted using light microscopy (10×) (Carson, COPYRIGHT 2006 USA, UK, HK, CHINA, Made in China) and held in cages (h by d: 10 cm by 10 cm) maintained at a 12:12 h light:dark photoperiod and 30 °C. Mosquitoes were provided with an oviposition substrate and 10% sucrose solution on cotton pads. Cohorts of mosquitoes were killed and stored at −80 °C at the following sample periods after ingesting infected blood: 3-h, 12-h (ZIKV only), 24-hours, 48-hours, 72-hours, 120-hours, 168-hours, and 240-hours. Mosquitoes were deprived of sucrose, but not water 1-day before trials used to measure transmission on 3, 5, 7, and 10 days following ingestion of ZIKV and CHIKV infection blood.
4.3. RNA Extraction
Samples were homogenized with a plastic pestle in 1.5 mL tubes. Total RNAs were extracted using TRIzol reagent according to the manufacturer’s instructions (Ambion, Life Technologies, Carlsbad, CA, USA) following the standard protocol [
40]. The RNA samples were digested by DNase I (RNase-free), according to the manufacturer’s instructions (Thermo Scientific, Wilmington, DE, USA). The purified RNA samples were quantitated by an NANODROP 2000 Spectrophotometer (Thermo Scientific, Wilmington, DE, USA).
4.4. cDNA Synthesis and qPCR Amplification
cDNAs from 2 µg of total purified RNA were synthesized using a Cloned AMV First-Strand cDNA Synthesis Kit Invitrogen™ and Oligo (dT) 20 primer, according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA). The reaction was terminated by heat inactivation at 95 °C for 5 min. The cDNA samples for qPCR from the developmental stages, infected treatment, and controls were diluted by adding 80 µL ddH
2O to 20 µl reaction solution [
47].
The quantitative PCR (qPCR) assay for target genes,
AaeLRIM1 and
AaeAPL1, and reference gene,
AaeActin, in
Ae. aegypti was achieved using a BIO-RAD C1000 Touch Thermal Cycler, CFX 96™ Real-Time System (BIO-RAD, Hercules, CA, USA). The qPCR reaction mixture with a volume of 15 µl in Multiwell Plates 96 contained 1 µl diluted cDNA, 0.5 µM primers, and 1× master mix of PowerUP SYBR
® Green Master Mix (Applied Biosystems, Thermo Fisher Scientific, Foster City, CA, USA). In every qPCR run,
AaeActin was employed as an internal control to normalize for variation in the amount of cDNA template. The PCR primers for
AaeLRPIM, and
AaeAPL1 genes were designed from the coding region based on GenBank, Accession Number using Primer3
http://primer3.ut.ee (accessed on: Jul 20, 2017) (
Table 6). The qPCR thermal cycling parameters were the same as previous publication [
48]. Relative expression levels were calculated as follows for the developmental stages. First,
AaeLRIM/AaeAPL1 transcript levels relative to a standard (
AaeActin) were calculated using the formula,
ΔCT = CT (
AaeLRIM/AaeAPL1) − CT (
AaeActin). Second, an average
ΔCT value for each sample was calculated. Third, relative expression levels were calculated using the equation, 10,000 × 2
[−average ΔCT] [
40,
48]. Relative expression levels were calculated as follows for the treatment and control adults. First,
AaeLRIM1 or
AaeAPL1 transcript levels relative to a standard (
AaeActin) were calculated using the formula,
ΔCT = CT (
AaeLRIM1/AaeAPL1) − CT (
AaeActin). Then,
ΔΔCT=
ΔCT (infected) −
ΔCT (control) value for each sample was calculated. Third, relative expression levels were calculated using the equation, 1 × 2
[−average ΔΔCT] [
40,
48,
49,
50,
51].
4.5. RNA-Seq Library Sequencing, Data Mining, and RNA-seq Analysis
RNA sequencing and RNA-seq analysis were featured in a previous publication [
34]. Gene expression was assessed by counting the number of mapped reads for each transcript [
52]. Significantly up- and downregulated genes were determined using the adjusted
p-value (p-adj), log2 fold-change (log2FC), or both for downstream analysis. The RNA-seq data have been deposited to NCBI (
https://www.ncbi.nlm.nih.gov/gds/?term=GSE118858). RNA-seq analysis data provided 60 leucine-rich repeat related transcriptions in the
Ae. aegypti in response to Zika virus (
Table S1A–D).
4.6. Statistical Analysis
For this study, we were interested in measuring the magnitude of gene expression of components of the leucine-rich repeats (LRR)-containing domain as well as the relationship between gene expressions. So, we used a multivariate analysis of variance approach (MANOVA), which enabled us to simultaneously measure various treatment effects (e.g., developmental stage, infection with ZIKV and CHIKV) on the expression of multiple genes. Further, the MANOVA is associated with standardized canonical coefficients, which describe the relative contribution of each of the variables (genes) to a treatment effect and their direction to one another (positive or negative) [
53].
Multivariate analysis of variance (MANOVA) and ANOVA were used to measure developmentally regulated gene expression of AaeLRIM1 and AaeAPL1. The relative contribution and relationship of AaeLRIM1 and AaeAPL1 to developmental treatment effects were assessed using standardized canonical coefficients (SCC) (PROC GLM, SAS 9.22). When significant effects were detected, we used univariate comparisons among treatment least-squares means for the developmental stages (Tukey-Kramer method). Separate analyses were performed for the immature stages and adult stages. Similarly, we used MANOVA to measure the expression of AaeLRIM1 or AaeAPL1 following ingestion of CHIKV and ZIKV infected blood. We tested for all main treatment factors and interactions.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}