Enterococcus faecium L-15 Cell-Free Extract Improves the Chondrogenic Differentiation of Human Dental Pulp Stem Cells

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
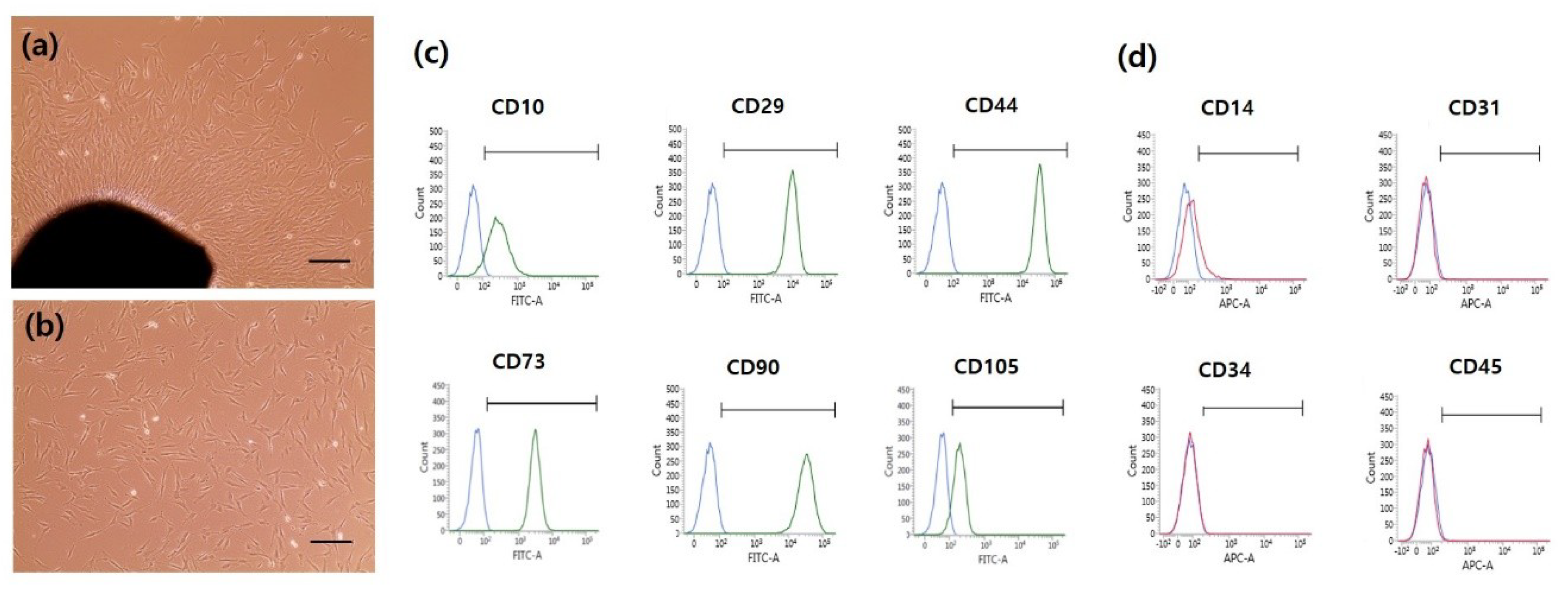
2.1. Human DPSC Characterization and Isolation
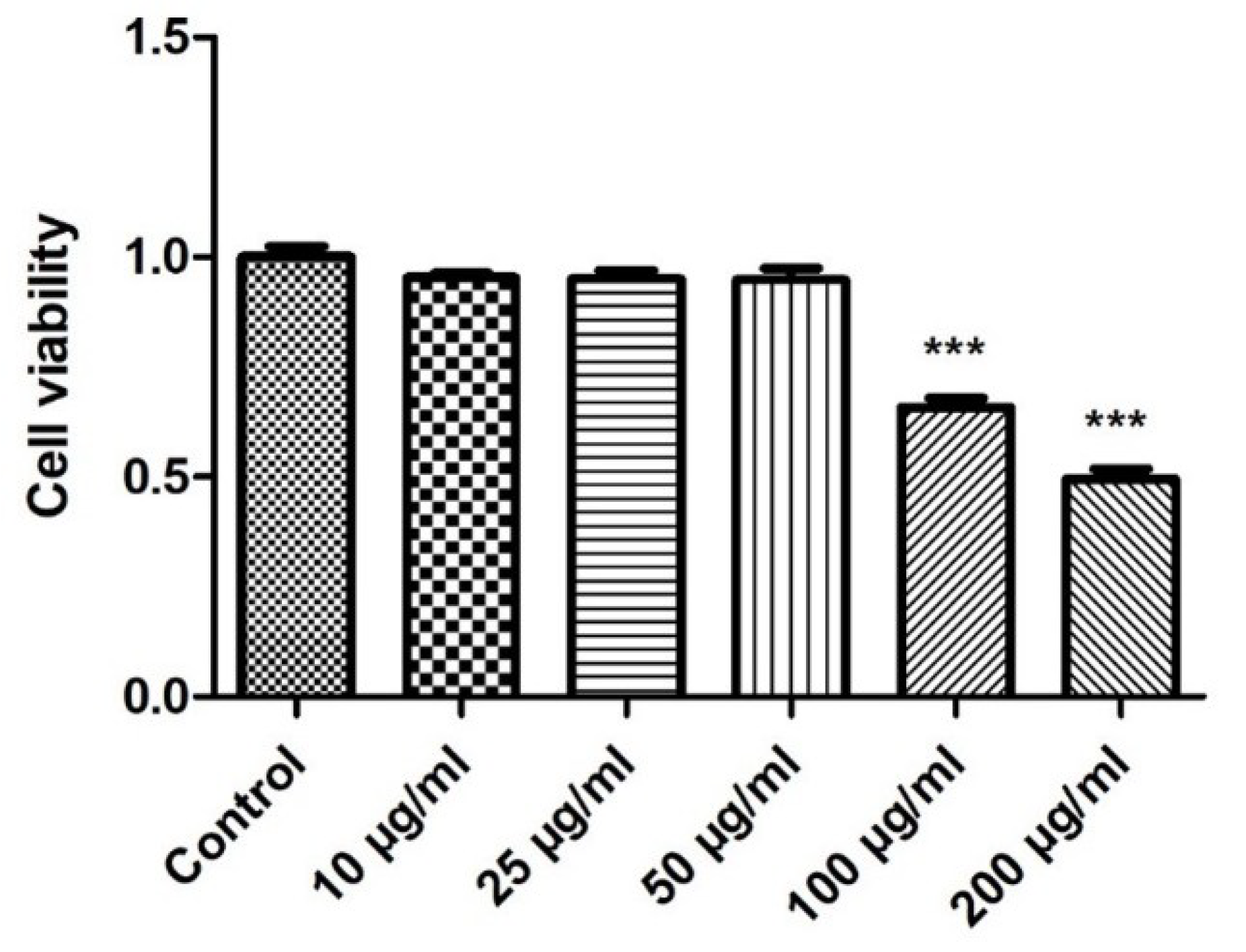
2.2. Effect of E. faecium L-15 Extract (L-15) on hDPSC Viability
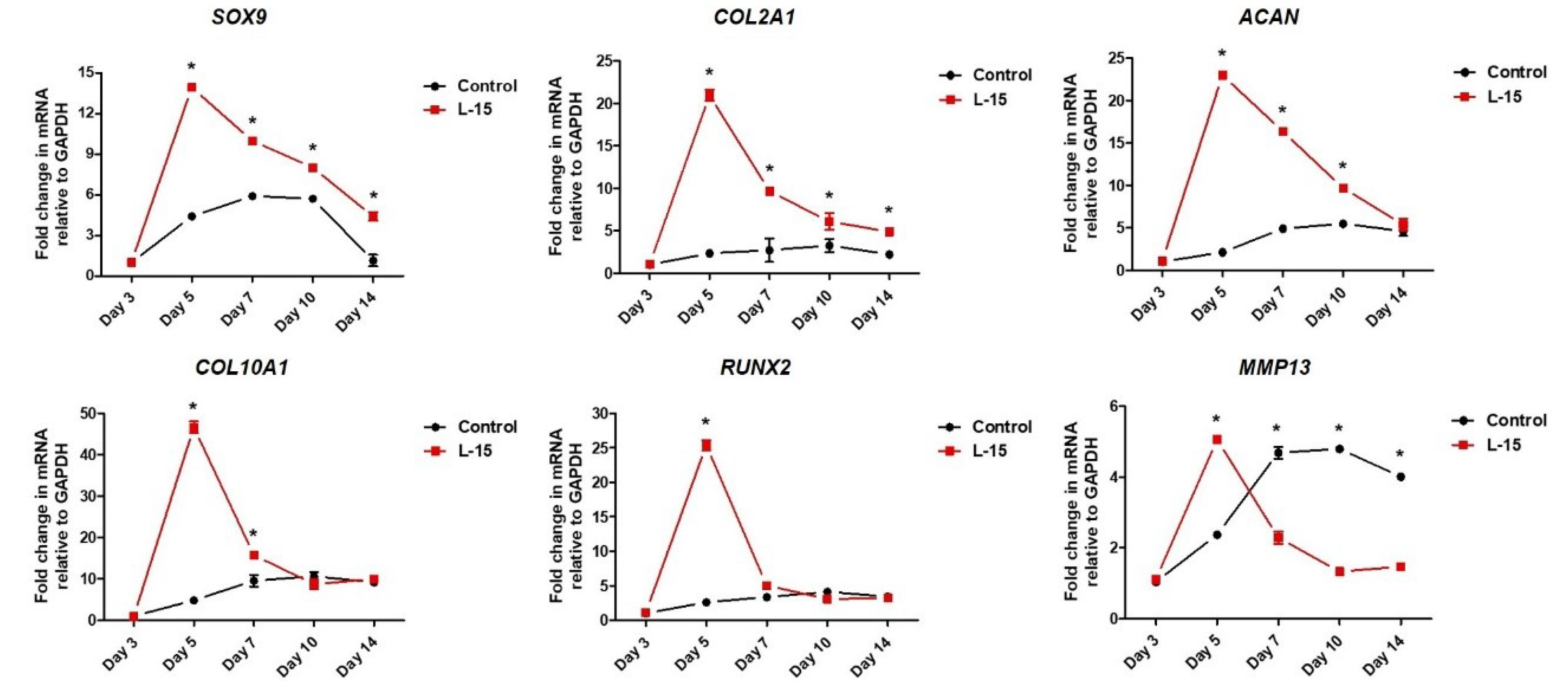
2.3. L-15 Extract Promotes Early-Stage Chondrogenic Differentiation
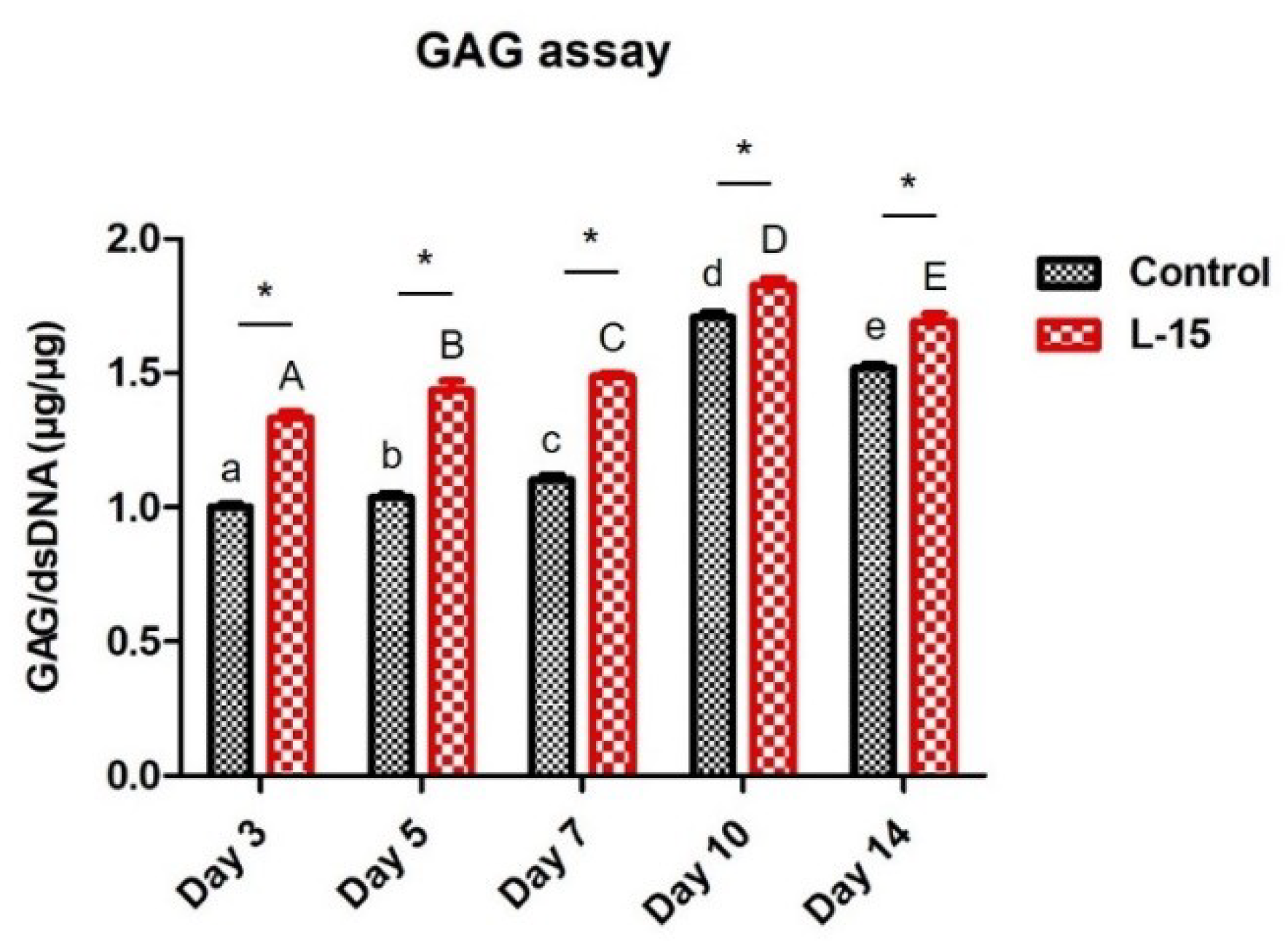
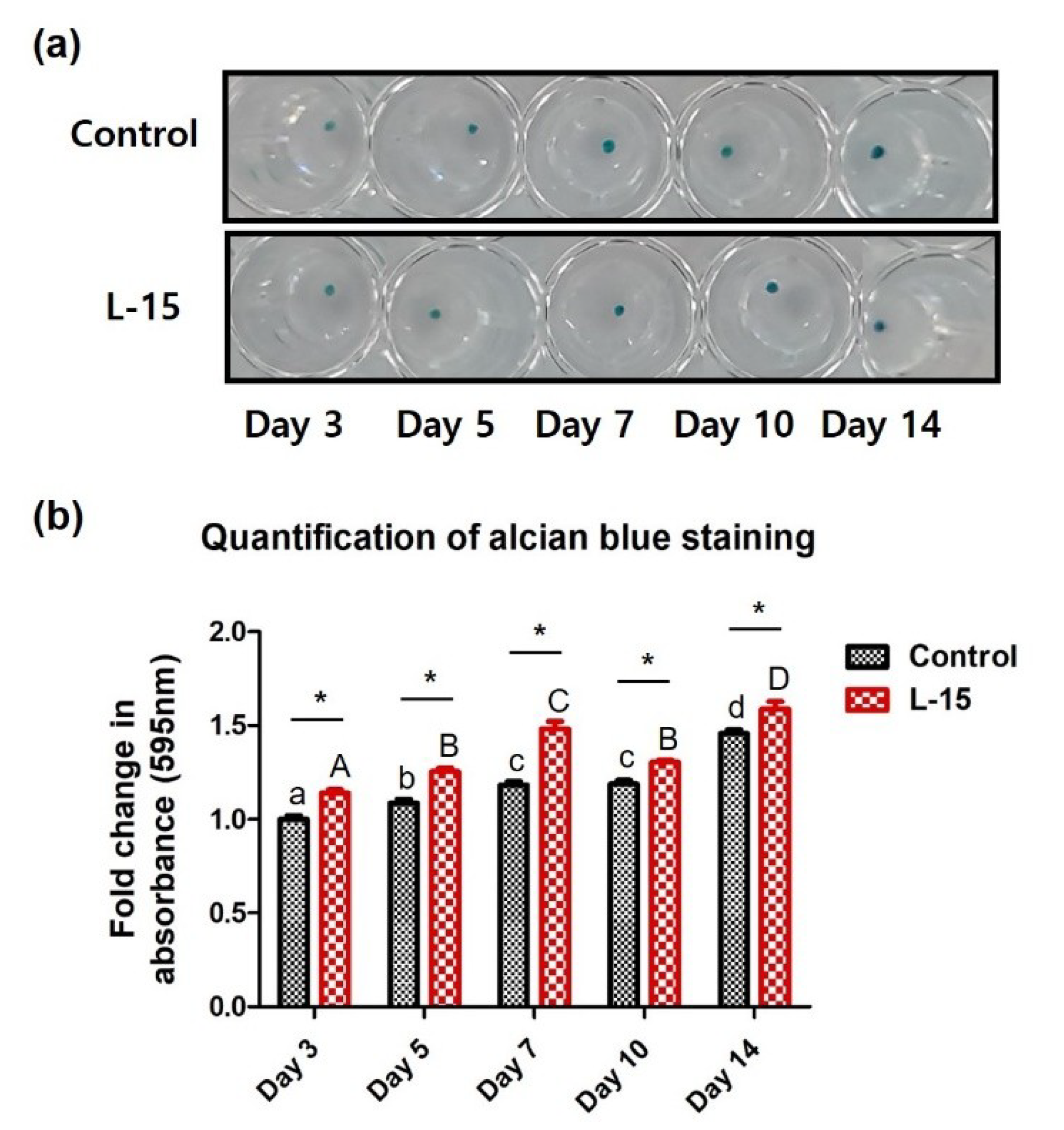
2.4. L-15 Extract Promotes ECM Formation during Chondrogenic Differentiation
3. Discussion
4. Materials and Methods
4.1. Preparation of the E. faecium Extract
4.2. Isolation and Expansion of Human Dental Pulp Stem Cells (hDPSCs)
4.3. Characterization of hDPSCs by Fluorescence-Activated Cell Sorting (FACS)
4.4. Chondrogenic Differentiation of hDPSCs
4.5. Cell Viability Assay
4.6. Real-Time Quantitative Polymerase Chain Reaction (PCR)
4.7. Quantitative Analysis of Glycosaminoglycan (GAG)
4.8. Quantification of Proteoglycan Content
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bornes, T.D.; Adesida, A.B.; Jomha, N.M. Mesenchymal stem cells in the treatment of traumatic articular cartilage defects: A comprehensive review. Arthritis Res. Ther. 2014, 16, 432. [Google Scholar] [CrossRef]
- Caldwell, K.L.; Wang, J. Cell-based articular cartilage repair: The link between development and regeneration. Osteoarthritis Cartil. 2015, 23, 351–362. [Google Scholar] [CrossRef]
- Gillogly, S.D.; Arnold, R.M. Autologous chondrocyte implantation and anteromedialization for isolated patellar articular cartilage lesions: 5- to 11-year follow-up. Am. J. Sports Med. 2014, 42, 912–920. [Google Scholar] [CrossRef]
- Kwak, S.K.; Kern, B.S.; Ferkel, R.D.; Chan, K.W.; Kasraeian, S.; Applegate, G.R. Autologous chondrocyte implantation of the ankle: 2- to 10-year results. Am. J. Sports Med. 2014, 42, 2156–2164. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Hincke, M.T. Mesenchymal stem cell-based tissue engineering strategies for repair of articular cartilage. Histol. Histopathol. 2014, 29, 669–689. [Google Scholar] [CrossRef]
- Johnson, K.; Zhu, S.; Tremblay, M.S.; Payette, J.N.; Wang, J.; Bouchez, L.C.; Meeusen, S.; Althage, A.; Cho, C.Y.; Wu, X.; et al. A stem cell-based approach to cartilage repair. Science 2012, 336, 717–721. [Google Scholar] [CrossRef]
- Lee, W.Y.; Wang, B. Cartilage repair by mesenchymal stem cells: Clinical trial update and perspectives. J. Orthop. Transl. 2017, 9, 76–88. [Google Scholar] [CrossRef]
- Sharpe, P.T. Dental mesenchymal stem cells. Development 2016, 143, 2273–2280. [Google Scholar] [CrossRef] [Green Version]
- Al-Zer, H.; Apel, C.; Heiland, M.; Friedrich, R.E.; Jung, O.; Kroeger, N.; Eichhorn, W.; Smeets, R. Enrichment and Schwann Cell Differentiation of Neural Crest-derived Dental Pulp Stem Cells. In Vivo 2015, 29, 319–326. [Google Scholar]
- Didilescu, A.C.; Rusu, M.C.; Nini, G. Dental pulp as a stem cell reservoir. Rom. J. Morphol. Embryol. 2013, 54, 473–478. [Google Scholar]
- Guimaraes, E.T.; Cruz Gda, S.; Almeida, T.F.; Souza, B.S.; Kaneto, C.M.; Vasconcelos, J.F.; Santos, W.L.; Santos, R.R.; Villarreal, C.F.; Soares, M.B. Transplantation of stem cells obtained from murine dental pulp improves pancreatic damage, renal function, and painful diabetic neuropathy in diabetic type 1 mouse model. Cell Transpl. 2013, 22, 2345–2354. [Google Scholar] [CrossRef]
- Ellis, K.M.; O’Carroll, D.C.; Lewis, M.D.; Rychkov, G.Y.; Koblar, S.A. Neurogenic potential of dental pulp stem cells isolated from murine incisors. Stem Cell Res. Ther. 2014, 5, 30. [Google Scholar] [CrossRef]
- Bojic, S.; Volarevic, V.; Ljujic, B.; Stojkovic, M. Dental stem cells—Characteristics and potential. Histol. Histopathol. 2014, 29, 699–706. [Google Scholar] [CrossRef]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef]
- Liu, H.; Yang, C.; Jing, Y.; Li, Z.; Zhong, W.; Li, G. Ability of lactic acid bacteria isolated from mink to remove cholesterol: In vitro and in vivo studies. Can. J. Microbiol. 2013, 59, 563–569. [Google Scholar] [CrossRef]
- Ispirli, H.; Demirbas, F.; Dertli, E. Characterization of functional properties of Enterococcus faecium strains isolated from human gut. Can. J. Microbiol. 2015, 61, 861–870. [Google Scholar] [CrossRef]
- Park, J.E.; Oh, S.H.; Cha, Y.S. Lactobacillus plantarum LG42 isolated from gajami sik-hae inhibits adipogenesis in 3T3-L1 adipocyte. Biomed. Res. Int. 2013, 2013, 460927. [Google Scholar] [CrossRef]
- Park, O.J.; Kim, J.; Yang, J.; Yun, C.H.; Han, S.H. Enterococcus faecalis Inhibits Osteoblast Differentiation and Induces Chemokine Expression. J. Endod. 2015, 41, 1480–1485. [Google Scholar] [CrossRef]
- Mansour, N.M.; Heine, H.; Abdou, S.M.; Shenana, M.E.; Zakaria, M.K.; El-Diwany, A. Isolation of Enterococcus faecium NM113, Enterococcus faecium NM213 and Lactobacillus casei NM512 as novel probiotics with immunomodulatory properties. Microbiol. Immunol. 2014, 58, 559–569. [Google Scholar] [CrossRef] [Green Version]
- Abdhul, K.; Ganesh, M.; Shanmughapriya, S.; Kanagavel, M.; Anbarasu, K.; Natarajaseenivasan, K. Antioxidant activity of exopolysaccharide from probiotic strain Enterococcus faecium (BDU7) from Ngari. Int. J. Biol. Macromol. 2014, 70, 450–454. [Google Scholar] [CrossRef]
- Marcacci, M.; Filardo, G.; Kon, E. Treatment of cartilage lesions: What works and why? Injury 2013, 44, S11–S15. [Google Scholar] [CrossRef]
- Jevotovsky, D.S.; Alfonso, A.R.; Einhorn, T.A.; Chiu, E.S. Osteoarthritis and stem cell therapy in humans: A systematic review. Osteoarthritis Cartilage 2018. [Google Scholar] [CrossRef] [PubMed]
- Marmotti, A.; Mattia, S.; Castoldi, F.; Barbero, A.; Mangiavini, L.; Bonasia, D.E.; Bruzzone, M.; Dettoni, F.; Scurati, R.; Peretti, G.M. Allogeneic Umbilical Cord-Derived Mesenchymal Stem Cells as a Potential Source for Cartilage and Bone Regeneration: An In Vitro Study. Stem Cells Int. 2017, 2017, 1732094. [Google Scholar] [CrossRef]
- Goldring, M.B.; Goldring, S.R. Articular cartilage and subchondral bone in the pathogenesis of osteoarthritis. Ann. N. Y. Acad. Sci. 2010, 1192, 230–237. [Google Scholar] [CrossRef]
- Studer, D.; Millan, C.; Ozturk, E.; Maniura-Weber, K.; Zenobi-Wong, M. Molecular and biophysical mechanisms regulating hypertrophic differentiation in chondrocytes and mesenchymal stem cells. Eur. Cell Mater. 2012, 24, 118–135. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, T.; Lemaitre, V.; D’Armiento, J.; Okada, Y. Matrix metalloproteinases, a disintegrin and metalloproteinases, and a disintegrin and metalloproteinases with thrombospondin motifs in non-neoplastic diseases. Pathol. Int. 2010, 60, 477–496. [Google Scholar] [CrossRef] [Green Version]
- Roach, H.I.; Yamada, N.; Cheung, K.S.; Tilley, S.; Clarke, N.M.; Oreffo, R.O.; Kokubun, S.; Bronner, F. Association between the abnormal expression of matrix-degrading enzymes by human osteoarthritic chondrocytes and demethylation of specific CpG sites in the promoter regions. Arthritis Rheumatol. 2005, 52, 3110–3124. [Google Scholar] [CrossRef] [Green Version]
- Aref-Eshghi, E.; Liu, M.; Harper, P.E.; Dore, J.; Martin, G.; Furey, A.; Green, R.; Rahman, P.; Zhai, G. Overexpression of MMP13 in human osteoarthritic cartilage is associated with the SMAD-independent TGF-beta signalling pathway. Arthritis Res. Ther. 2015, 17, 264. [Google Scholar] [CrossRef]
- Neuhold, L.A.; Killar, L.; Zhao, W.; Sung, M.L.; Warner, L.; Kulik, J.; Turner, J.; Wu, W.; Billinghurst, C.; Meijers, T.; et al. Postnatal expression in hyaline cartilage of constitutively active human collagenase-3 (MMP-13) induces osteoarthritis in mice. J. Clin. Investig. 2001, 107, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Sampson, E.R.; Jin, H.; Li, J.; Ke, Q.H.; Im, H.J.; Chen, D. MMP13 is a critical target gene during the progression of osteoarthritis. Arthritis Res. Ther. 2013, 15, R5. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Bai, Y.; Liu, C.; Dou, C.; Li, J.; Xiang, J.; Zhao, C.; Xie, Z.; Xiang, Q.; Dong, S. Hypertrophic differentiation of mesenchymal stem cells is suppressed by xanthotoxin via the p38MAPK/HDAC4 pathway. Mol. Med. Rep. 2017, 16, 2740–2746. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, A.J.; Converse, G.L.; Hopkins, R.A.; Detamore, M.S. The bioactivity of cartilage extracellular matrix in articular cartilage regeneration. Adv. Healthc. Mater. 2015, 4, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Wylie, J.D.; Hartley, M.K.; Kapron, A.L.; Aoki, S.K.; Maak, T.G. What is the effect of matrices on cartilage repair? A systematic review. Clin. Orthop. Relat. Res. 2015, 473, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Kiyotake, E.A.; Beck, E.C.; Detamore, M.S. Cartilage extracellular matrix as a biomaterial for cartilage regeneration. Ann. N. Y. Acad. Sci. 2016, 1383, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.; Borges, S.; Teixeira, P. Selection of potential probiotic Enterococcus faecium isolated from Portuguese fermented food. Int. J. Food Microbiol. 2014, 191, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, N.; Arroyo, R.; Calzada, J.; Peiroten, A.; Medina, M.; Rodriguez, J.M.; Fernandez, L. Evaluation of technological properties of Enterococcus faecium CECT 8849, a strain isolated from human milk, for the dairy industry. Appl. Microbiol. Biotechnol. 2016, 100, 7665–7677. [Google Scholar] [CrossRef] [PubMed]
- Park, D.Y.; Ahn, Y.T.; Huh, C.S.; Jeon, S.M.; Choi, M.S. The inhibitory effect of Lactobacillus plantarum KY1032 cell extract on the adipogenesis of 3T3-L1 Cells. J. Med. Food 2011, 14, 670–675. [Google Scholar] [CrossRef]
- Tong, Z.; Ma, J.; Tan, J.; Huang, L.; Ling, J. Effects of inactivated Enterococcus faecalis on the proliferation and osteogenic induction of osteoblasts. Mol. Med. Rep. 2016, 14, 5125–5133. [Google Scholar] [CrossRef] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Park, S.; Kim, K.; Ku, S.; Seo, J.; Roh, S. Enterococcus faecium L-15 Cell-Free Extract Improves the Chondrogenic Differentiation of Human Dental Pulp Stem Cells. Int. J. Mol. Sci. 2019, 20, 624. https://doi.org/10.3390/ijms20030624
Kim H, Park S, Kim K, Ku S, Seo J, Roh S. Enterococcus faecium L-15 Cell-Free Extract Improves the Chondrogenic Differentiation of Human Dental Pulp Stem Cells. International Journal of Molecular Sciences. 2019; 20(3):624. https://doi.org/10.3390/ijms20030624
Chicago/Turabian StyleKim, Hyewon, Sangkyu Park, Kichul Kim, Seockmo Ku, Jeongmin Seo, and Sangho Roh. 2019. "Enterococcus faecium L-15 Cell-Free Extract Improves the Chondrogenic Differentiation of Human Dental Pulp Stem Cells" International Journal of Molecular Sciences 20, no. 3: 624. https://doi.org/10.3390/ijms20030624
APA StyleKim, H., Park, S., Kim, K., Ku, S., Seo, J., & Roh, S. (2019). Enterococcus faecium L-15 Cell-Free Extract Improves the Chondrogenic Differentiation of Human Dental Pulp Stem Cells. International Journal of Molecular Sciences, 20(3), 624. https://doi.org/10.3390/ijms20030624