hCMV-Mediated Immune Escape Mechanisms Favor Pathogen Growth and Disturb the Immune Privilege of the Eye

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
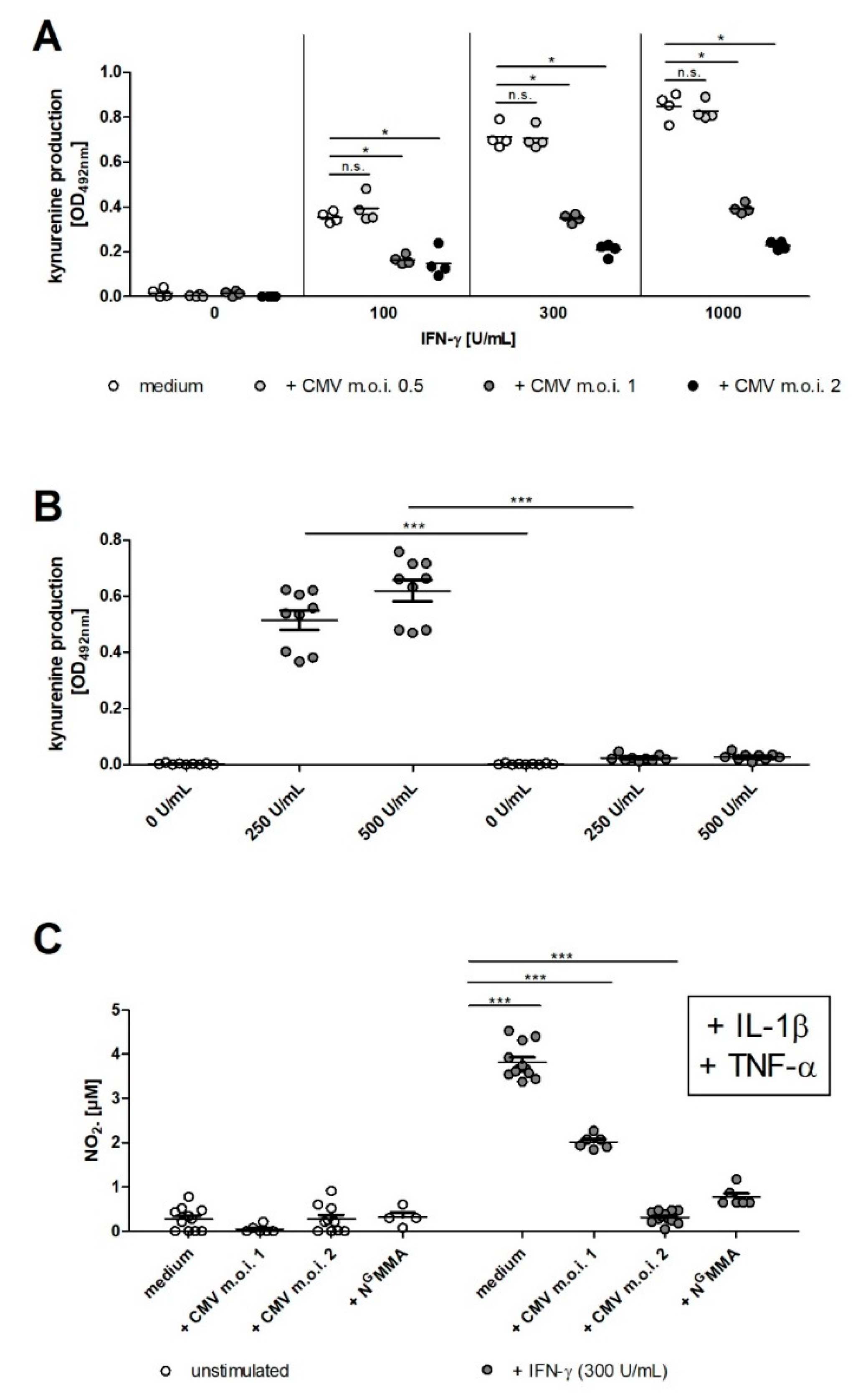
2.1. hCMV Controls Induction of IFN-γ Dependent IDO Expression
2.2. IDO1-Mediated Antibacterial and Antiparasitic Effects are Lost in hRPE Cells upon CMV Infection
2.3. CMV Infection Results in Lower IDO1 and iNOS Activity in Human RPE Cells
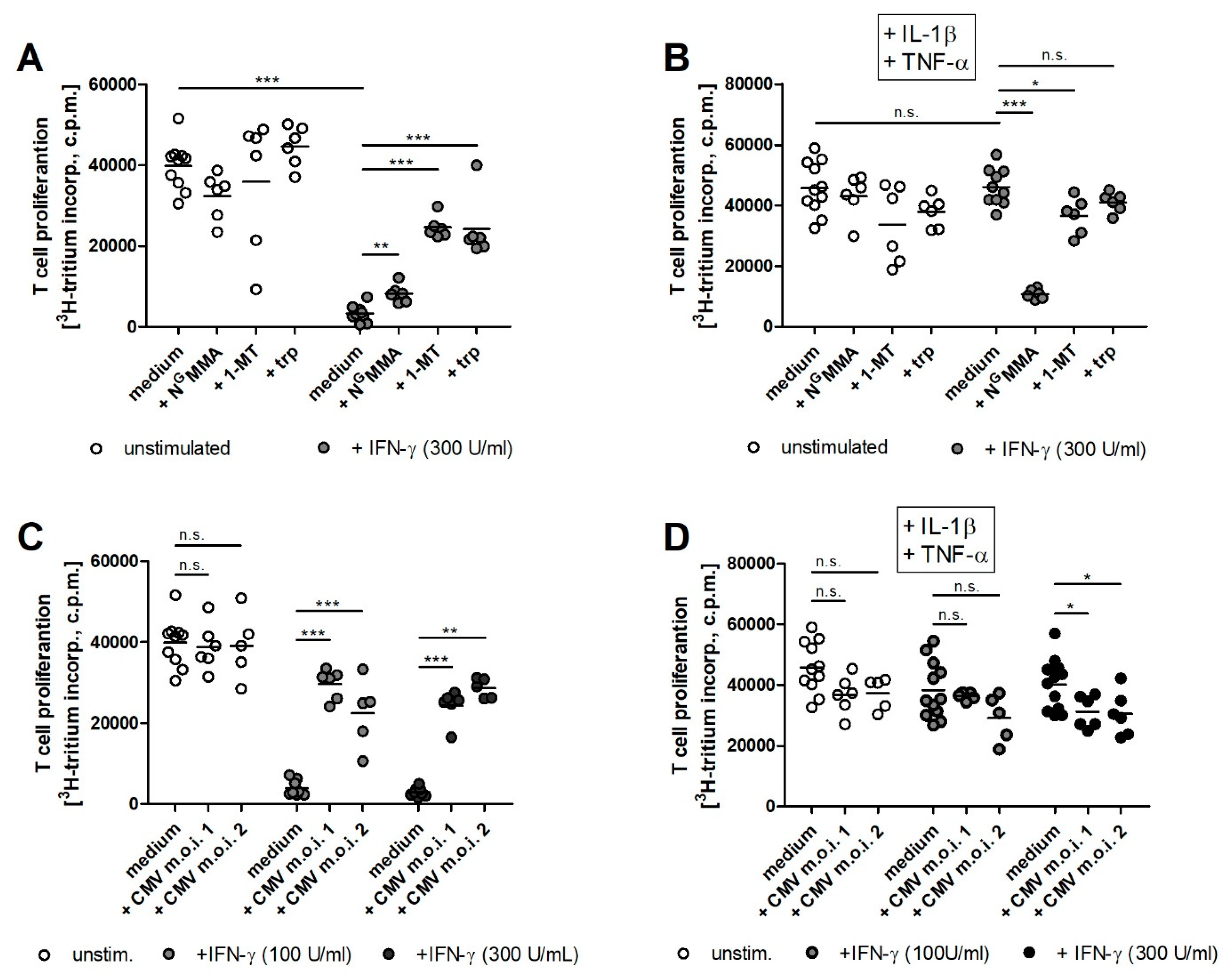
2.4. iNOS Activity and CMV Infection Impair IDO1-Mediated Immunoregulatory Effects in Human RPE Cells
3. Discussion
4. Material and Methods
4.1. Cells, Media and Reagents
4.2. Flow Cytometry
4.3. Stimulation of hRPE Cells
4.4. Kynurenine Assay
4.5. Nitric Oxide Assay
4.6. CMV Infection
4.7. Determination of Toxoplasma Growth
4.8. Determination of Bacterial Growth
4.9. T Cell Proliferation Assay
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Lachmann, R.; Loenenbach, A.; Waterboer, T.; Brenner, N.; Pawlita, M.; Michel, A.; Thamm, M.; Poethko-Müller, C.; Wichmann, O.; Wiese-Posselt, M. Cytomegalovirus (CMV) seroprevalence in the adult population of Germany. PLoS ONE 2018, 13, e0200267. [Google Scholar] [CrossRef]
- Melendez-Munoz, R.; Marchalik, R.; Jerussi, T.; Dimitrova, D.; Nussenblatt, V.; Beri, A.; Rai, K.; Wilder, J.S.; Barrett, A.J.; Battiwalla, M.; et al. Cytomegalovirus Infection Incidence and Risk Factors Across Diverse Hematopoietic Cell Transplantation Platforms Using a Standardized Monitoring and Treatment Approach: A Comprehensive Evaluation from a Single Institution. Biol. Blood Marrow Transpl. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fowler, K.B.; Boppana, S.B. Congenital cytomegalovirus infection. Semin. Perinatol. 2018, 42, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Faith, S.C.; Durrani, A.F.; Jhanji, V. Cytomegalovirus keratitis. Curr. Opin. Ophthalmol. 2018, 29, 373–377. [Google Scholar] [CrossRef]
- Laovirojjanakul, W.; Thanathanee, O. Opportunistic ocular infections in the setting of HIV. Curr. Opin. Ophthalmol. 2018, 29, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Van der Heiden, P.; Marijt, E.; Falkenburg, F.; Jedema, I. Control of Cytomegalovirus Viremia after Allogeneic Stem Cell Transplantation: A Review on CMV-Specific T Cell Reconstitution. Biol. Blood Marrow Transpl. 2018, 24, 1776–1782. [Google Scholar] [CrossRef]
- Nikitina, E.; Larionova, I.; Choinzonov, E.; Kzhyshkowska, J. Monocytes and Macrophages as Viral Targets and Reservoirs. Int. J. Mol. Sci. 2018, 19, 2821. [Google Scholar] [CrossRef]
- Lenac Rovis, T.; Kucan Brlic, P.; Kaynan, N.; Juranic Lisnic, V.; Brizic, I.; Jordan, S.; Tomic, A.; Kvestak, D.; Babic, M.; Tsukerman, P.; et al. Inflammatory monocytes and NK cells play a crucial role in DNAM-1-dependent control of cytomegalovirus infection. J. Exp. Med. 2016, 213, 1835–1850. [Google Scholar] [CrossRef]
- Carmichael, A. Cytomegalovirus and the eye. Eye 2012, 26, 237–240. [Google Scholar] [CrossRef]
- Keino, H.; Horie, S.; Sugita, S. Immune Privilege and Eye-Derived T-Regulatory Cells. Immunol. Res. 2018, 1679197. [Google Scholar] [CrossRef] [PubMed]
- Nagineni, C.N.; Pardhasaradhi, K.; Martins, M.C.; Detrick, B.; Hooks, J.J. Mechanisms of interferon-induced inhibition of Toxoplasma gondii replication in human retinal pigment epithelial cells. Infect. Immun. 1996, 64, 4188–4196. [Google Scholar] [PubMed]
- Bodaghi, B.; Goureau, O.; Zipeto, D.; Laurent, L.; Virelizier, J.L.; Michelson, S. Role of IFN-gamma-induced indoleamine 2,3 dioxygenase and inducible nitric oxide synthase in the replication of human cytomegalovirus in retinal pigment epithelial cells. J. Immunol. 1999, 162, 957–964. [Google Scholar] [PubMed]
- Däubener, W.; Schmidt, S.K.; Heseler, K.; Spekker, K.H.; MacKenzie, C.R. Antimicrobial and immunoregulatory effector mechanisms in human endothelial cells. Indoleamine 2,3-dioxygenase versus inducible nitric oxide synthase. Thromb. Haemost. 2009, 102, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Heseler, K.; Schmidt, S.K.; Spekker, K.; Sinzger, C.; Sorg, R.V.; Quambusch, M.; Zimmermann, A.; Meisel, R.; Däubener, W. Cytomegalovirus impairs the induction of indoleamine 2,3-dioxygenase mediated antimicrobial and immunoregulatory effects in human fibroblasts. PLoS ONE 2013, 8, e64442. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, A.; Hauka, S.; Maywald, M.; Le, V.T.; Schmidt, S.K.; Däubener, W.; Hengel, H. Checks and balances between human cytomegalovirus replication and indoleamine-2,3-dioxygenase. J. Gen. Virol. 2014, 95, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Pleyer, U.; Schlüter, D.; Mänz, M. Ocular toxoplasmosis: Recent aspects of pathophysiology and clinical implications. Ophthalmic Res. 2014, 52, 116–123. [Google Scholar] [CrossRef]
- Schlüter, D.; Däubener, W.; Schares, G.; Groß, U.; Pleyer, U.; Lüder, C. Animals are key to human toxoplasmosis. Int. J. Med. Microbiol. 2014, 304, 917–929. [Google Scholar] [CrossRef]
- Baron, M.; Davignon, J.L. Inhibition of IFN-gamma-induced STAT1 tyrosine phosphorylation by human CMV is mediated by SHP2. J. Immunol. 2008, 181, 5530–5536. [Google Scholar] [CrossRef]
- Miller, D.M.; Rahill, B.M.; Boss, J.M.; Lairmore, M.D.; Durbin, J.E.; Waldman, J.W.; Sedmak, D.D. Human cytomegalovirus inhibits major histocompatibility complex class II expression by disruption of the Jak/Stat pathway. J. Exp. Med. 1998, 187, 675–683. [Google Scholar] [CrossRef]
- Gregerson, D.S.; Heuss, N.D.; Lew, K.L.; McPherson, S.W.; Ferrington, D.A. Interaction of retinal pigmented epithelial cells and CD4 T cells leads to T-cell anergy. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4654–4663. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S.; Kawazoe, Y.; Imai, A.; Usui, Y.; Takahashi, M.; Mochizuki, M. Suppression of IL-22-producing T helper 22 cells by RPE cells via PD-L1/PD-1 interactions. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6926–6933. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Yang, S.H.; Chuck, R.S.; Gehlbach, P.L.; Park, C.G. The role of indoleamine 2,3-dioxygenase in retinal pigment epithelial cell-mediated immune modulation. Ocul. Immunol. Inflamm. 2010, 18, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Kaestel, C.G.; Lovato, P.; Ødum, N.; Nissen, M.H.; Röpke, C. The immune privilege of the eye: Human retinal pigment epithelial cells selectively modulate T-cell activation in vitro. Curr. Eye Res. 2005, 30, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Munn, D.H.; Shafizadeh, E.; Attwood, J.T.; Bondarev, I.; Pashine, A.; Mellor, A.L. Inhibition of T cell proliferation by macrophage tryptophan catabolism. J. Exp. Med. 1999, 189, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Däubener, W.; Hucke, C.; Seidel, K.; Hadding, U.; MacKenzie, C.R. Interleukin-1 inhibits gamma interferon-induced bacteriostasis in human uroepithelial cells. Infect. Immun. 1999, 67, 5615–5620. [Google Scholar] [PubMed]
- Hucke, C.; MacKenzie, C.R.; Adjogble, K.D.; Takikawa, O.; Däubener, W. Nitric oxide-mediated regulation of gamma interferon-induced bacteriostasis: Inhibition and degradation of human indoleamine 2,3-dioxygenase. Infect. Immun. 2004, 72, 2723–2730. [Google Scholar] [CrossRef]
- Caspi, R.R. A look at autoimmunity and inflammation in the eye. J. Clin. Investig. 2010, 120, 3073–3083. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Agarwal, A.; Mahendradas, P.; Lee, C.S.; Gupta, V.; Pavesio, C.E.; Agrawal, R. Viral posterior uveitis. Surv. Ophthalmol. 2017, 62, 404–445. [Google Scholar] [CrossRef]
- Däubener, W.; Wanagat, N.; Pilz, K.; Seghrouchni, S.; Fischer, H.G.; Hadding, U. A new, simple bioassay for human IFN-gamma. J. Immunol. Methods 1994, 168, 39–47. [Google Scholar] [CrossRef]
- Ding, A.H.; Nathan, C.F.; Stuehr, D.J. Release of reactive nitrogen intermediates and reactive oxygen intermediates from mouse peritoneal macrophages. Comparison of activating cytokines and evidence for independent production. J. Immunol. 1988, 141, 2407–2412. [Google Scholar] [PubMed]
- Sinzger, C.; Schmidt, K.; Knapp, J.; Kahl, M.; Beck, R.; Waldman, J.; Hebart, H.; Einsele, H.; Jahn, G. Modification of human cytomegalovirus tropism through propagation in vitro is associated with changes in the viral genome. J. Gen. Virol. 1999, 80, 2867–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinzger, C.; Hahn, G.; Digel, M.; Katona, R.; Sampaio, K.L.; Messerle, M.; Hengel, H.; Koszinowski, U.; Brune, W.; Adler, B. Cloning and sequencing of a highly productive, endotheliotropic virus strain derived from human cytomegalovirus TB40/E. J. Gen. Virol. 2008, 89, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, V.T.; Trilling, M.; Wilborn, M.; Hengel, H.; Zimmermann, A. Human cytomegalovirus interferes with signal transducer and activator of transcription (STAT) 2 protein stability and tyrosine phosphorylation. J. Gen. Virol. 2008, 89, 2416–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfefferkorn, E.R.; Pfefferkorn, L.C. Specific labeling of intracellular Toxoplasma gondii with uracil. J. Protozool. 1977, 24, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Heseler, K.; Spekker, K.; Schmidt, S.K.; MacKenzie, C.R.; Däubener, W. Antimicrobial and immunoregulatory effects mediated by human lung cells: Role of IFN-gamma-induced tryptophan degradation. FEMS Immunol. Med. Microbiol. 2008, 52, 273–281. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spekker-Bosker, K.; Ufermann, C.-M.; Maywald, M.; Zimmermann, A.; Domröse, A.; Woite, C.; Däubener, W.; Eller, S.K. hCMV-Mediated Immune Escape Mechanisms Favor Pathogen Growth and Disturb the Immune Privilege of the Eye. Int. J. Mol. Sci. 2019, 20, 858. https://doi.org/10.3390/ijms20040858
Spekker-Bosker K, Ufermann C-M, Maywald M, Zimmermann A, Domröse A, Woite C, Däubener W, Eller SK. hCMV-Mediated Immune Escape Mechanisms Favor Pathogen Growth and Disturb the Immune Privilege of the Eye. International Journal of Molecular Sciences. 2019; 20(4):858. https://doi.org/10.3390/ijms20040858
Chicago/Turabian StyleSpekker-Bosker, Katrin, Christoph-Martin Ufermann, Marco Maywald, Albert Zimmermann, Andreas Domröse, Claudia Woite, Walter Däubener, and Silvia Kathrin Eller. 2019. "hCMV-Mediated Immune Escape Mechanisms Favor Pathogen Growth and Disturb the Immune Privilege of the Eye" International Journal of Molecular Sciences 20, no. 4: 858. https://doi.org/10.3390/ijms20040858
APA StyleSpekker-Bosker, K., Ufermann, C. -M., Maywald, M., Zimmermann, A., Domröse, A., Woite, C., Däubener, W., & Eller, S. K. (2019). hCMV-Mediated Immune Escape Mechanisms Favor Pathogen Growth and Disturb the Immune Privilege of the Eye. International Journal of Molecular Sciences, 20(4), 858. https://doi.org/10.3390/ijms20040858