Telomerase-Targeted Cancer Immunotherapy

Abstract
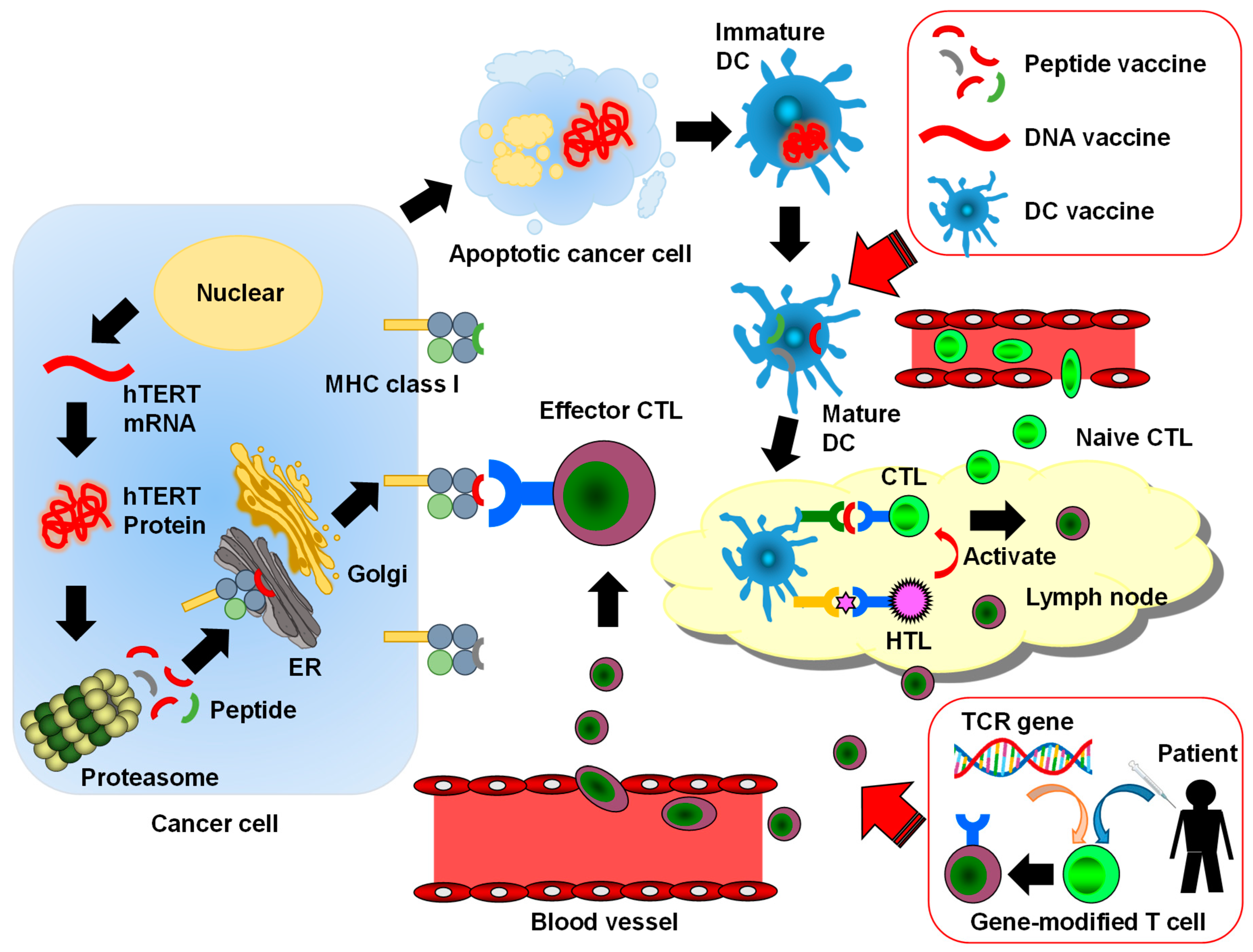
:1. Introduction
2. Recent Advancements in Cancer Immunotherapy
3. Expression of hTERT as a Target Antigen in Cancer Immunotherapy
4. Development of Peptide Vaccines That Target hTERT
4.1. GV1001
4.2. GX301
4.3. UV1
4.4. Vx-001
5. Immunotherapy Using hTERT-Targeting Dendritic Cells (DCs)
5.1. GRNVAC1
5.2. TAPCells
5.3. Other DC-Based Approaches
6. DNA Vaccines
6.1. phTERT
6.2. INVAC-1
7. Cell-Based Immunological Approaches
8. Gene-Modified T-Cell Therapy
9. hTERT-Targeted Cancer Immunotherapy: Future Perspectives
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| hTERT | human telomerase reverse transcriptase |
| CAR | chimeric antigen receptor |
| CTL | cytotoxic T lymphocyte |
| DC | dendritic cell |
| TAA | tumor associated antigen |
| TCR | T-cell receptor |
| HCC | hepatocellular carcinoma |
| HSP | heat shock protein |
| IFN | interferon |
References
- Maciejowski, J.; de Lange, T. Telomeres in cancer: Tumour suppression and genome instability. Nat. Rev. Mol. Cell Biol. 2017, 18, 175–186. [Google Scholar] [CrossRef]
- Okamoto, K.; Seimiya, H. Revisiting Telomere Shortening in Cancer. Cells 2019, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Ivancich, M.; Schrank, Z.; Wojdyla, L.; Leviskas, B.; Kuckovic, A.; Sanjali, A.; Puri, N. Treating Cancer by Targeting Telomeres and Telomerase. Antioxidants 2017, 6. [Google Scholar] [CrossRef]
- Martinez, P.; Blasco, M.A. Telomere-driven diseases and telomere-targeting therapies. J. Cell Biol. 2017, 216, 875–887. [Google Scholar] [CrossRef] [Green Version]
- Carrozza, F.; Santoni, M.; Piva, F.; Cheng, L.; Lopez-Beltran, A.; Scarpelli, M.; Montironi, R.; Battelli, N.; Tamberi, S. Emerging immunotherapeutic strategies targeting telomerases in genitourinary tumors. Crit. Rev. Oncol. Hematol. 2018, 131, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Eitsuka, T.; Nakagawa, K.; Kato, S.; Ito, J.; Otoki, Y.; Takasu, S.; Shimizu, N.; Takahashi, T.; Miyazawa, T. Modulation of Telomerase Activity in Cancer Cells by Dietary Compounds: A Review. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. TERT promoter mutations in familial and sporadic melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef]
- Stogbauer, L.; Stummer, W.; Senner, V.; Brokinkel, B. Telomerase activity, TERT expression, hTERT promoter alterations, and alternative lengthening of the telomeres (ALT) in meningiomas—A systematic review. Neurosurg. Rev. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, M. A second chance for telomerase reverse transcriptase in anticancer immunotherapy. Nat. Rev. Clin. Oncol. 2017, 14, 115–128. [Google Scholar] [CrossRef]
- Wolchok, J.D.; Chiarion-Sileni, V.; Gonzalez, R.; Rutkowski, P.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Wagstaff, J.; Schadendorf, D.; Ferrucci, P.F.; et al. Overall Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. New Engl. J. Med. 2017, 377, 1345–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motzer, R.J.; Tannir, N.M.; McDermott, D.F.; Aren Frontera, O.; Melichar, B.; Choueiri, T.K.; Plimack, E.R.; Barthelemy, P.; Porta, C.; George, S.; et al. Nivolumab plus Ipilimumab versus Sunitinib in Advanced Renal-Cell Carcinoma. New Engl. J. Med. 2018, 378, 1277–1290. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Riviere, I.; Gonen, M.; Wang, X.; Senechal, B.; Curran, K.J.; Sauter, C.; Wang, Y.; Santomasso, B.; Mead, E.; et al. Long-Term Follow-up of CD19 CAR Therapy in Acute Lymphoblastic Leukemia. N. Engl. J Med. 2018, 378, 449–459. [Google Scholar] [CrossRef]
- Maude, S.L.; Laetsch, T.W.; Buechner, J.; Rives, S.; Boyer, M.; Bittencourt, H.; Bader, P.; Verneris, M.R.; Stefanski, H.E.; Myers, G.D.; et al. Tisagenlecleucel in Children and Young Adults with B-Cell Lymphoblastic Leukemia. N. Engl. J. Med. 2018, 378, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Farhood, B.; Najafi, M.; Mortezaee, K. CD8(+) cytotoxic T lymphocytes in cancer immunotherapy: A review. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef]
- Durgeau, A.; Virk, Y.; Corgnac, S.; Mami-Chouaib, F. Recent Advances in Targeting CD8 T-Cell Immunity for More Effective Cancer Immunotherapy. Front. Immunol. 2018, 9, 14. [Google Scholar] [CrossRef]
- Schooten, E.; Di Maggio, A.; van Bergen En Henegouwen, P.M.P.; Kijanka, M.M. MAGE-A antigens as targets for cancer immunotherapy. Cancer Treat. Rev. 2018, 67, 54–62. [Google Scholar] [CrossRef]
- Thomas, R.; Al-Khadairi, G.; Roelands, J.; Hendrickx, W.; Dermime, S.; Bedognetti, D.; Decock, J. NY-ESO-1 Based Immunotherapy of Cancer: Current Perspectives. Front. Immunol. 2018, 9, 947. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Mellman, I. Oncology meets immunology: The cancer-immunity cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rauch, J.; Gires, O. SEREX, Proteomex, AMIDA, and beyond: Serological screening technologies for target identification. Proteom. Clin. Appl. 2008, 2, 355–371. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Tomita, Y.; Yuno, A.; Yoshitake, Y.; Shinohara, M. Cancer immunotherapy using novel tumor-associated antigenic peptides identified by genome-wide cDNA microarray analyses. Cancer Sci. 2015, 106, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.P.; Chen, W.; Schwarer, A.P.; Li, H. Telomerase in cancer immunotherapy. Biochim. Biophys. Acta 2010, 1805, 35–42. [Google Scholar] [CrossRef]
- Nakamura, T.M.; Morin, G.B.; Chapman, K.B.; Weinrich, S.L.; Andrews, W.H.; Lingner, J.; Harley, C.B.; Cech, T.R. Telomerase catalytic subunit homologs from fission yeast and human. Science 1997, 277, 955–959. [Google Scholar] [CrossRef]
- Shay, J.W.; Bacchetti, S. A survey of telomerase activity in human cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Hannen, R.; Bartsch, J.W. Essential roles of telomerase reverse transcriptase hTERT in cancer stemness and metastasis. FEBS Lett. 2018, 592, 2023–2031. [Google Scholar] [CrossRef] [Green Version]
- Dogan, F.; Biray Avci, C. Correlation between telomerase and mTOR pathway in cancer stem cells. Gene 2018, 641, 235–239. [Google Scholar] [CrossRef]
- Vonderheide, R.H.; Hahn, W.C.; Schultze, J.L.; Nadler, L.M. The telomerase catalytic subunit is a widely expressed tumor-associated antigen recognized by cytotoxic T lymphocytes. Immunity 1999, 10, 673–679. [Google Scholar] [CrossRef]
- Lev, A.; Denkberg, G.; Cohen, C.J.; Tzukerman, M.; Skorecki, K.L.; Chames, P.; Hoogenboom, H.R.; Reiter, Y. Isolation and characterization of human recombinant antibodies endowed with the antigen-specific, major histocompatibility complex-restricted specificity of T cells directed toward the widely expressed tumor T-cell epitopes of the telomerase catalytic subunit. Cancer Res. 2002, 62, 3184–3194. [Google Scholar]
- Kumagai, M.; Mizukoshi, E.; Tamai, T.; Kitahara, M.; Yamashita, T.; Arai, K.; Terashima, T.; Iida, N.; Fushimi, K.; Kaneko, S. Immune response to human telomerase reverse transcriptase-derived helper T cell epitopes in hepatocellular carcinoma patients. Liver Int. 2018, 38, 1635–1645. [Google Scholar] [CrossRef]
- Minev, B.; Hipp, J.; Firat, H.; Schmidt, J.D.; Langlade-Demoyen, P.; Zanetti, M. Cytotoxic T cell immunity against telomerase reverse transcriptase in humans. Proc. Natl. Acad. Sci. USA 2000, 97, 4796–4801. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, J.; Garcia-Pons, F.; Lone, Y.C.; Firat, H.; Schmidt, J.D.; Langlade-Demoyen, P.; Zanetti, M. Identification of a human telomerase reverse transcriptase peptide of low affinity for HLA A2.1 that induces cytotoxic T lymphocytes and mediates lysis of tumor cells. Proc. Natl. Acad. Sci. USA 2002, 99, 12275–12280. [Google Scholar] [CrossRef] [Green Version]
- Scardino, A.; Gross, D.A.; Alves, P.; Schultze, J.L.; Graff-Dubois, S.; Faure, O.; Tourdot, S.; Chouaib, S.; Nadler, L.M.; Lemonnier, F.A.; et al. HER-2/neu and hTERT cryptic epitopes as novel targets for broad spectrum tumor immunotherapy. J. Immunol. 2002, 168, 5900–5906. [Google Scholar] [CrossRef]
- Thorn, M.; Wang, M.; Kloverpris, H.; Schmidt, E.G.; Fomsgaard, A.; Wenandy, L.; Berntsen, A.; Brunak, S.; Buus, S.; Claesson, M.H. Identification of a new hTERT-derived HLA-A*0201 restricted, naturally processed CTL epitope. Cancer Immunol. Immunother. 2007, 56, 1755–1763. [Google Scholar] [CrossRef]
- Schreurs, M.W.; Hermsen, M.A.; Geltink, R.I.; Scholten, K.B.; Brink, A.A.; Kueter, E.W.; Tijssen, M.; Meijer, C.J.; Ylstra, B.; Meijer, G.A.; et al. Genomic stability and functional activity may be lost in telomerase-transduced human CD8+ T lymphocytes. Blood 2005, 106, 2663–2670. [Google Scholar] [CrossRef] [Green Version]
- Mizukoshi, E.; Nakamoto, Y.; Marukawa, Y.; Arai, K.; Yamashita, T.; Tsuji, H.; Kuzushima, K.; Takiguchi, M.; Kaneko, S. Cytotoxic T cell responses to human telomerase reverse transcriptase in patients with hepatocellular carcinoma. Hepatology 2006, 43, 1284–1294. [Google Scholar] [CrossRef] [Green Version]
- Adotevi, O.; Mollier, K.; Neuveut, C.; Cardinaud, S.; Boulanger, E.; Mignen, B.; Fridman, W.H.; Zanetti, M.; Charneau, P.; Tartour, E.; et al. Immunogenic HLA-B*0702-restricted epitopes derived from human telomerase reverse transcriptase that elicit antitumor cytotoxic T-cell responses. Clin. Cancer Res. 2006, 12, 3158–3167. [Google Scholar] [CrossRef]
- Cortez-Gonzalez, X.; Sidney, J.; Adotevi, O.; Sette, A.; Millard, F.; Lemonnier, F.; Langlade-Demoyen, P.; Zanetti, M. Immunogenic HLA-B7-restricted peptides of hTRT. Int. Immunol. 2006, 18, 1707–1718. [Google Scholar] [CrossRef] [Green Version]
- Bernardeau, K.; Kerzhero, J.; Fortun, A.; Moreau-Aubry, A.; Favry, E.; Echasserieau, K.; Tartour, E.; Maillere, B.; Lang, F. A simple competitive assay to determine peptide affinity for HLA class II molecules: A useful tool for epitope prediction. J. Immunol. Methods 2011, 371, 97–105. [Google Scholar] [CrossRef]
- Suso, E.M.; Dueland, S.; Rasmussen, A.M.; Vetrhus, T.; Aamdal, S.; Kvalheim, G.; Gaudernack, G. hTERT mRNA dendritic cell vaccination: Complete response in a pancreatic cancer patient associated with response against several hTERT epitopes. Cancer Immunol. Immunother. 2011, 60, 809–818. [Google Scholar] [CrossRef]
- Wang, J.; Yu, L.; Li, J.; Deng, R.; Wang, X. Characterization of a human telomerase reverse transcriptase sequence containing two antigenic epitopes with high affinity for human leucocyte antigen. Biotechnol. Appl. Biochem. 2007, 48, 93–99. [Google Scholar]
- Vonderheide, R.H.; Anderson, K.S.; Hahn, W.C.; Butler, M.O.; Schultze, J.L.; Nadler, L.M. Characterization of HLA-A3-restricted cytotoxic T lymphocytes reactive against the widely expressed tumor antigen telomerase. Clin. Cancer Res. 2001, 7, 3343–3348. [Google Scholar]
- Schroers, R.; Shen, L.; Rollins, L.; Rooney, C.M.; Slawin, K.; Sonderstrup, G.; Huang, X.F.; Chen, S.Y. Human telomerase reverse transcriptase-specific T-helper responses induced by promiscuous major histocompatibility complex class II-restricted epitopes. Clin. Cancer Res. 2003, 9, 4743–4755. [Google Scholar]
- Schroers, R.; Huang, X.F.; Hammer, J.; Zhang, J.; Chen, S.Y. Identification of HLA DR7-restricted epitopes from human telomerase reverse transcriptase recognized by CD4+ T-helper cells. Cancer Res. 2002, 62, 2600–2605. [Google Scholar]
- Huang, G.; Geng, J.; Wang, R.; Chen, L. Identification of a new cytotoxic T-cell epitope p675 of human telomerase reverse transcriptase. Cancer Biother. Radiopharm. 2012, 27, 600–605. [Google Scholar] [CrossRef]
- Vonderheide, R.H. Telomerase as a universal tumor-associated antigen for cancer immunotherapy. Oncogene 2002, 21, 674–679. [Google Scholar] [CrossRef] [Green Version]
- Vonderheide, R.H. Prospects and challenges of building a cancer vaccine targeting telomerase. Biochimie 2008, 90, 173–180. [Google Scholar] [CrossRef]
- Kailashiya, C.; Sharma, H.B.; Kailashiya, J. Telomerase based anticancer immunotherapy and vaccines approaches. Vaccine 2017, 35, 5768–5775. [Google Scholar] [CrossRef]
- Xu, Y.; Goldkorn, A. Telomere and Telomerase Therapeutics in Cancer. Genes 2016, 7, 22. [Google Scholar] [CrossRef]
- Bolonaki, I.; Kotsakis, A.; Papadimitraki, E.; Aggouraki, D.; Konsolakis, G.; Vagia, A.; Christophylakis, C.; Nikoloudi, I.; Magganas, E.; Galanis, A.; et al. Vaccination of patients with advanced non-small-cell lung cancer with an optimized cryptic human telomerase reverse transcriptase peptide. J. Clin. Oncol. 2007, 25, 2727–2734. [Google Scholar] [CrossRef]
- Kotsakis, A.; Vetsika, E.K.; Christou, S.; Hatzidaki, D.; Vardakis, N.; Aggouraki, D.; Konsolakis, G.; Georgoulias, V.; Christophyllakis, C.; Cordopatis, P.; et al. Clinical outcome of patients with various advanced cancer types vaccinated with an optimized cryptic human telomerase reverse transcriptase (TERT) peptide: Results of an expanded phase II study. Ann. Oncol. 2012, 23, 442–449. [Google Scholar] [CrossRef]
- Vetsika, E.K.; Konsolakis, G.; Aggouraki, D.; Kotsakis, A.; Papadimitraki, E.; Christou, S.; Menez-Jamet, J.; Kosmatopoulos, K.; Georgoulias, V.; Mavroudis, D. Immunological responses in cancer patients after vaccination with the therapeutic telomerase-specific vaccine Vx-001. Cancer Immunol. Immunother. 2012, 61, 157–168. [Google Scholar] [CrossRef]
- Greten, T.F.; Forner, A.; Korangy, F.; N’Kontchou, G.; Barget, N.; Ayuso, C.; Ormandy, L.A.; Manns, M.P.; Beaugrand, M.; Bruix, J. A phase II open label trial evaluating safety and efficacy of a telomerase peptide vaccination in patients with advanced hepatocellular carcinoma. BMC Cancer 2010, 10, 209. [Google Scholar] [CrossRef]
- Kyte, J.A.; Gaudernack, G.; Dueland, S.; Trachsel, S.; Julsrud, L.; Aamdal, S. Telomerase peptide vaccination combined with temozolomide: A clinical trial in stage IV melanoma patients. Clin. Cancer Res. 2011, 17, 4568–4580. [Google Scholar] [CrossRef]
- Inderberg-Suso, E.M.; Trachsel, S.; Lislerud, K.; Rasmussen, A.M.; Gaudernack, G. Widespread CD4+ T-cell reactivity to novel hTERT epitopes following vaccination of cancer patients with a single hTERT peptide GV1001. Oncoimmunology 2012, 1, 670–686. [Google Scholar] [CrossRef] [Green Version]
- Staff, C.; Mozaffari, F.; Frodin, J.E.; Mellstedt, H.; Liljefors, M. Telomerase (GV1001) vaccination together with gemcitabine in advanced pancreatic cancer patients. Int. J. Oncol. 2014, 45, 1293–1303. [Google Scholar] [CrossRef]
- Middleton, G.; Silcocks, P.; Cox, T.; Valle, J.; Wadsley, J.; Propper, D.; Coxon, F.; Ross, P.; Madhusudan, S.; Roques, T.; et al. Gemcitabine and capecitabine with or without telomerase peptide vaccine GV1001 in patients with locally advanced or metastatic pancreatic cancer (TeloVac): An open-label, randomised, phase 3 trial. Lancet Oncol. 2014, 15, 829–840. [Google Scholar] [CrossRef]
- Brunsvig, P.F.; Aamdal, S.; Gjertsen, M.K.; Kvalheim, G.; Markowski-Grimsrud, C.J.; Sve, I.; Dyrhaug, M.; Trachsel, S.; Moller, M.; Eriksen, J.A.; et al. Telomerase peptide vaccination: A phase I/II study in patients with non-small cell lung cancer. Cancer Immunol. Immunother. 2006, 55, 1553–1564. [Google Scholar] [CrossRef]
- Hunger, R.E.; Kernland Lang, K.; Markowski, C.J.; Trachsel, S.; Moller, M.; Eriksen, J.A.; Rasmussen, A.M.; Braathen, L.R.; Gaudernack, G. Vaccination of patients with cutaneous melanoma with telomerase-specific peptides. Cancer Immunol. Immunother. 2011, 60, 1553–1564. [Google Scholar] [CrossRef]
- Lilleby, W.; Gaudernack, G.; Brunsvig, P.F.; Vlatkovic, L.; Schulz, M.; Mills, K.; Hole, K.H.; Inderberg, E.M. Phase I/IIa clinical trial of a novel hTERT peptide vaccine in men with metastatic hormone-naive prostate cancer. Cancer Immunol. Immunother. 2017, 66, 891–901. [Google Scholar] [CrossRef]
- Kotsakis, A.; Papadimitraki, E.; Vetsika, E.K.; Aggouraki, D.; Dermitzaki, E.K.; Hatzidaki, D.; Kentepozidis, N.; Mavroudis, D.; Georgoulias, V. A phase II trial evaluating the clinical and immunologic response of HLA-A2(+) non-small cell lung cancer patients vaccinated with an hTERT cryptic peptide. Lung Cancer 2014, 86, 59–66. [Google Scholar] [CrossRef]
- Fenoglio, D.; Traverso, P.; Parodi, A.; Tomasello, L.; Negrini, S.; Kalli, F.; Battaglia, F.; Ferrera, F.; Sciallero, S.; Murdaca, G.; et al. A multi-peptide, dual-adjuvant telomerase vaccine (GX301) is highly immunogenic in patients with prostate and renal cancer. Cancer Immunol. Immunother. 2013, 62, 1041–1052. [Google Scholar] [CrossRef]
- Mizukoshi, E.; Nakagawa, H.; Kitahara, M.; Yamashita, T.; Arai, K.; Sunagozaka, H.; Fushimi, K.; Kobayashi, E.; Kishi, H.; Muraguchi, A.; et al. Immunological features of T cells induced by human telomerase reverse transcriptase-derived peptides in patients with hepatocellular carcinoma. Cancer Lett. 2015, 364, 98–105. [Google Scholar] [CrossRef]
- Vonderheide, R.H.; Domchek, S.M.; Schultze, J.L.; George, D.J.; Hoar, K.M.; Chen, D.Y.; Stephans, K.F.; Masutomi, K.; Loda, M.; Xia, Z.; et al. Vaccination of cancer patients against telomerase induces functional antitumor CD8+ T lymphocytes. Clin. Cancer Res. 2004, 10, 828–839. [Google Scholar] [CrossRef]
- Su, Z.; Dannull, J.; Yang, B.K.; Dahm, P.; Coleman, D.; Yancey, D.; Sichi, S.; Niedzwiecki, D.; Boczkowski, D.; Gilboa, E.; et al. Telomerase mRNA-transfected dendritic cells stimulate antigen-specific CD8+ and CD4+ T cell responses in patients with metastatic prostate cancer. J. Immunol. 2005, 174, 3798–3807. [Google Scholar] [CrossRef]
- Aloysius, M.M.; Mc Kechnie, A.J.; Robins, R.A.; Verma, C.; Eremin, J.M.; Farzaneh, F.; Habib, N.A.; Bhalla, J.; Hardwick, N.R.; Satthaporn, S.; et al. Generation in vivo of peptide-specific cytotoxic T cells and presence of regulatory T cells during vaccination with hTERT (class I and II) peptide-pulsed DCs. J. Transl. Med. 2009, 7, 18. [Google Scholar] [CrossRef]
- Khoury, H.J.; Collins, R.H., Jr.; Blum, W.; Stiff, P.S.; Elias, L.; Lebkowski, J.S.; Reddy, A.; Nishimoto, K.P.; Sen, D.; Wirth, E.D., 3rd; et al. Immune responses and long-term disease recurrence status after telomerase-based dendritic cell immunotherapy in patients with acute myeloid leukemia. Cancer 2017, 123, 3061–3072. [Google Scholar] [CrossRef]
- Salazar-Onfray, F.; Pereda, C.; Reyes, D.; Lopez, M.N. TAPCells, the Chilean dendritic cell vaccine against melanoma and prostate cancer. Biol. Res. 2013, 46, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Mehrotra, S.; Britten, C.D.; Chin, S.; Garrett-Mayer, E.; Cloud, C.A.; Li, M.; Scurti, G.; Salem, M.L.; Nelson, M.H.; Thomas, M.B.; et al. Vaccination with poly(IC:LC) and peptide-pulsed autologous dendritic cells in patients with pancreatic cancer. J. Hematol. Oncol. 2017, 10, 82. [Google Scholar] [CrossRef]
- Jafri, M.A.; Ansari, S.A.; Alqahtani, M.H.; Shay, J.W. Roles of telomeres and telomerase in cancer, and advances in telomerase-targeted therapies. Genome Med. 2016, 8, 69. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.K.; Kim, B.R.; Lee, H.J.; Lee, S.A.; Kim, B.J.; Kim, H.; Won, Y.S.; Shon, W.J.; Lee, N.R.; Inn, K.S. Tumor-suppressive effect of a telomerase-derived peptide by inhibiting hypoxia-induced HIF-1alpha-VEGF signaling axis. Biomaterials 2014, 35, 2924–2933. [Google Scholar] [CrossRef]
- Kim, H.; Seo, E.H.; Lee, S.H.; Kim, B.J. The Telomerase-Derived Anticancer Peptide Vaccine GV1001 as an Extracellular Heat Shock Protein-Mediated Cell-Penetrating Peptide. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef]
- Kim, G.E.; Jung, A.R.; Kim, M.Y.; Lee, J.B.; Im, J.H.; Lee, K.W.; Park, Y.H.; Lee, J.Y. GV1001 Induces Apoptosis by Reducing Angiogenesis in Renal Cell Carcinoma Cells Both In Vitro and In Vivo. Urology 2018, 113, 129–137. [Google Scholar] [CrossRef]
- Chang, J.E.; Kim, H.J.; Yi, E.; Jheon, S.; Kim, K. Reduction of ischaemia-reperfusion injury in a rat lung transplantation model by low-concentration GV1001. Eur. J. Cardiothorac. Surg. 2016, 50, 972–979. [Google Scholar] [CrossRef]
- Choi, J.; Kim, H.; Kim, Y.; Jang, M.; Jeon, J.; Hwang, Y.I.; Shon, W.J.; Song, Y.W.; Kang, J.S.; Lee, W.J. The Anti-inflammatory Effect of GV1001 Mediated by the Downregulation of ENO1-induced Pro-inflammatory Cytokine Production. Immune Netw. 2015, 15, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Ko, Y.J.; Kwon, K.Y.; Kum, K.Y.; Lee, W.C.; Baek, S.H.; Kang, M.K.; Shon, W.J. The Anti-Inflammatory Effect of Human Telomerase-Derived Peptide on P. gingivalis Lipopolysaccharide-Induced Inflammatory Cytokine Production and Its Mechanism in Human Dental Pulp Cells. Mediat. Inflamm. 2015, 2015, 385127. [Google Scholar] [CrossRef]
- Kim, H.; Choi, M.S.; Inn, K.S.; Kim, B.J. Inhibition of HIV-1 reactivation by a telomerase-derived peptide in a HSP90-dependent manner. Sci. Rep. 2016, 6, 28896. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.A.; Kim, J.; Sim, J.; Kim, S.G.; Kook, Y.H.; Park, C.G.; Kim, H.R.; Kim, B.J. A telomerase-derived peptide regulates reactive oxygen species and hepatitis C virus RNA replication in HCV-infected cells via heat shock protein 90. Biochem. Biophys. Res. Commun. 2016, 471, 156–162. [Google Scholar] [CrossRef]
- Park, H.H.; Lee, K.Y.; Kim, S.; Lee, J.W.; Choi, N.Y.; Lee, E.H.; Lee, Y.J.; Lee, S.H.; Koh, S.H. Novel vaccine peptide GV1001 effectively blocks beta-amyloid toxicity by mimicking the extra-telomeric functions of human telomerase reverse transcriptase. Neurobiol. Aging 2014, 35, 1255–1274. [Google Scholar] [CrossRef]
- Brower, V. Telomerase-based therapies emerging slowly. J. Natl. Cancer Inst. 2010, 102, 520–521. [Google Scholar] [CrossRef]
- Park, J.K.; Kim, Y.; Kim, H.; Jeon, J.; Kim, T.W.; Park, J.H.; Hwnag, Y.I.; Lee, W.J.; Kang, J.S. The anti-fibrotic effect of GV1001 combined with gemcitabine on treatment of pancreatic ductal adenocarcinoma. Oncotarget 2016, 7, 75081–75093. [Google Scholar] [CrossRef] [Green Version]
- Middleton, G.; Greenhalf, W.; Costello, E.; Shaw, V.; Cox, T.; Ghaneh, P.; Palmer, D.H.; Neoptolemos, J.P. Immunobiological effects of gemcitabine and capecitabine combination chemotherapy in advanced pancreatic ductal adenocarcinoma. Br J Cancer 2016, 114, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Fenoglio, D.; Parodi, A.; Lavieri, R.; Kalli, F.; Ferrera, F.; Tagliamacco, A.; Guastalla, A.; Lamperti, M.G.; Giacomini, M.; Filaci, G. Immunogenicity of GX301 cancer vaccine: Four (telomerase peptides) are better than one. Hum. Vaccin. Immunother. 2015, 11, 838–850. [Google Scholar] [CrossRef] [Green Version]
- Ruden, M.; Puri, N. Novel anticancer therapeutics targeting telomerase. Cancer Treat. Rev. 2013, 39, 444–456. [Google Scholar] [CrossRef]
- Menez-Jamet, J.; Gallou, C.; Rougeot, A.; Kosmatopoulos, K. Optimized tumor cryptic peptides: The basis for universal neo-antigen-like tumor vaccines. Ann. Transl. Med. 2016, 4, 266. [Google Scholar] [CrossRef]
- Galati, D.; Zanotta, S. Empowering dendritic cell cancer vaccination: The role of combinatorial strategies. Cytotherapy 2018, 20, 1309–1323. [Google Scholar] [CrossRef]
- Kantoff, P.W.; Higano, C.S.; Shore, N.D.; Berger, E.R.; Small, E.J.; Penson, D.F.; Redfern, C.H.; Ferrari, A.C.; Dreicer, R.; Sims, R.B.; et al. Sipuleucel-T immunotherapy for castration-resistant prostate cancer. New Engl. J. Med. 2010, 363, 411–422. [Google Scholar] [CrossRef]
- Nair, S.K.; Heiser, A.; Boczkowski, D.; Majumdar, A.; Naoe, M.; Lebkowski, J.S.; Vieweg, J.; Gilboa, E. Induction of cytotoxic T cell responses and tumor immunity against unrelated tumors using telomerase reverse transcriptase RNA transfected dendritic cells. Nat. Med. 2000, 6, 1011–1017. [Google Scholar] [CrossRef]
- Sioud, M.; Nyakas, M.; Saeboe-Larssen, S.; Mobergslien, A.; Aamdal, S.; Kvalheim, G. Diversification of Antitumour Immunity in a Patient with Metastatic Melanoma Treated with Ipilimumab and an IDO-Silenced Dendritic Cell Vaccine. Case Rep. Med. 2016, 2016, 9639585. [Google Scholar] [CrossRef]
- Frolkis, M.; Fischer, M.B.; Wang, Z.; Lebkowski, J.S.; Chiu, C.P.; Majumdar, A.S. Dendritic cells reconstituted with human telomerase gene induce potent cytotoxic T-cell response against different types of tumors. Cancer Gene 2003, 10, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Pankhong, P.; Shin, T.H.; Obeng-Adjei, N.; Morrow, M.P.; Walters, J.N.; Khan, A.S.; Sardesai, N.Y.; Weiner, D.B. Highly optimized DNA vaccine targeting human telomerase reverse transcriptase stimulates potent antitumor immunity. Cancer Immunol. Res. 2013, 1, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Thalmensi, J.; Pliquet, E.; Liard, C.; Escande, M.; Bestetti, T.; Julithe, M.; Kostrzak, A.; Pailhes-Jimenez, A.S.; Bourges, E.; Loustau, M.; et al. Anticancer DNA vaccine based on human telomerase reverse transcriptase generates a strong and specific T cell immune response. Oncoimmunology 2016, 5, e1083670. [Google Scholar] [CrossRef]
- Mu, X.; Sang, Y.; Fang, C.; Shao, B.; Yang, L.; Yao, K.; Zhao, X.; Gou, J.; Wei, Y.; Yi, T.; et al. Immunotherapy of tumors with human telomerase reverse transcriptase immortalized human umbilical vein endothelial cells. Int. J. Oncol. 2015, 47, 1901–1911. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Wu, Y.; Ding, Z.Y.; Luo, X.M.; Liu, J.; Zhong, W.N.; Deng, G.H.; Xia, X.Y.; Deng, Y.T.; et al. Mannan-modified adenovirus targeting TERT and VEGFR-2: A universal tumour vaccine. Sci. Rep. 2015, 5, 11275. [Google Scholar] [CrossRef] [Green Version]
- Morgan, R.A.; Dudley, M.E.; Wunderlich, J.R.; Hughes, M.S.; Yang, J.C.; Sherry, R.M.; Royal, R.E.; Topalian, S.L.; Kammula, U.S.; Restifo, N.P.; et al. Cancer regression in patients after transfer of genetically engineered lymphocytes. Science 2006, 314, 126–129. [Google Scholar] [CrossRef]
- Johnson, L.A.; Morgan, R.A.; Dudley, M.E.; Cassard, L.; Yang, J.C.; Hughes, M.S.; Kammula, U.S.; Royal, R.E.; Sherry, R.M.; Wunderlich, J.R.; et al. Gene therapy with human and mouse T-cell receptors mediates cancer regression and targets normal tissues expressing cognate antigen. Blood 2009, 114, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, S.; Ikeda, H.; Miyahara, Y.; Imai, N.; Ishihara, M.; Saito, K.; Sugino, S.; Ueda, S.; Ishikawa, T.; Kokura, S.; et al. Adoptive Transfer of MAGE-A4 T-cell Receptor Gene-Transduced Lymphocytes in Patients with Recurrent Esophageal Cancer. Clin. Cancer Res. 2015, 21, 2268–2277. [Google Scholar] [CrossRef] [Green Version]
- Tawara, I.; Kageyama, S.; Miyahara, Y.; Fujiwara, H.; Nishida, T.; Akatsuka, Y.; Ikeda, H.; Tanimoto, K.; Terakura, S.; Murata, M.; et al. Safety and persistence of WT1-specific T-cell receptor gene-transduced lymphocytes in patients with AML and MDS. Blood 2017, 130, 1985–1994. [Google Scholar] [CrossRef]
- Jackson, H.J.; Rafiq, S.; Brentjens, R.J. Driving CAR T-cells forward. Nat. Rev. Clin. Oncol. 2016, 13, 370–383. [Google Scholar] [CrossRef] [Green Version]
- Neelapu, S.S.; Locke, F.L.; Bartlett, N.L.; Lekakis, L.J.; Miklos, D.B.; Jacobson, C.A.; Braunschweig, I.; Oluwole, O.O.; Siddiqi, T.; Lin, Y.; et al. Axicabtagene Ciloleucel CAR T-Cell Therapy in Refractory Large B-Cell Lymphoma. N. Engl. J. Med. 2017, 377, 2531–2544. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Fujiwara, H.; Asai, H.; Ochi, F.; Ochi, T.; Azuma, T.; Ishida, T.; Okamoto, S.; Mineno, J.; Kuzushima, K.; et al. Development of a novel redirected T-cell-based adoptive immunotherapy targeting human telomerase reverse transcriptase for adult T-cell leukemia. Blood 2013, 121, 4894–4901. [Google Scholar] [CrossRef] [Green Version]
- Kyte, J.A.; Gaudernack, G.; Faane, A.; Lislerud, K.; Inderberg, E.M.; Brunsvig, P.; Aamdal, S.; Kvalheim, G.; Walchli, S.; Pule, M. T-helper cell receptors from long-term survivors after telomerase cancer vaccination for use in adoptive cell therapy. Oncoimmunology 2016, 5, e1249090. [Google Scholar] [CrossRef] [Green Version]
- Ohta, R.; Demachi-Okamura, A.; Akatsuka, Y.; Fujiwara, H.; Kuzushima, K. Improving TCR affinity on 293T cells. J. Immunol. Methods 2019, 466, 1–8. [Google Scholar] [CrossRef]
- Hu, J.; Hwang, S.S.; Liesa, M.; Gan, B.; Sahin, E.; Jaskelioff, M.; Ding, Z.; Ying, H.; Boutin, A.T.; Zhang, H.; et al. Antitelomerase therapy provokes ALT and mitochondrial adaptive mechanisms in cancer. Cell 2012, 148, 651–663. [Google Scholar] [CrossRef]
- Sharma, P.; Allison, J.P. The future of immune checkpoint therapy. Science 2015, 348, 56–61. [Google Scholar] [CrossRef]
- Schumacher, T.N.; Scheper, W.; Kvistborg, P. Cancer Neoantigens. Annu. Rev. Immunol. 2018. [Google Scholar] [CrossRef]
- Terbuch, A.; Lopez, J. Next Generation Cancer Vaccines-Make It Personal! Vaccines 2018, 6. [Google Scholar] [CrossRef]
- Brahmer, J.; Reckamp, K.L.; Baas, P.; Crino, L.; Eberhardt, W.E.; Poddubskaya, E.; Antonia, S.; Pluzanski, A.; Vokes, E.E.; Holgado, E.; et al. Nivolumab versus Docetaxel in Advanced Squamous-Cell Non-Small-Cell Lung Cancer. New Engl. J. Med. 2015, 373, 123–135. [Google Scholar] [CrossRef]
- Marquez-Rodas, I.; Cerezuela, P.; Soria, A.; Berrocal, A.; Riso, A.; Gonzalez-Cao, M.; Martin-Algarra, S. Immune checkpoint inhibitors: Therapeutic advances in melanoma. Ann. Transl. Med. 2015, 3, 267. [Google Scholar] [CrossRef]
- Rittmeyer, A.; Barlesi, F.; Waterkamp, D.; Park, K.; Ciardiello, F.; von Pawel, J.; Gadgeel, S.M.; Hida, T.; Kowalski, D.M.; Dols, M.C.; et al. Atezolizumab versus docetaxel in patients with previously treated non-small-cell lung cancer (OAK): A phase 3, open-label, multicentre randomised controlled trial. Lancet 2017, 389, 255–265. [Google Scholar] [CrossRef]
- Larkin, J.; Hodi, F.S.; Wolchok, J.D. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. New Engl. J. Med. 2015, 373, 1270–1271. [Google Scholar] [CrossRef]
- Hellmann, M.D.; Ciuleanu, T.E.; Pluzanski, A.; Lee, J.S.; Otterson, G.A.; Audigier-Valette, C.; Minenza, E.; Linardou, H.; Burgers, S.; Salman, P.; et al. Nivolumab plus Ipilimumab in Lung Cancer with a High Tumor Mutational Burden. New Engl. J. Med. 2018, 378, 2093–2104. [Google Scholar] [CrossRef]
- Galon, J.; Bruni, D. Approaches to treat immune hot, altered and cold tumours with combination immunotherapies. Nat. Rev. Drug Discov. 2019. [Google Scholar] [CrossRef]


{kind=link}
| Sequence * | Position | HLA Restriction | Immune Response for CD4/CD8 | Year of Report | Refs. |
|---|---|---|---|---|---|
| MPRAPRCRA | 1–9 | HLA-B7 | −/+ | 2006 | [36] |
| RLGPQGWR | 30–37 | HLA-A2 | −/+ | 2007 | [33] |
| RLGPQGWRV | 30–38 | HLA-A2 | −/+ | 2007 | [33] |
| APSFRQVSCL | 68–77 | HLA-B7 | −/+ | 2001 | [41] |
| APSFRQVSCLKELVA | 68–82 | HLA-DR | +/− | 2018 | [29] |
| AYQVCGPPL | 167–175 | HLA-A24 | −/+ | 2006 | [35] |
| RPAEEATSL | 277–285 | HLA-B7 | −/+ | 2006 | [36] |
| VYAETKHFL | 324–332 | HLA-A24 | −/+ | 2006 | [35] |
| YLEPACAKY | 325–333 | HLA-A1 | −/+ | 2005 | [34] |
| RPSFLLSSL | 342–350 | HLA-B7 | −/+ | 2006 | [36] |
| RPSLTGARRL | 351–360 | HLA-B7 | −/+ | 2006 | [36] |
| YWQMRPLFLELLGNH | 386–400 | HLA-DP | +/− | 2011 | [39] |
| DPRRLVQLL | 444–452 | HLA-B7 | −/+ | 2006 | [37] |
| VYGFVRACL | 461–469 | HLA-A24 | −/+ | 2006 | [35] |
| FVRACLRRL | 464–472 | HLA-B7 | −/+ | 2006 | [37] |
| ILAKFLHWL | 540–548 | HLA-A2 | −/+ | 2000 | [30] |
| LAKFLHWLMSVYVVE | 541–555 | HLA-DP | +/− | 2011 | [38] |
| LLRSFFYN | 555–563 | HLA-A2 | −/+ | 2007 | [40] |
| RLFFYRKSV | 572–580 | HLA-A2 | −/+ | 2002 | [31] |
| YLFFYRKSV | 572–580 | HLA-A2 | −/+ | 2002 | [32] |
| LFFYRKSVWSKLQSI | 573–584 | HLA-DP | +/− | 2011 | [38] |
| EARPALLTSRLRFIPK | 611–626 | HLA-DR,DQ,DP | +/− | 2011 | [38] |
| RPALLTSRLRFIPKP | 613–627 | HLA-DP | +/− | 2011 | [38] |
| DYVVGARTF | 637–645 | HLA-A24 | −/+ | 2006 | [35] |
| ALFSVLNYERARRPGLLGASVLGLDDIHRA | 660–689 | HLA-A2,DR | +/+ | 2011 | [39] |
| SVLNYERARRPGLLG | 663–677 | HLA-DR | +/− | 2011 | [39] |
| RPGLLGASVLGLDDI | 672–686 | HLA-DR1,7,15 | +/− | 2002 | [43] |
| PGLLGASVLGLDDIH | 673–687 | HLA-A2,DR | +/+ | 2011 | [39] |
| GLLGASVLGL | 674–683 | HLA-A2 | −/+ | 2011 | [39] |
| LLGASVLGL | 675–683 | HLA-A2 | −/+ | 2012 | [44] |
| LTDLQPYMRQFVAHL | 766–780 | HLA-DR1,7,15 | +/− | 2003 | [42] |
| CYGDMENKL | 845–853 | HLA-A24 | −/+ | 2006 | [35] |
| RLVDDFLLV | 865–873 | HLA-A2 | −/+ | 2000 | [30] |
| KLFGVLRLK | 973–981 | HLA-A2,A3 | −/+ | 2001 | [41] |
| DLQVNSLQTV | 988–997 | HLA-A2 | −/+ | 2002 | [32] |
| YLQVNSLQTV | 988–997 | HLA-A2 | −/+ | 2002 | [32] |
| TYVPLLGSL | 1088–1096 | HLA-A24 | −/+ | 2006 | [35] |
| LPGTTLTAL | 1107–1115 | HLA-B7 | −/+ | 2006 | [37] |
| LPSDFKTIL | 1123–1131 | HLA-B7 | −/+ | 2006 | [37] |
| Name | Clinical Trial Phase | Cancer Targeted | Clinical Response | Adverse Events | Year of Report | Ref. |
|---|---|---|---|---|---|---|
| GV1001 | Phase II (combined with cyclophosphamide) | Hepatocellular carcinoma (HCC) | No clear GV1001-specific immune responses 17/40 SD | Well-tolerated | 2010 | [52] |
| Phase I/II (combined with temozilomide) | Melanoma | Immune responses 5/25 PR, 6/25 SD | Well-tolerated | 2011 | [53] | |
| Phase I/II | Lung and colon cancer and melanoma | Immune responses | Well-tolerated | 2012 | [54] | |
| Phase I/II (combined with or without GM-CSF or gemcitabine) | Pancreatic cancer | Immune responses | Mild vaccination-related adverse events | 2014 | [55] | |
| Phase III (GV1001 with or without gemcitabine and capecitabine) | Pancreatic cancer | Adding GV1001 to chemotherapy did not improve the overall survival of patients. | No additional adverse events | 2014 | [56] | |
| Phase I/II (combined with hTERT540 peptides) | Non-small cell lung cancer (NSCLC) | Immune responses 7/26 SD (1/26CR after clinical trial) | Well-tolerated | 2006 | [57] | |
| Phase I | Melanoma | Immune responses | Well-tolerated | 2011 | [58] | |
| UV1 | Phase I/IIa | Prostate cancer | Immune responses 17/22 SD | Injection site pruritus | 2017 | [59] |
| Vx-001 | Phase I/II | NSCLC | Immune responses 8/22 SD | Well-tolerated; Local skin reactions | 2007 | [49] |
| Phase I/II (optimized Vx-001) | Breast cancer, colorectal cancer, head and neck cancer, HCC, melanoma, prostate cancer, kidney cancer, pancreatic cancer, cholangiocarcinoma, and others with advanced solid tumors, other than NSCLC | Immune responses 1/55 CR, 1/55 PR, 18/55 SD | Well-tolerated | 2012 | [50] | |
| Phase I/II (optimized Vx-001-TERT(572Y)) | Chemo-resistant advanced solid tumors | Immune responses Better clinical outcome in responders than nonresponders | Well-tolerated | 2012 | [51] | |
| Phase II | NSCLC | Immune responses 3/46 PR, 13/46 SD | Well-tolerated | 2014 | [60] | |
| Gx-301 | Phase I/II | Prostate and renal cancer | Immune responses | Well-tolerated | 2013 | [61] |
| hTERT461 | Phase I | HCC | Immune responses | No significant adverse events | 2015 | [62] |
| Dendritic cell vaccines | Phase I (Pulsed with hTERT540 peptide) | Breast and prostate cancer | Immune responses 4/7 SD | No significant adverse events | 2004 | [63] |
| Phase I/II (Transfected with hTERT mRNA) | Prostate cancer | Immune responses; a reduction of PSA and molecular clearance of circulating micrometastases | Well-tolerated | 2005 | [64] | |
| Phase I/II (Pulsed with TERT540 peptide) | Prostate, breast, lung, colorectal, renal, head and neck cancer, and melanoma | Immune responses 4/16 PR | Well-tolerated; mild flu-like symptoms and fever | 2009 | [65] | |
| Phase I/II (GRNVAC1) | Acute myeloid leukemia | Immune responses Favorable disease-free survival | Well-tolerated; local transient erythema | 2010 | [66] | |
| Phase I/II (TAPCells vaccine) | Melanoma and prostate cancer | Immune responses | Well-tolerated | 2013 | [67] | |
| Phase I (DC pulsed with hTERT572, CEA and survivin-derived peptides. | Pancreatic cancer | Immune responses 4/8 SD | Well-tolerated Fatigue and self-limiting flu-like symptoms | 2017 | [68] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mizukoshi, E.; Kaneko, S. Telomerase-Targeted Cancer Immunotherapy. Int. J. Mol. Sci. 2019, 20, 1823. https://doi.org/10.3390/ijms20081823
Mizukoshi E, Kaneko S. Telomerase-Targeted Cancer Immunotherapy. International Journal of Molecular Sciences. 2019; 20(8):1823. https://doi.org/10.3390/ijms20081823
Chicago/Turabian StyleMizukoshi, Eishiro, and Shuichi Kaneko. 2019. "Telomerase-Targeted Cancer Immunotherapy" International Journal of Molecular Sciences 20, no. 8: 1823. https://doi.org/10.3390/ijms20081823
APA StyleMizukoshi, E., & Kaneko, S. (2019). Telomerase-Targeted Cancer Immunotherapy. International Journal of Molecular Sciences, 20(8), 1823. https://doi.org/10.3390/ijms20081823