Two Sides of the Coin: Ezrin/Radixin/Moesin and Merlin Control Membrane Structure and Contact Inhibition

 ,
,
Abstract
:1. Introduction
2. Overview
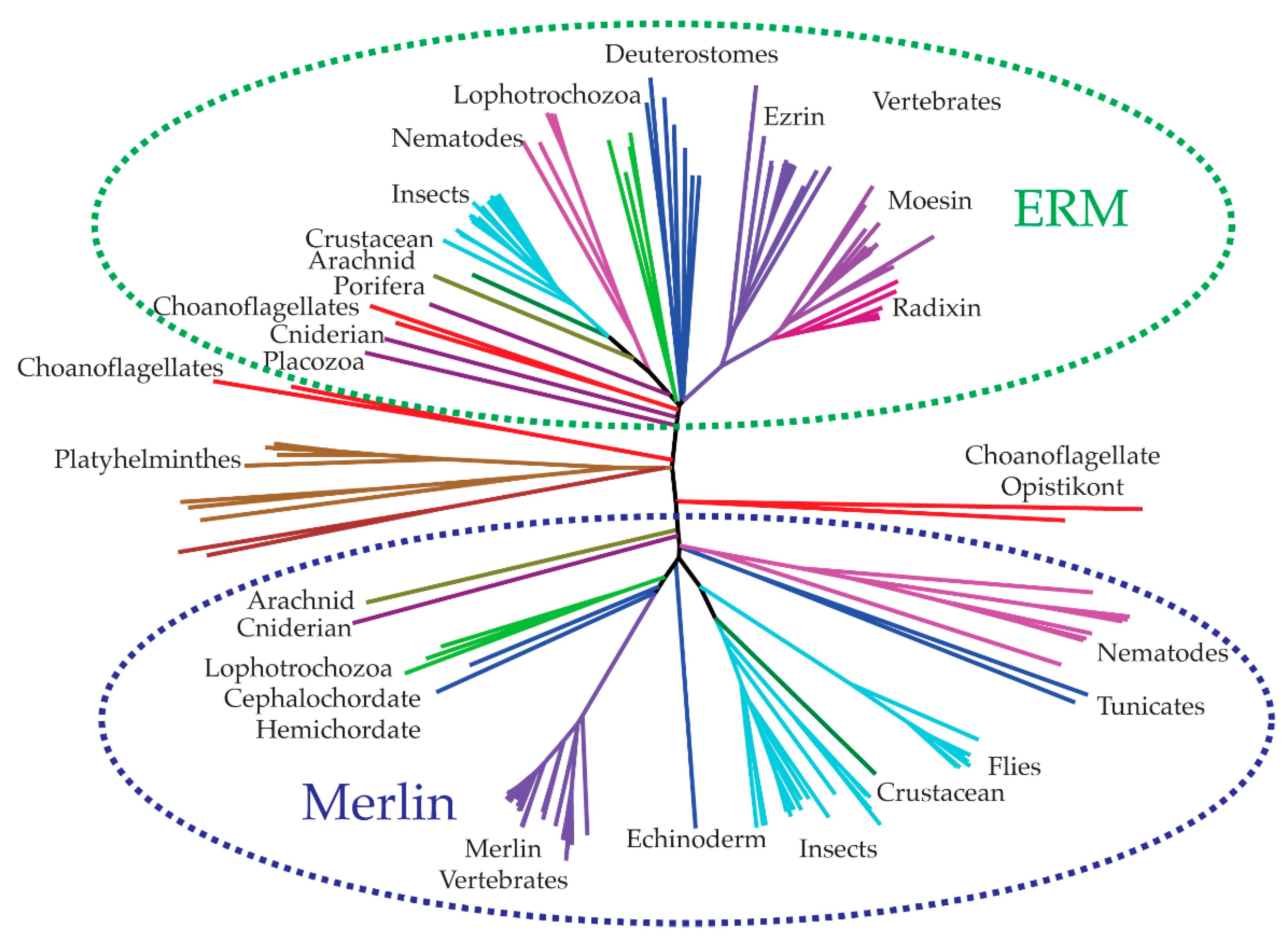
3. Evolution and Biology of Merlin-ERM (Ezrin, Radixin and Moesin) Proteins
3.1. Evolution of the Merlin-ERM Family of Protein
3.2. Isoforms, Biological Expression and Disease
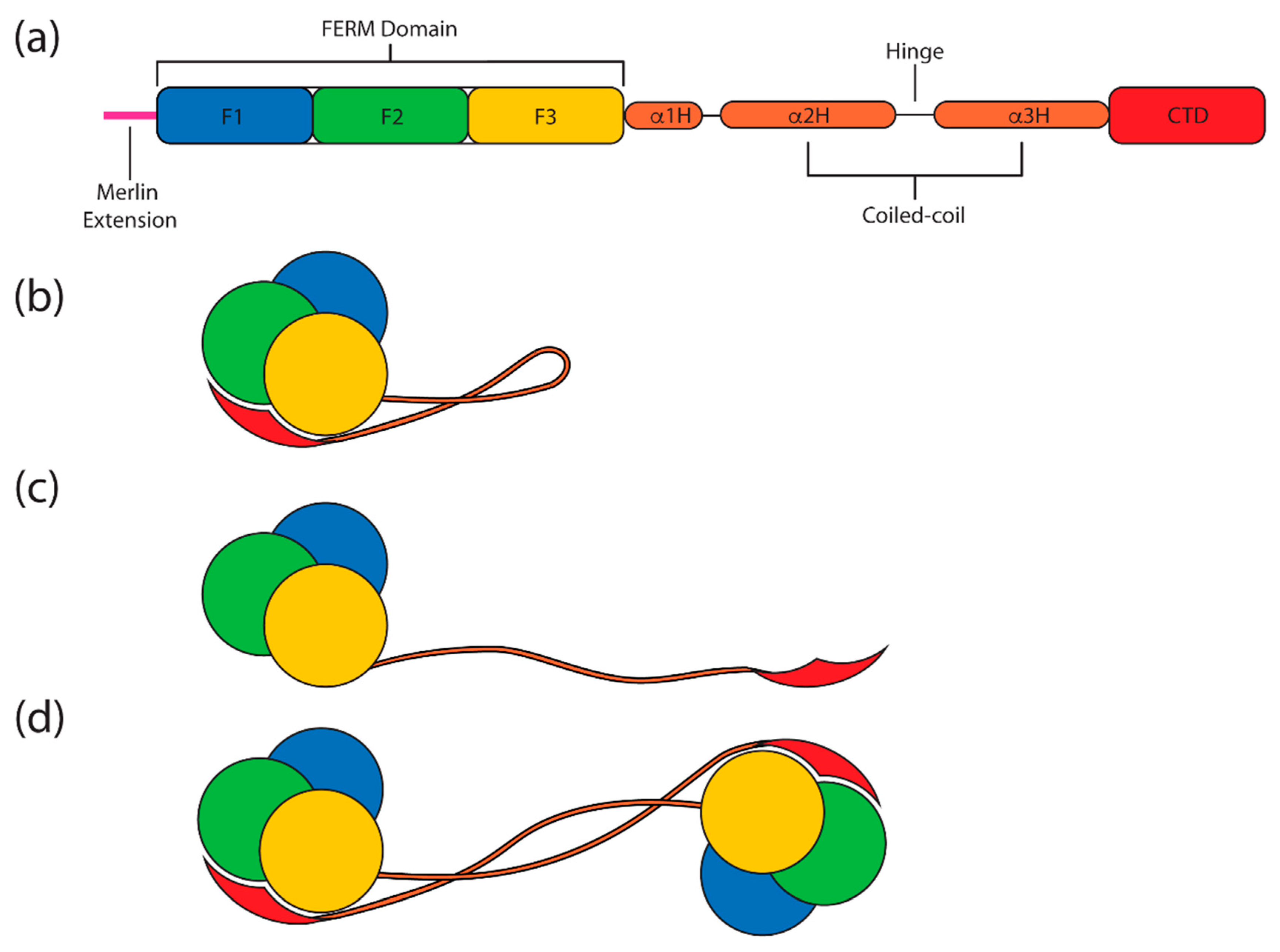
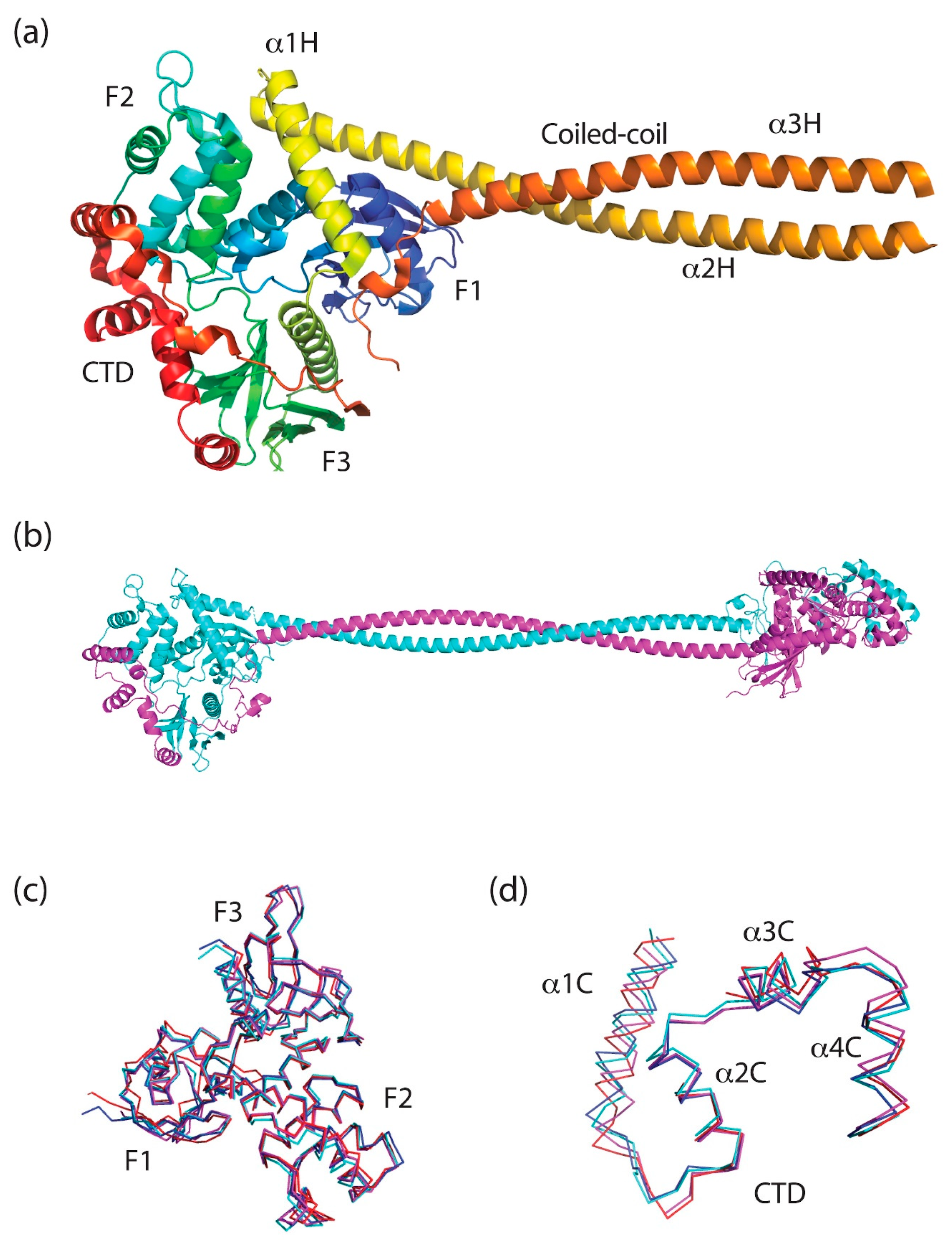
4. Merlin-ERM Structure
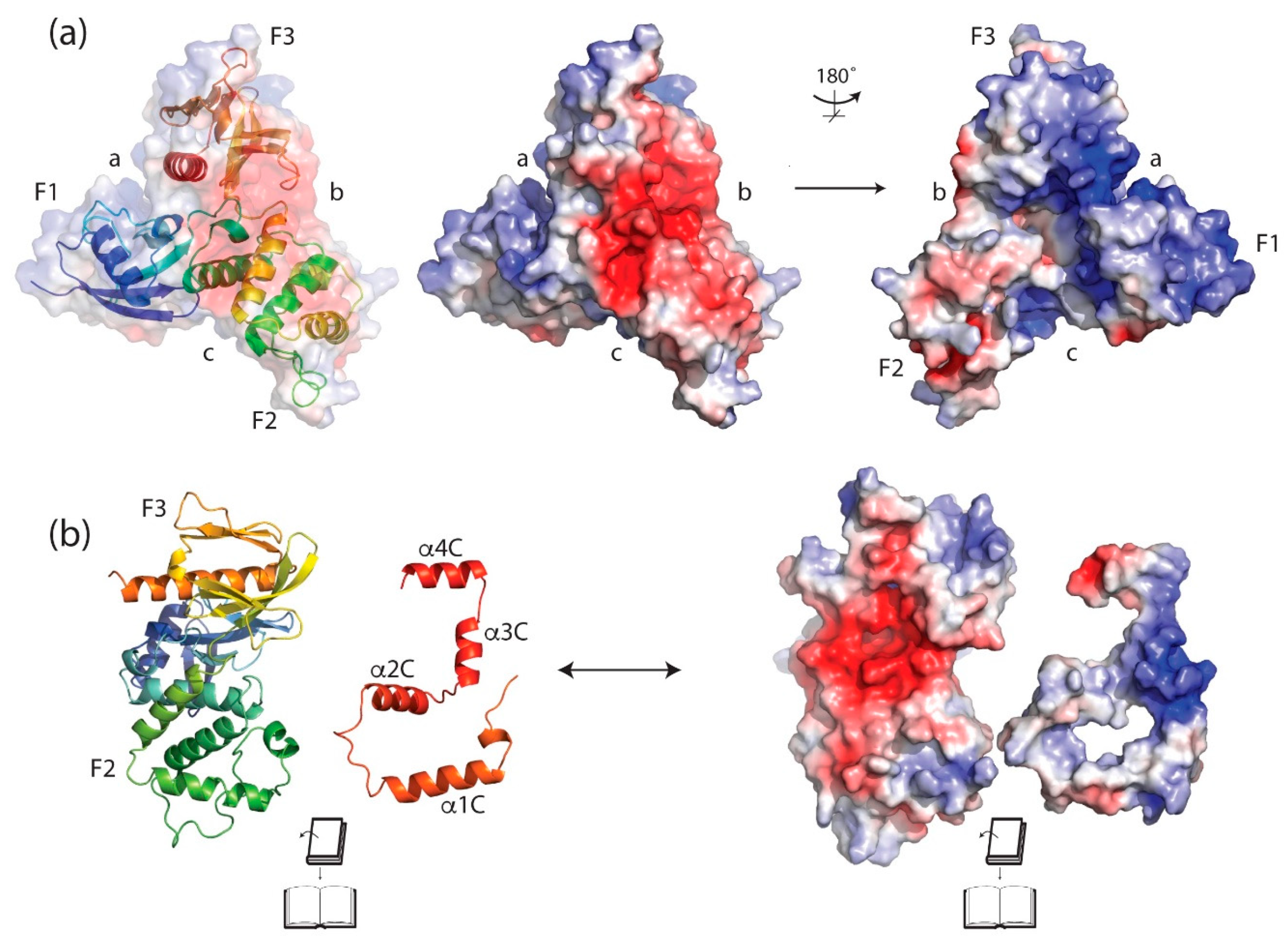
4.1. The FERM (Band 4.1 Protein, Ezrin, Radixin, Moesin) Domain
4.2. The Central Helical Domain
The Hinge Sequence
4.3. The C-Terminal Domain (CTD) or C-Terminal ERM Association Domain (C-ERMAD)
Polyproline Region
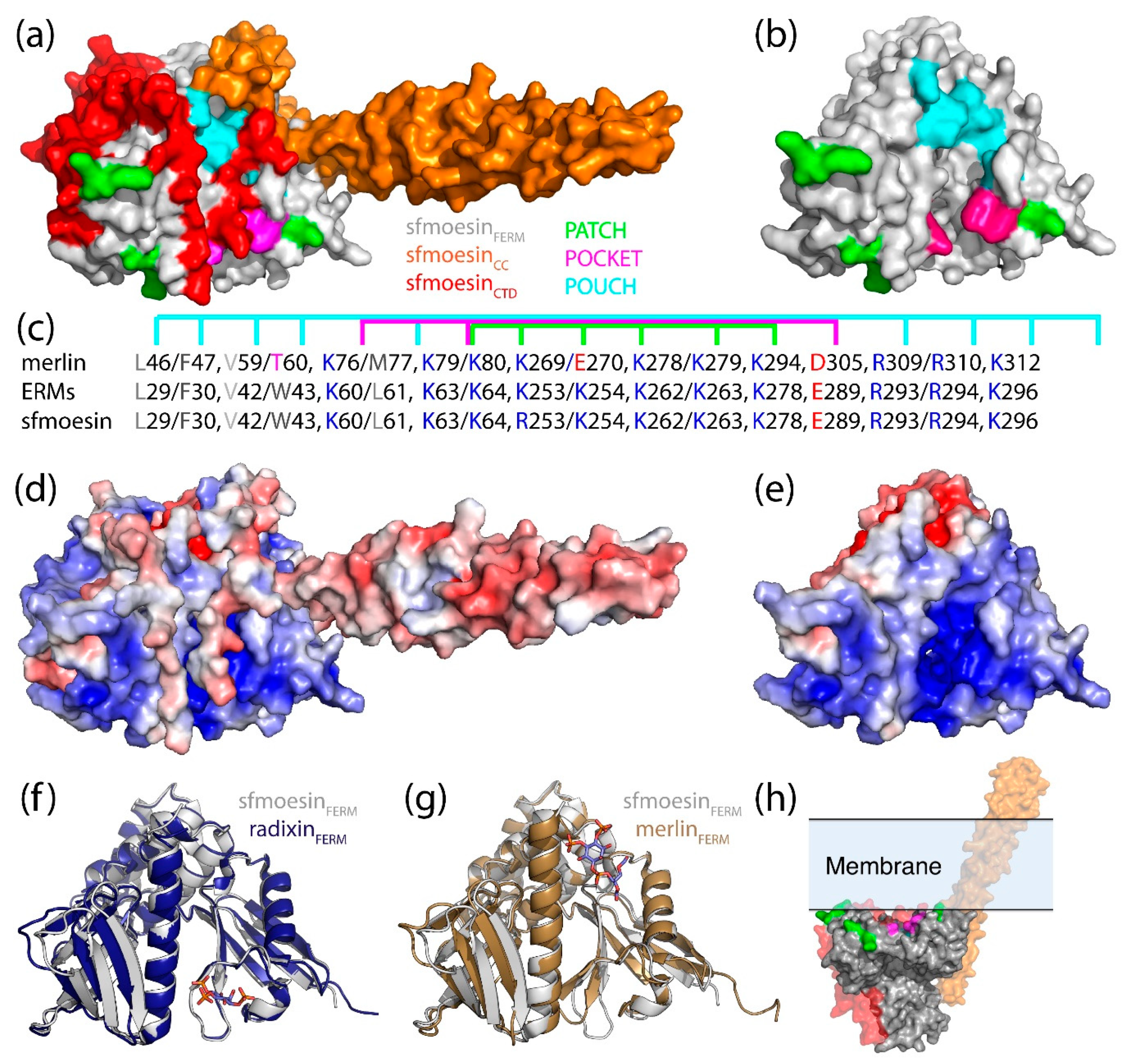
4.4. Interaction between the CTD and FERM Domain and Implications
4.5. Oligomeric States of Merlin-ERM Proteins
4.6. The Open and Closed Forms of ERM Proteins
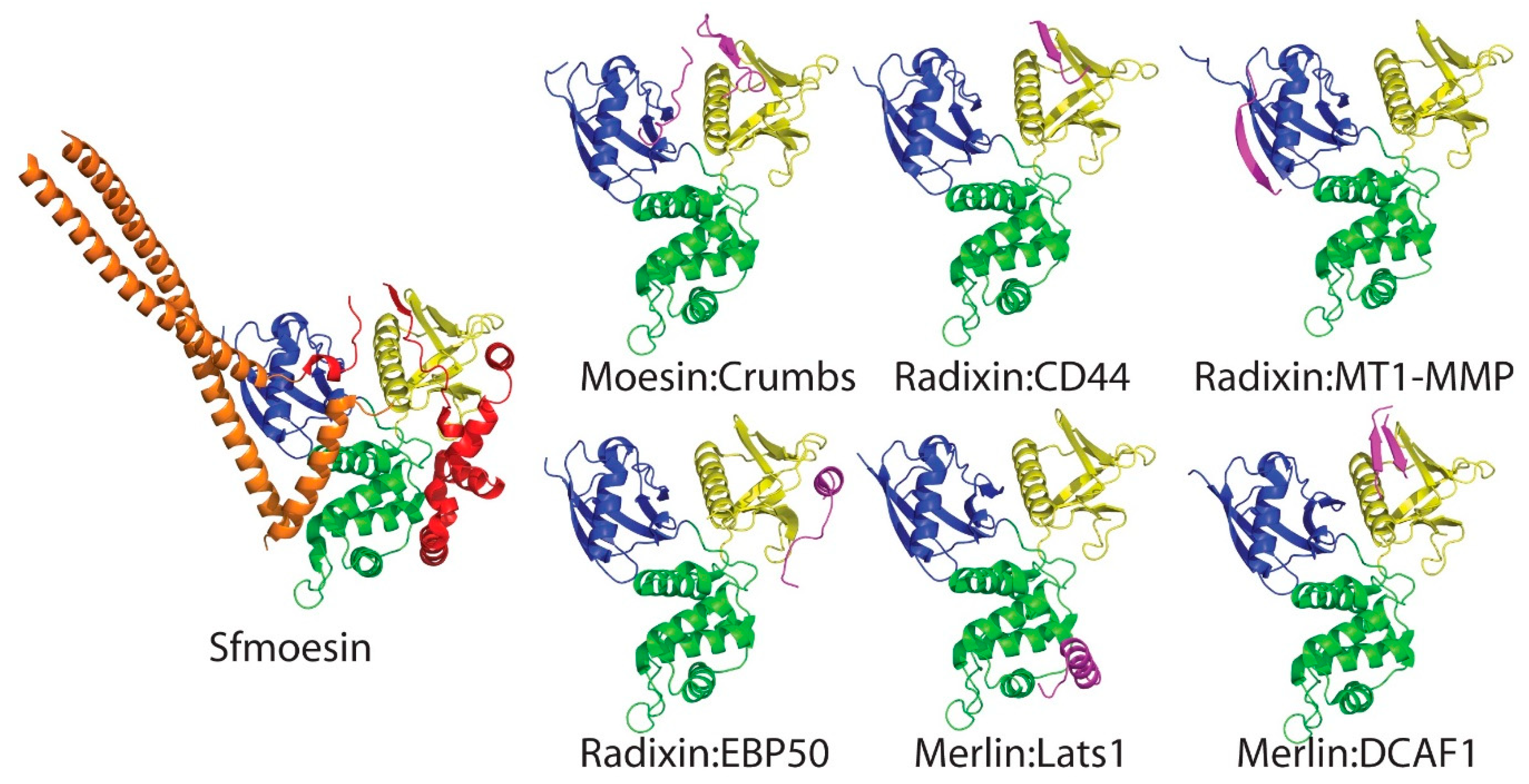
5. Complexes between Merlin-ERM Protein FERM Domains and Their Binding Partners: The FERM as a Signaling Hub and Integrator
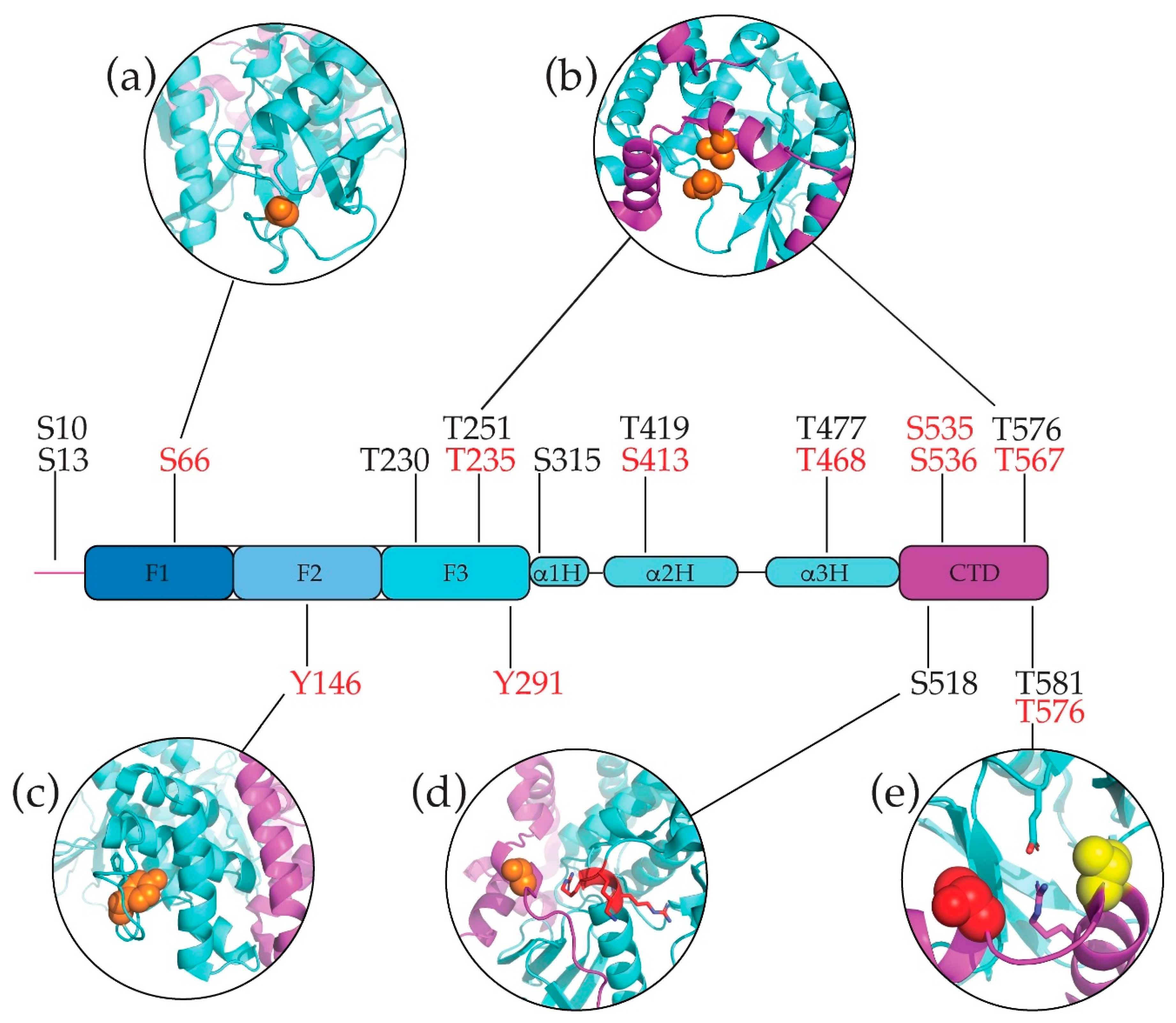
6. Post-Translational Modifications (PTM) of Merlin-ERM Proteins
6.1. Phosphorylation
6.1.1. Common Phosphorylated Merlin-ERM Residues
6.1.2. Merlin Specific Phosphorylation—Serines and Threonines
6.1.3. ERM Phosphorylation
6.2. Cysteine Modifications: S-Nitrosylation of the Conserved F3 Loop
6.3. Ubiquitination and Acetylation of Merlin-ERM Proteins
6.4. Sumoylation of Merlin-ERMs
6.5. Succinylation
6.6. ADP-Ribosylation of ERM
7. Common Structural and Functional Characteristics of Merlin-ERM Proteins
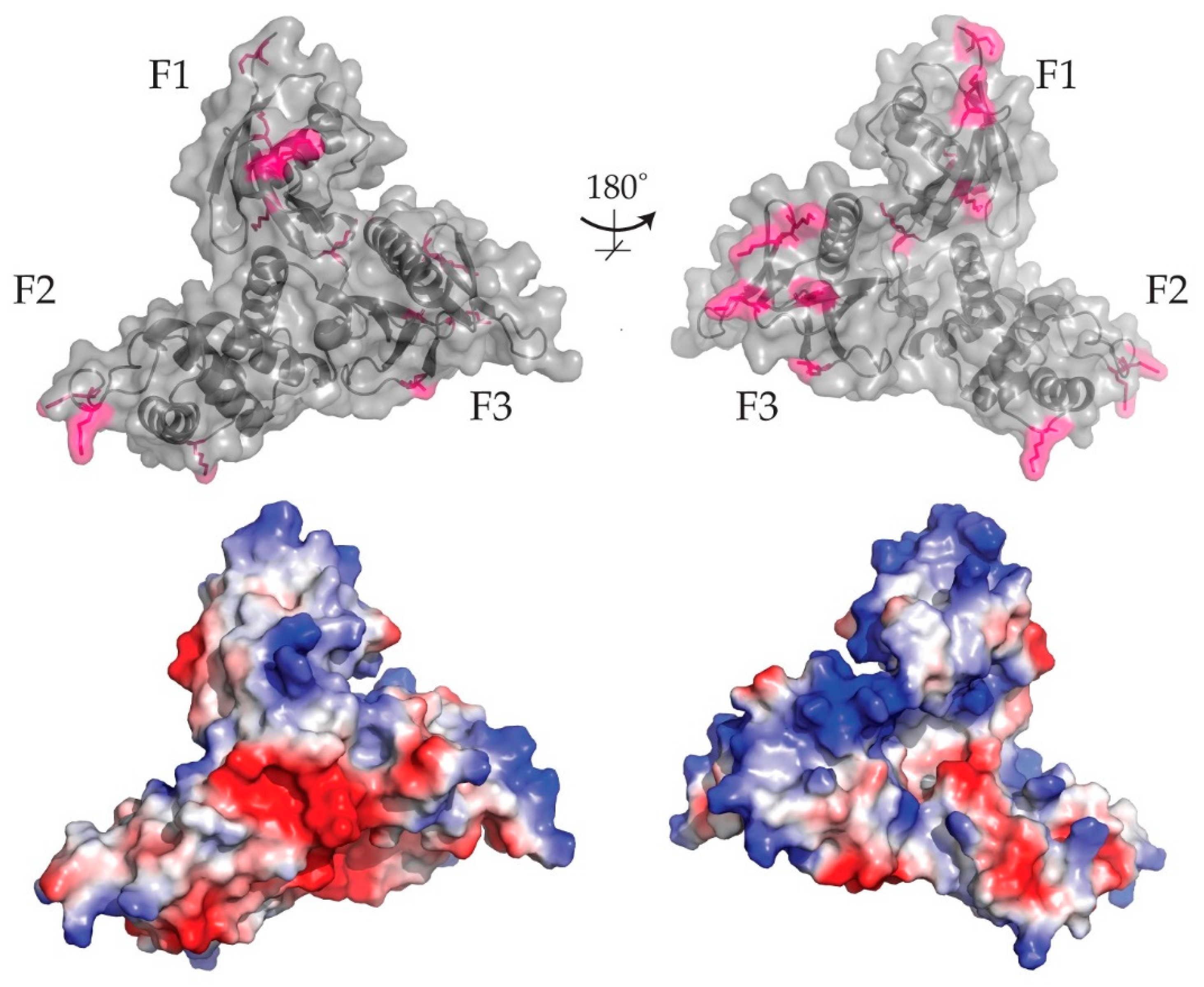
7.1. Lipid Binding
7.2. Interactions with Microtubules
8. Specific Structural and Functional Characteristic—the Differences between Merlin and the ERMs
8.1. Merlin and the Nucleus
8.2. C-Terminal ERM Actin-Binding Domain
8.3. Merlin–Actin Binding
9. Discussion
10. Speculation
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ERM | Ezrin, radixin, moesin |
| FERM | Protein 4.1, ezrin, radixin, moesin domain |
| CTD | C-terminal domain |
| PI(4,5)P2 | Phosphatidylinositol 4,5-bisphosphate |
| PI(4,5)P2diC8 | Phosphatidylinositol 4,5-bisphosphate with two 8-carbon fatty acid moieties |
| PPII | Poly-proline type II helix |
| ADP | Adenosine diphosphate |
References
- Turunen, O.; Sainio, M.; Jääskeläinen, J.; Carpén, O.; Vaheri, A. Structure-function relationships in the ezrin family and the effect of tumor-associated point mutations in neurofibromatosis 2 protein. Biochim. Biophys. Acta-Protein Struct. Mol. Enzymol. 1998, 1387, 1–16. [Google Scholar] [CrossRef]
- Bretscher, A.; Edwards, K.; Fehon, R.G. ERM proteins and merlin: Integrators at the cell cortex. Nat. Rev. Mol. Cell Biol. 2002, 3, 586–599. [Google Scholar] [CrossRef]
- Fehon, R.G.; McClatchey, A.I.; Bretscher, A. Organizing the cell cortex: The role of ERM proteins. Nat. Rev. Mol. Cell Biol. 2010, 11, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Fiévet, B.; Louvard, D.; Arpin, M. ERM proteins in epithelial cell organization and functions. Biochim. Biophys. Acta-Mol. Cell Res. 2007, 1173, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Louvet-Vallée, S. ERM proteins: From cellular architecture to cell signaling. Biol. Cell 2000, 92, 305–316. [Google Scholar] [CrossRef]
- Neisch, A.L.; Fehon, R.G. Ezrin, Radixin and Moesin: Key regulators of membrane-cortex interactions and signaling. Curr. Opin. Cell Biol. 2011, 23, 377–382. [Google Scholar] [CrossRef] [Green Version]
- McClatchey, A.I.; Fehon, R.G. Merlin and the ERM proteins-regulators of receptor distribution and signaling at the cell cortex. Trends Cell Biol. 2009, 19, 198–206. [Google Scholar] [CrossRef]
- McClatchey, A.I. ERM proteins at a glance. J. Cell Sci. 2014, 127, 3199–3204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautreau, A.; Louvard, D.; Arpin, M. ERM proteins and NF2 tumor suppressor: The Yin and Yang of cortical actin organization and cell growth signaling. Curr. Opin. Cell Biol. 2002, 14, 104–109. [Google Scholar] [CrossRef]
- Bretscher, A.; Chambers, D.; Nguyen, R.; Reczek, D. ERM-Merlin and EBP50 protein families in plasma membrane organization and function. Annu. Rev. Cell Dev. Biol. 2000, 16, 113–143. [Google Scholar] [CrossRef]
- Li, W.; Cooper, J.; Karajannis, M.A.; Giancotti, F.G. Merlin: A tumour suppressor with functions at the cell cortex and in the nucleus. EMBO Rep. 2012, 13, 204–215. [Google Scholar] [CrossRef]
- Yu, F.X.; Guan, K.L. The Hippo pathway: Regulators and regulations. Genes Dev. 2013, 27, 355–371. [Google Scholar] [CrossRef] [PubMed]
- Morrow, K.A.; Das, S.; Meng, E.; Menezes, M.E.; Sarah, K.; Metge, B.J.; Buchsbaum, D.J.; Samant, R.S.; Lalita, A. Loss of tumor suppressor Merlin results in aberrant activation of Wnt/β-catenin signaling in cancer. Oncotarget 2016, 7, 17991–18005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3420. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004. [Google Scholar] [CrossRef]
- Golovnina, K.; Blinov, A.; Akhmametyeva, E.M.; Omelyanchuk, L.V.; Chang, L.S. Evolution and origin of merlin, the product of the Neurofibromatosis type 2 (NF2) tumor-suppressor gene. BMC Evol. Biol. 2005, 5. [Google Scholar] [CrossRef]
- Ohno, S. Evolution by gene duplication; Springer: Berlin/Heidelberg, Germany, 1970. [Google Scholar]
- Lien, S.; Koop, B.F.; Sandve, S.R.; Miller, J.R.; Kent, M.P.; Nome, T.; Hvidsten, T.R.; Leong, J.S.; Minkley, D.R.; Zimin, A.; et al. The Atlantic salmon genome provides insights into rediploidization. Nature 2016, 533, 200–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, J.M.; Zvelebil, M.J.; Cummings, M.P. FigTree. In Dictionary of Bioinformatics and Computational Biology; Wiley-Liss: Chichester, UK, 2014. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2-A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Lagresle-Peyrou, C.; Luce, S.; Ouchani, F.; Soheili, T.S.; Sadek, H.; Chouteau, M.; Durand, A.; Pic, I.; Majewski, J.; Brouzes, C.; et al. X-linked primary immunodeficiency associated with hemizygous mutations in the moesin (MSN) gene. J. Allergy Clin. Immunol. 2016, 138, 1681–1689. [Google Scholar] [CrossRef] [PubMed]
- Dunster, L.M.; Schneider-schaulies, R.; Dunster, L.E.E.M.; Schwartz-albiez, R. Physical Association of Moesin and CD46 as a Receptor Complex for Measles Virus. J. Virol. 1995, 69, 2248–2256. [Google Scholar]
- Hecker, C.; Weise, C.; Schneider-Schaulies, J.; Holmes, H.C.; Ter Meulen, V. Specific binding of HIV-1 envelope protein gp120 to the structural membrane proteins ezrin and moesin. Virus Res. 1997, 49, 215–223. [Google Scholar] [CrossRef]
- Bukong, T.N.; Kodys, K.; Szabo, G. Human Ezrin-Moesin-Radixin Proteins Modulate Hepatitis C Virus Infection. Hepatology 2013, 5, 1569–1579. [Google Scholar] [CrossRef] [PubMed]
- Pines, M.; Levi, O.; Genin, O.; Lavy, A.; Angelini, C.; Allamand, V.; Halevy, O. Elevated Expression of Moesin in Muscular Dystrophies. Am. J. Pathol. 2017, 187, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Charafe-Jauffret, E.; Monville, F.; Bertucci, F.; Esterni, B.; Ginestier, C.; Finetti, P.; Cervera, N.; Geneix, J.; Hassanein, M.; Rabayrol, L.; et al. Moesin expression is a marker of basal breast carcinomas. Int. J. Cancer 2007, 121, 1779–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Cheng, Y.; Li, W.; Liu, Q.; Liu, J.; Huang, J.; Fu, X. Vascular endothelial growth factor C promotes cervical cancer metastasis via up-regulation and activation of RhoA/ROCK-2/moesin cascade. BMC Cancer 2010, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Keshamouni, V.G.; Michailidis, G.; Grasso, C.S.; Anthwal, S.; Strahler, J.R.; Walker, A.; Arenberg, D.A.; Reddy, R.C.; Akulapalli, S.; Thannickal, V.J.; et al. Differential protein expression profiling by iTRAQ-2DLC-MS/MS of lung cancer cells undergoing epithelial-mesenchymal transition reveals a migratory/invasive phenotype. J. Proteome Res. 2006, 5, 1143–1154. [Google Scholar] [CrossRef]
- Yu, Y.; Khan, J.; Khanna, C.; Helman, L.; Meltzer, P.S.; Merlino, G. Expression profiling identifies the cytoskeletal organizer ezrin and the developmental homeoprotein Six-1 as key metastatic regulators. Nat. Med. 2004, 10, 175. [Google Scholar] [CrossRef]
- Khanna, C.; Wan, X.; Bose, S.; Cassaday, R.; Olomu, O.; Mendoza, A.; Yeung, C.; Gorlick, R.; Hewitt, S.M.; Helman, L.J. The membrane-cytoskeleton linker ezrin is necessary for osteosarcoma metastasis. Nat. Med. 2004, 10, 182–186. [Google Scholar] [CrossRef]
- Curto, M.; McClatchey, A.I. Ezrin… a metastatic detERMinant? Cancer Cell 2004, 5, 113–114. [Google Scholar] [CrossRef] [Green Version]
- Elliott, B.E.; Meens, J.A.; SenGupta, S.K.; Louvard, D.; Arpin, M. The membrane cytoskeletal crosslinker ezrin is required for metastasis of breast carcinoma cells. Breast Cancer Res. 2005, 7, R365. [Google Scholar] [CrossRef]
- Meng, Y.; Lu, Z.; Yu, S.; Zhang, Q.; Ma, Y.; Chen, J. Ezrin promotes invasion and metastasis of pancreatic cancer cells. J. Transl. Med. 2010, 8, 61. [Google Scholar] [CrossRef]
- Bruce, B.; Khanna, G.; Ren, L.; Landberg, G.; Jirström, K.; Powell, C.; Borczuk, A.; Keller, E.T.; Wojno, K.J.; Meltzer, P.; et al. Expression of the cytoskeleton linker protein ezrin in human cancers. Clin. Exp. Metastasis 2007, 24, 69–78. [Google Scholar] [CrossRef]
- Leiphrakpam, P.D.; Rajput, A.; Mathiesen, M.; Agarwal, E.; Lazenby, A.J.; Are, C.; Brattain, M.G.; Chowdhury, S. Ezrin expression and cell survival regulation in colorectal cancer. Cell. Signal. 2014, 26, 868–879. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Mao, W.; Zheng, S. MicroRNA-183 regulates Ezrin expression in lung cancer cells. FEBS Lett. 2008, 582, 3663–3668. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.; Li, Y.; Liu, S.; Jin, H.; Shang, Y.; Quan, C.; Li, Y.; Lin, Z. High expression of ezrin predicts poor prognosis in uterine cervical cancer. BMC Cancer 2013, 13, 520. [Google Scholar] [CrossRef]
- Roy, N.H.; Lambele, M.; Chan, J.; Symeonides, M.; Thali, M. Ezrin Is a Component of the HIV-1 Virological Presynapse and Contributes to the Inhibition of Cell-Cell Fusion. J. Virol. 2014, 88, 7645–7658. [Google Scholar] [CrossRef] [Green Version]
- Millet, J.K.; Kien, F.; Cheung, C.Y.; Siu, Y.L.; Chan, W.L.; Li, H.; Leung, H.L.; Jaume, M.; Bruzzone, R.; Malik Peiris, J.S.; et al. Ezrin Interacts with the SARS Coronavirus Spike Protein and Restrains Infection at the Entry Stage. PLoS ONE 2012, 7, e49566. [Google Scholar] [CrossRef] [Green Version]
- Yeh, C.N.; Pang, S.T.; Chen, T.W.; Wu, R.C.; Weng, W.H.; Chen, M.F. Expression of ezrin is associated with invasion and dedifferentiation of hepatitis B related hepatocellular carcinoma. BMC Cancer 2009, 9, 233. [Google Scholar] [CrossRef]
- Xie, J.J.; Zhang, F.R.; Tao, L.H.; Lü, Z.; Xu, X.E.; Jian-Shen; Xu, L.Y.; Li, E.M. Expression of Ezrin in Human Embryonic, Fetal, and Normal Adult Tissues. J. Histochem. Cytochem. 2011, 59, 1001–1008. [Google Scholar] [CrossRef] [Green Version]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Bianchi, A.B.; Hara, T.; Ramesh, V.; Gao, J.; Klein-Szanto, A.J.P.; Morin, F.; Menon, A.G.; Trofatter, J.A.; Gusella, J.F.; Seizinger, B.R.; et al. Mutations in transcript isoforms of the neurofibromatosis 2 gene in multiple human tumour types. Nat. Genet. 1994, 6, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Sherman, L.; Xu, H.; Geist, R.; Saporito-Irwin, S.; Howells, N.; Ponta, H.; Herrlich, P.; Gutmann, D. Interdomain binding mediates tumor growth suppression by the NF2 gene product. Oncogene 1997, 15, 2505–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, D.G.R. Neurofibromatosis type 2 (NF2): A clinical and molecular review. Orphanet J. Rare Dis. 2009, 4, 1–11. [Google Scholar] [CrossRef]
- Brodhun, M.; Stahn, V.; Harder, A. Pathogenesis and molecular pathology of vestibular schwannoma. HNO 2017, 65, 362–372. [Google Scholar] [CrossRef]
- Sekido, Y. Molecular pathogenesis of malignant mesothelioma. Carcinogenesis 2013, 34, 1413–1419. [Google Scholar] [CrossRef] [Green Version]
- Kitajiri, S.I.; Fukumoto, K.; Hata, M.; Sasaki, H.; Katsuno, T.; Nakagawa, T.; Ito, J.; Tsukita, S.; Tsukita, S. Radixin deficiency causes deafness associated with progressive degeneration of cochlear stereocilia. J. Cell Biol. 2004, 166, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Pataky, F.; Pironkova, R.; Hudspeth, A.J. Radixin is a constituent of stereocilia in hair cells. Proc. Natl. Acad. Sci. USA 2004, 101, 2601–2606. [Google Scholar] [CrossRef] [Green Version]
- Sauvanet, C.; Wayt, J.; Pelaseyed, T.; Bretscher, A. Structure, Regulation, and Functional Diversity of Microvilli on the Apical Domain of Epithelial Cells. Annu. Rev. Cell Dev. Biol. 2015, 31, 593–621. [Google Scholar] [CrossRef]
- Pelaseyed, T.; Bretscher, A. Regulation of actin-based apical structures on epithelial cells. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef]
- Khan, S.Y.; Ahmed, Z.M.; Shabbir, M.I.; Kitajiri, S.; Kalsoom, S.; Tasneem, S.; Shayiq, S.; Ramesh, A.; Srisailpathy, S.; Khan, S.N.; et al. Mutations of the RDX gene cause nonsyndromic hearing loss at the DFNB24 locus. Hum. Mutat. 2007, 28, 417–423. [Google Scholar] [CrossRef]
- Jiang, Q.H.; Wang, A.X.; Chen, Y. Radixin enhances colon cancer cell invasion by increasing MMP-7 production via rac1-ERK pathway. Sci. World J. 2014. [Google Scholar] [CrossRef]
- Li, Q.; Nance, M.R.; Kulikauskas, R.; Nyberg, K.; Fehon, R.; Karplus, P.A.; Bretscher, A.; Tesmer, J.J.G. Self-masking in an Intact ERM-merlin Protein: An Active Role for the Central α-Helical Domain. J. Mol. Biol. 2007, 365, 1446–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phang, J.M.; Harrop, S.J.; Duff, A.P.; Sokolova, A.V.; Crossett, B.; Walsh, J.C.; Beckham, S.A.; Nguyen, C.D.; Davies, R.B.; Glöckner, C.; et al. Structural characterization suggests models for monomeric and dimeric forms of full-length ezrin. Biochem. J. 2016, 473, 2763–2782. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.J.; Nassar, N.; Bretscher, A.; Cerione, R.A.; Karplus, P.A. Structure of the active N-terminal domain of ezrin: Conformational and mobility changes identify keystone interactions. J. Biol. Chem. 2003, 278, 4949–4956. [Google Scholar] [CrossRef] [PubMed]
- Hamada, K.; Shimizu, T.; Matsui, T.; Tsukita, S.; Hakoshima, T. Structural basis of the membrane-targeting and unmasking mechanisms of the radixin FERM domain. EMBO J. 2000, 19, 4449–4462. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.D.; Keep, N.H. The 2.7 A crystal structure of the activated FERM domain of moesin: An analysis of structural changes on activation. Biochemistry 2001, 40, 7061–7068. [Google Scholar] [CrossRef]
- Shimizu, T.; Seto, A.; Maita, N.; Hamada, K.; Tsukita, S.; Tsukita, S.; Hakoshima, T. Structural Basis for Neurofibromatosis Type 2. J. Biol. Chem. 2002, 277, 10332–10336. [Google Scholar] [CrossRef]
- Pearson, M.A.; Reczek, D.; Bretscher, A.; Karplus, P.A. Structure of the ERM protein moesin reveals the FERM domain fold masked by an extended actin binding tail domain. Cell 2000, 101, 259–270. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, H.; Li, F.; Chan, S.W.; Lin, Z.; Wei, Z.; Yang, Z.; Guo, F.; Lim, C.J.; Xing, W.; et al. Angiomotin binding-induced activation of Merlin/NF2 in the Hippo pathway. Cell Res. 2015, 25, 801–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrödinger LLC The PyMOL Molecular Graphics System. Schrödinger LLC 2016, Version 1. Available online: http://www.pymol.org (accessed on 23 April 2019).
- Chishti, A.H.; Kim, A.C.; Marfatia, S.M.; Lutchman, M.; Hanspal, M.; Jindal, H.; Liu, S.C.; Low, P.S.; Rouleau, G.A.; Mohandas, N.; et al. The FERM domain: A unique module involved in the linkage of cytoplasmic proteins to the membrane. Trends Biochem. Sci. 1998, 23, 281–282. [Google Scholar] [CrossRef]
- Elliott, P.R.; Goult, B.T.; Kopp, P.M.; Bate, N.; Grossmann, J.G.; Roberts, G.C.K.; Critchley, D.R.; Barsukov, I.L. The Structure of the Talin Head Reveals a Novel Extended Conformation of the FERM Domain. Structure 2010, 18, 1289–1299. [Google Scholar] [CrossRef] [Green Version]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS biomolecular solvation software suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Pan, L.; Wei, Z.; Zhang, M. Structure of myTH4-FERM domains in myosin VIIa tail bound to cargo. Science 2011, 331, 757–760. [Google Scholar] [CrossRef]
- Yu, I.M.; Planelles-Herrero, V.J.; Sourigues, Y.; Moussaoui, D.; Sirkia, H.; Kikuti, C.; Stroebel, D.; Titus, M.A.; Houdusse, A. Myosin 7 and its adaptors link cadherins to actin. Nat. Commun. 2017, 8, 15864. [Google Scholar] [CrossRef]
- Li, J.; He, Y.; Weck, M.L.; Lu, Q.; Tyska, M.J.; Zhang, M. Structure of Myo7b/USH1C complex suggests a general PDZ domain binding mode by MyTH4-FERM myosins. Proc. Natl. Acad. Sci. USA 2017, 114, E3776–E3785. [Google Scholar] [CrossRef]
- Chinthalapudi, K.; Mandati, V.; Zheng, J.; Sharff, A.J.; Bricogne, G.; Griffin, P.R.; Kissil, J.; Izard, T. Lipid binding promotes the open conformation and tumor-suppressive activity of neurofibromin 2. Nat. Commun. 2018, 9, 1338. [Google Scholar] [CrossRef] [Green Version]
- Sellers, J.R. Myosins: A diverse superfamily. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2000, 1496, 3–22. [Google Scholar] [CrossRef]
- Hamada, K.; Shimizu, T.; Yonemura, S.; Tsukita, S.; Tsukita, S.; Hakoshima, T. Structural basis of adhesion-molecule recognition by ERM proteins revealed by the crystal structure of the radixin-ICAM-2 complex. EMBO J. 2003, 22, 502–514. [Google Scholar] [CrossRef]
- Takai, Y.; Kitano, K.; Terawaki, S.; Maesaki, R.; Hakoshima, T. Structural Basis of the Cytoplasmic Tail of Adhesion Molecule CD43 and Its Binding to ERM Proteins. J. Mol. Biol. 2008, 381, 634–644. [Google Scholar] [CrossRef]
- Takai, Y.; Kitano, K.; Terawaki, S.I.; Maesaki, R.; Hakoshima, T. Structural basis of PSGL-1 binding to ERM proteins. Genes Cells 2007, 12, 1329–1338. [Google Scholar] [CrossRef] [Green Version]
- Lupas, A.; Van Dyke, M.; Stock, J. Predicting coiled coils from protein sequences. Science 1991, 252, 1162–1164. [Google Scholar] [CrossRef]
- Gould, K.L.; Bretscher, A.; Esch, F.S.; Hunter, T. cDNA cloning and sequencing of the protein-tyrosine kinase substrate, ezrin, reveals homology to band 4.1. EMBO J. 1989, 8, 4133–4142. [Google Scholar] [CrossRef]
- Gautreau, A.; Poullet, P.; Louvard, D.; Arpin, M. Ezrin, a plasma membrane-microfilament linker, signals cell survival through the phosphatidylinositol 3-kinase/Akt pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 7300–7305. [Google Scholar] [CrossRef] [Green Version]
- Mandal, A.; Mandal, S.; Park, M.H. Global quantitative proteomics reveal up-regulation of endoplasmic reticulum stress response proteins upon depletion of eIF5A in HeLa cells. Sci. Rep. 2016, 6, 25795. [Google Scholar] [CrossRef]
- Hersch, S.J.; Wang, M.; Zou, S.B.; Moon, K.M.; Foster, L.J.; Ibba, M.; Navarre, W.W. Divergent protein motifs direct elongation factor P-mediated translational regulation in salmonella enterica and Escherichia coli. MBio 2013, 4, e00180-13. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Awad, R.; Marion, S.; Isabel, A.; Anne, C.; Philippe, F.; Pierre, G.; Jean-Baptiste, R.; Jean-Philippe, K. The SH3 regulatory domain of the hematopoietic cell kinase Hck binds ELMO via its polyproline motif. FEBS Open Bio 2015, 5, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Pellicena, P.; Miller, W.T. Processive Phosphorylation of p130Cas by Src Depends on SH3-Polyproline Interactions. J. Biol. Chem. 2001, 276, 28190–28196. [Google Scholar] [CrossRef] [Green Version]
- Lewitzky, M.; Harkiolaki, M.; Domart, M.C.; Jones, E.Y.; Feller, S.M. Mona/Gads SH3C binding to hematopoietic progenitor kinase 1 (HPK1) combines an atypical SH3 binding motif, R/KXXK, with a classical PXXP motif embedded in a polyproline type II (PPII) helix. J. Biol. Chem. 2004, 279, 28724–28732. [Google Scholar] [CrossRef]
- Yu, H.; Chen, J.K.; Feng, S.; Dalgarno, D.C.; Brauer, A.W.; Schrelber, S.L. Structural basis for the binding of proline-rich peptides to SH3 domains. Cell 1994, 76, 933–945. [Google Scholar] [CrossRef]
- Wu, X.; Knudsen, B.; Feller, S.M.; Zheng, J.; Sali, A.; Cowburn, D.; Hanafusa, H.; Kuriyan, J. Structural basis for the specific interaction of lysine-containing proline-rich peptides with the N-terminal SH3 domain of c-Crk. Structure 1995, 3, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Ghose, R.; Shekhtman, A.; Goger, M.J.; Ji, H.; Cowburn, D. A novel, specific interaction involving the Csk SH3 domain and its natural ligand. Nat. Struct. Biol. 2001, 8, 998–1004. [Google Scholar] [CrossRef]
- Schlessinger, J. SH2/SH3 signaling proteins. Curr. Opin. Genet. Dev. 1994, 4, 25–30. [Google Scholar] [CrossRef]
- Cubellis, M.V.; Caillez, F.; Blundell, T.L.; Lovell, S.C. Properties of polyproline II, a secondary structure element implicated in protein-protein interactions. Proteins Struct. Funct. Genet. 2005, 58, 880–892. [Google Scholar] [CrossRef]
- Brown, A.M.; Zondlo, N.J. A propensity scale for type II polyproline helices (PPII): Aromatic amino acids in proline-rich sequences strongly disfavor PPII due to proline-aromatic interactions. Biochemistry 2012, 51, 5041–5051. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, Y.; Forte, J.G. Ezrin oligomers are the membrane-bound dormant form in gastric parietal cells. Am. J. Physiol. Cell Physiol. 2005, 288, C1242–C1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berryman, M.; Gary, R.; Bretscher, A. Ezrin oligomers are major cytoskeletal components of placental microvilli: A proposal for their involvement in cortical morphogenesis. J. Cell Biol. 1995, 131, 1231–1242. [Google Scholar] [CrossRef]
- Bretscher, A.; Gary, R.; Berryman, M. Soluble Ezrin Purified from Placenta Exists as Stable Monomers and Elongated Dimers with Masked C-Terminal Ezrin-Radixin-Moesin Association Domains. Biochemistry 1995, 34, 16830–16837. [Google Scholar] [CrossRef]
- Nguyen, R.; Reczek, D.; Bretscher, A. Hierarchy of Merlin and Ezrin N- and C-terminal Domain Interactions in Homo- and Heterotypic Associations and their Relationship to Binding of Scaffolding Proteins EBP50 and E3KARP. J. Biol. Chem. 2001, 276, 7621–7629. [Google Scholar] [CrossRef]
- Ishikawa, H.; Tamura, A.; Matsui, T.; Sasaki, H.; Hakoshima, T.; Tsukita, S.; Tsukita, S. Structural conversion between open and closed forms of radixin: Low-angle shadowing electron microscopy. J. Mol. Biol. 2001, 310, 973–978. [Google Scholar] [CrossRef]
- Lubart, Q.; Vitet, H.; Dalonneau, F.; Le Roy, A.; Kowalski, M.; Lourdin, M.; Ebel, C.; Weidenhaupt, M.; Picart, C. Role of Phosphorylation in Moesin Interactions with PIP2-Containing Biomimetic Membranes. Biophys. J. 2018, 114, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.-C.; Bertin, A.; Bousquet, H.; Manzi, J.; Senju, Y.; Tsai, M.-C.; Picas, L.; Miserey-Lenkei, S.; Lappalainen, P.; Lemichez, E.; et al. Ezrin enrichment on curved membranes requires a specific conformation or interaction with a curvature-sensitive partner. Elife 2018, 7, e37262. [Google Scholar] [CrossRef]
- Grönholm, M.; Sainio, M.; Zhao, F.; Heiska, L.; Vaheri, A.; Carpén, O. Homotypic and heterotypic interaction of the neurofibromatosis 2 tumor suppressor protein merlin and the ERM protein ezrin. J. Cell Sci. 1999, 112, 895–904. [Google Scholar]
- Meng, J.J.; Lowrie, D.J.; Sun, H.; Dorsey, E.; Pelton, P.D.; Bashour, A.M.; Groden, J.; Ratner, N.; Ip, W. Interaction between two isoforms of the NF2 tumor suppressor protein, merlin, and between merlin and ezrin, suggests modulation of ERM proteins by merlin. J. Neurosci. Res. 2000, 62, 491–502. [Google Scholar] [CrossRef]
- Gonzalez-Agosti, C.; Wiederhold, T.; Herndon, M.E.; Gusella, J.; Ramesh, V. Interdomain interaction of merlin isoforms and its influence on intermolecular binding to NHE-RF. J. Biol. Chem. 1999, 274, 34438–34442. [Google Scholar] [CrossRef]
- Gary, R.; Bretscher, A. Heterotypic and homotypic associations between ezrin and moesin, two putative membrane-cytoskeletal linking proteins. Proc. Natl. Acad. Sci. USA 1993, 90, 10846–10850. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.C.; Molina, J.R.; Hayashi, Y.; Georgescu, M.M. Overexpression of ezrin inactivates NF2 tumor suppressor in glioblastoma. Neuro. Oncol. 2010, 12, 528–539. [Google Scholar] [CrossRef] [Green Version]
- Knight, P.J.; Thirumurugan, K.; Xu, Y.; Wang, F.; Kalverda, A.P.; Stafford, W.F.; Sellers, J.R.; Peckham, M. The predicted coiled-coil domain of myosin 10 forms a novel elongated domain that lengthens the head. J. Biol. Chem. 2005, 280, 34702–34708. [Google Scholar] [CrossRef] [PubMed]
- Fievet, B.T.; Gautreau, A.; Roy, C.; Del Maestro, L.; Mangeat, P.; Louvard, D.; Arpin, M. Phosphoinositide binding and phosphorylation act sequentially in the activation mechanism of ezrin. J. Cell Biol. 2004, 164, 653–659. [Google Scholar] [CrossRef] [Green Version]
- Mori, T.; Kitano, K.; Terawaki, S.I.; Maesaki, R.; Fukami, Y.; Hakoshima, T. Structural basis for CD44 recognition by ERM proteins. J. Biol. Chem. 2008, 283, 29602–29612. [Google Scholar] [CrossRef]
- Terawaki, S.I.; Kitano, K.; Hakoshima, T. Structural basis for type II membrane protein binding by ERM proteins revealed by the radixin-neutral endopeptidase 24.11 (NEP) complex. J. Biol. Chem. 2007, 282, 19854–19862. [Google Scholar] [CrossRef] [PubMed]
- Terawaki, S.; Kitano, K.; Aoyama, M.; Mori, T.; Hakoshima, T. MT1-MMP recognition by ERM proteins and its implication in CD44 shedding. Genes Cells 2015, 20, 847–859. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Li, Y.; Ye, F.; Zhang, M. Structural basis for the phosphorylation-regulated interaction between the cytoplasmic tail of cell polarity protein crumbs and the actin-binding protein moesin. J. Biol. Chem. 2015, 290, 11384–11392. [Google Scholar] [CrossRef] [PubMed]
- Terawaki, S.; Maesaki, R.; Hakoshima, T. Structural basis for NHERF recognition by ERM proteins. Structure 2006, 14, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.C.; Takahashi, Y.; Momin, S.; Adams, H.; Chen, X.; Georgescu, M.-M. NHERF1/EBP50 Head-to-Tail Intramolecular Interaction Masks Association with PDZ Domain Ligands. Mol. Cell. Biol. 2007, 27, 2527–2537. [Google Scholar] [CrossRef]
- Mori, T.; Gotoh, S.; Shirakawa, M.; Hakoshima, T. Structural basis of DDB1-and-Cullin 4-associated Factor 1 (DCAF1) recognition by merlin/NF2 and its implication in tumorigenesis by CD44-mediated inhibition of merlin suppression of DCAF1 function. Genes to Cells 2014, 19, 603–619. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wei, Z.; Zhang, J.; Yang, Z.; Zhang, M. Structural basis of the binding of Merlin FERM domain to the E3 ubiquitin ligase substrate adaptor DCAF1. J. Biol. Chem. 2014, 289, 14674–14681. [Google Scholar] [CrossRef]
- Prag, S.; Parsons, M.; Keppler, M.D.; Ameer-Beg, S.M.; Barber, P.; Hunt, J.; Beavil, A.J.; Calvert, R.; Arpin, M.; Vojnovic, B.; et al. Activated ezrin promotes cell migration through recruitment of the GEF Dbl to lipid rafts and preferential downstream activation of Cdc42. Mol. Biol. Cell 2007, 18, 2935–2948. [Google Scholar] [CrossRef]
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and recalibrations. Nucleic Acids Res. 2015, 43, D512–D520. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2018, 46, 2699. [Google Scholar] [CrossRef]
- Surace, E.I.; Haipek, C.A.; Gutmann, D.H. Effect of merlin phosphorylation on neurofibromatosis 2 (NF2) gene function. Oncogene 2004, 23, 580–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, M.; Potin, S.; Sebbagh, M.; Bertoglio, J.; Breard, J.; Hamelin, J. Rho-ROCK-Dependent Ezrin-Radixin-Moesin Phosphorylation Regulates Fas-Mediated Apoptosis in Jurkat Cells. J. Immunol. 2008, 181, 5963–5973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Jang, S.W.; Wang, X.; Liu, Z.; Bahr, S.M.; Sun, S.Y.; Brat, D.; Gutmann, D.H.; Ye, K. Akt phosphorylation regulates the tumour-suppressor merlin through ubiquitination and degradation. Nat. Cell Biol. 2007, 9, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Maeda, M.; Doi, Y.; Yonemura, S.; Amano, M.; Kaibuchi, K.; Tsukita, S.; Tsukita, S. Rho-kinase phosphorylates COOH-terminal threonines of ezrin/radixin/moesin (ERM) proteins and regulates their head-to-tail association. J. Cell Biol. 1998, 140, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Ye, K. Phosphorylation of merlin regulates its stability and tumor suppressive activity. Cell Adh. Migr. 2007, 1, 196–198. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.C.; Fehon, R.G. Phosphorylation and activity of the tumor suppressor Merlin and the ERM protein Moesin are coordinately regulated by the Slik kinase. J. Cell Biol. 2006, 175, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Schweppe, D.K.; Rigas, J.R.; Gerber, S.A. Quantitative phosphoproteomic profiling of human non-small cell lung cancer tumors. J. Proteom. 2013, 91, 286–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laulajainen, M.; Muranen, T.; Carpén, O.; Grönholm, M. Protein kinase A-mediated phosphorylation of the NF2 tumor suppressor protein merlin at serine 10 affects the actin cytoskeleton. Oncogene 2008, 27, 3233–3243. [Google Scholar] [CrossRef] [PubMed]
- Laulajainen, M.; Muranen, T.; Nyman, T.A.; Carpén, O.; Grönholm, M. Multistep Phosphorylation by Oncogenic Kinases Enhances the Degradation of the NF2 Tumor Suppressor Merlin. Neoplasia 2011, 13, 643–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, R.J.; Paez, J.G.; Curto, M.; Yaktine, A.; Pruitt, W.M.; Saotome, I.; O’Bryan, J.P.; Gupta, V.; Ratner, N.; Der, C.J.; et al. The Nf2 Tumor Suppressor, Merlin, Functions in Rac-Dependent Signaling. Dev. Cell 2001, 1, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Rong, R.; Surace, E.I.; Haipek, C.A.; Gutmann, D.H.; Ye, K. Serine 518 phosphorylation modulates merlin intramolecular association and binding to critical effectors important for NF2 growth suppression. Oncogene 2004, 23, 8447–8454. [Google Scholar] [CrossRef] [Green Version]
- Xing, W.; Li, M.; Zhang, F.; Ma, X.; Long, J.; Zhou, H. The conformation change and tumor suppressor role of Merlin are both independent of Serine 518 phosphorylation. Biochem. Biophys. Res. Commun. 2017, 493, 46–51. [Google Scholar] [CrossRef]
- Ali Khajeh, J.; Ju, J.H.; Atchiba, M.; Allaire, M.; Stanley, C.; Heller, W.T.; Callaway, D.J.; Bu, Z. Molecular conformation of the full-length tumor suppressor NF2/Merlin—a small angle neutron scattering study. J. Mol. Biol. 2014, 426, 2755–2768. [Google Scholar] [CrossRef]
- Guo, X.; Wang, L.; Chen, B.; Li, Q.; Wang, J.; Zhao, M.; Wu, W.; Zhu, P.; Huang, X.; Huang, Q. ERM protein moesin is phosphorylated by advanced glycation end products and modulates endothelial permeability. Am. J. Physiol.-Hear. Circ. Physiol. 2009, 297, H238–H246. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.-S.; Hinds, P.W. Increased ezrin expression and activation by CDK5 coincident with acquisition of the senescent phenotype. Mol. Cell 2003, 11, 1163–1176. [Google Scholar] [CrossRef]
- Adyshev, D.M.; Moldobaeva, N.K.; Elangovan, V.R.; Garcia, J.G.N.; Dudek, S.M. Differential involvement of ezrin/radixin/moesin proteins in sphingosine 1-phosphate-induced human pulmonary endothelial cell barrier enhancement. Cell. Signal. 2011, 23, 2086–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, E.-J.; Ko, H.R.; Hwang, I.; Kim, B.-S.; Choi, J.-Y.; Park, K.W.; Cho, S.-W.; Ahn, J.-Y. Akt regulates neurite growth by phosphorylation-dependent inhibition of radixin proteasomal degradation. Sci. Rep. 2018, 8, 2557. [Google Scholar] [CrossRef] [PubMed]
- Gautreau, A.; Louvard, D.; Arpin, M. Morphogenic effects of ezrin require a phosphorylation-induced transition from oligomers to monomers at the plasma membrane. J. Cell Biol. 2000, 150, 193–203. [Google Scholar] [CrossRef]
- Liu, Y.; Belkina, N.V.; Park, C.; Nambiar, R.; Loughhead, S.M.; Patino-Lopez, G.; Ben-Aissa, K.; Hao, J.-J.; Kruhlak, M.J.; Qi, H.; et al. Constitutively active ezrin increases membrane tension, slows migration, and impedes endothelial transmigration of lymphocytes in vivo in mice. Blood 2012, 119, 445–453. [Google Scholar] [CrossRef]
- Bosk, S.; Braunger, J.A.; Gerke, V.; Steinem, C. Activation of F-actin binding capacity of ezrin: Synergism of PIP2 interaction and phosphorylation. Biophys. J. 2011, 100, 1708–1717. [Google Scholar] [CrossRef]
- Shabardina, V.; Kramer, C.; Gerdes, B.; Braunger, J.; Cordes, A.; Schäfer, J.; Mey, I.; Grill, D.; Gerke, V.; Steinem, C. Mode of Ezrin-Membrane Interaction as a Function of PIP2 Binding and Pseudophosphorylation. Biophys. J. 2016, 110, 2710–2719. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Stamler, J.S. Enzymatic mechanisms regulating protein s-nitrosylation: Implications in health and disease. J. Mol. Med. 2012, 90, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Arif, A.; Terenzi, F.; Willard, B.; Plow, E.F.; Hazen, S.L.; Fox, P.L. Target-selective protein S-nitrosylation by sequence motif recognition. Cell 2014, 159, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Wolhuter, K.; Eaton, P. How widespread is stable protein S-nitrosylation as an end-effector of protein regulation? Free Radic. Biol. Med. 2017, 109, 156–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaarour, R.F.; Chirivino, D.; Del Maestro, L.; Daviet, L.; Atfi, A.; Louvard, D.; Arpin, M. Ezrin ubiquitylation by the E3 ubiquitin ligase, WWP1, and consequent regulation of hepatocyte growth factor receptor activity. PLoS ONE 2012, 7, e37490. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; You, L.; Cooper, J.; Schiavon, G.; Pepe-Caprio, A.; Zhou, L.; Ishii, R.; Giovannini, M.; Hanemann, C.O.; Long, S.B.; et al. Merlin/NF2 Suppresses Tumorigenesis by Inhibiting the E3 Ubiquitin Ligase CRL4DCAF1 in the Nucleus. Cell 2010, 140, 477–490. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, C.; Kumar, C.; Gnad, F.; Nielsen, M.L.; Rehman, M.; Walther, T.C.; Olsen, J.V.; Mann, M. Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science 2009, 325, 834–840. [Google Scholar] [CrossRef]
- Park, J.; Chen, Y.; Tishkoff, D.X.; Peng, C.; Tan, M.; Dai, L.; Xie, Z.; Zhang, Y.; Zwaans, B.M.M.; Skinner, M.E.; et al. SIRT5-Mediated Lysine Desuccinylation Impacts Diverse Metabolic Pathways. Mol. Cell 2013, 50, 919–930. [Google Scholar] [CrossRef]
- Wang, X.; Herr, R.A.; Chua, W.J.; Lybarger, L.; Wiertz, E.J.H.J.; Hansen, T.H. Ubiquitination of serine, threonine, or lysine residues on the cytoplasmic tail can induce ERAD of MHC-I by viral E3 ligase mK3. J. Cell Biol. 2007, 177, 613–624. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, Y.; Okuda-Shimizu, Y.; Hendershot, L.M. Ubiquitylation of an ERAD Substrate Occurs on Multiple Types of Amino Acids. Mol. Cell 2010, 40, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A.; Ben-Saadon, R. N-terminal ubiquitination: More protein substrates join in. Trends Cell Biol. 2004, 14, 103–106. [Google Scholar] [CrossRef]
- Qi, Q.; Liu, X.; Brat, D.J.; Ye, K. Merlin sumoylation is required for its tumor suppressor activity. Oncogene 2014, 33, 4893–4903. [Google Scholar] [CrossRef]
- Nie, M.; Xie, Y.; Loo, J.A.; Courey, A.J. Genetic and proteomic evidence for roles of Drosophila SUMO in cell cycle control, Ras signaling, and early pattern formation. PLoS ONE 2009, 4, e5905. [Google Scholar] [CrossRef]
- Weinert, B.T.; Schölz, C.; Wagner, S.A.; Iesmantavicius, V.; Su, D.; Daniel, J.A.; Choudhary, C. Lysine succinylation is a frequently occurring modification in prokaryotes and eukaryotes and extensively overlaps with acetylation. Cell Rep. 2013, 4, 842–851. [Google Scholar] [CrossRef]
- Song, Y.; Wang, J.; Cheng, Z.; Gao, P.; Sun, J.; Chen, X.; Chen, C.; Wang, Y.; Wang, Z. Quantitative global proteome and lysine succinylome analyses provide insights into metabolic regulation and lymph node metastasis in gastric cancer. Sci. Rep. 2017, 7, 42053. [Google Scholar] [CrossRef] [Green Version]
- Maresso, A.W.; Deng, Q.; Pereckas, M.S.; Wakim, B.T.; Barbieri, J.T. Pseudomonas aeruginosa ExoS ADP-ribosyltransferase inhibits ERM phosphorylation. Cell. Microbiol. 2007, 9, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maresso, A.W.; Baldwin, M.R.; Barbieri, J.T. Ezrin/radixin/moesin proteins are high affinity targets for ADP-ribosylation by Pseudomonas aeruginosa ExoS. J. Biol. Chem. 2004, 279, 38402–38408. [Google Scholar] [CrossRef] [PubMed]
- Millet, J.K.; Nal, B. Investigation of the functional roles of host cell proteins involved in coronavirus infection using highly specific and scalable rna interference (RNAi) approach. In Coronaviruses: Methods and Protocols; Humana Press: New York, NY, USA, 2015. [Google Scholar]
- Kamiyama, H.; Izumida, M.; Umemura, Y.; Hayashi, H.; Matsuyama, T.; Kubo, Y. Role of ezrin phosphorylation in HIV-1 replication. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Nuesch, J.P.F.; Bar, S.; Lachmann, S.; Rommelaere, J. Ezrin-Radixin-Moesin Family Proteins Are Involved in Parvovirus Replication and Spreading. J. Virol. 2009, 83, 5854–5863. [Google Scholar] [CrossRef]
- Niggli, V.; Andréoli, C.; Roy, C.; Mangeat, P. Identification of a phosphatidylinositol-4,5-bisphosphate-binding domain in the N-terminal region of ezrin. FEBS Lett. 1995, 376, 172–176. [Google Scholar] [CrossRef] [Green Version]
- Mani, T.; Hennigan, R.F.; Foster, L.A.; Conrady, D.G.; Herr, A.B.; Ip, W. FERM domain phosphoinositide binding targets merlin to the membrane and is essential for its growth-suppressive function. Mol. Cell. Biol. 2011, 31, 1983–1996. [Google Scholar] [CrossRef] [PubMed]
- Blin, G.; Margeat, E.; Carvalho, K.; Royer, C.A.; Roy, C.; Picart, C. Quantitative analysis of the binding of ezrin to large unilamellar vesicles containing phosphatidylinositol 4,5 bisphosphate. Biophys. J. 2008, 94, 1021–1033. [Google Scholar] [CrossRef] [PubMed]
- Maniti, O.; Carvalho, K.; Picart, C. Model membranes to shed light on the biochemical and physical properties of ezrin/radixin/moesin. Biochimie 2013, 95, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Wang, Y.; Jang, S.W.; Tang, X.; Neri, L.M.; Ye, K. Akt phosphorylation of merlin enhances its binding to phosphatidylinositols and inhibits the tumor-suppressive activities of merlin. Cancer Res. 2009, 69, 4043–4051. [Google Scholar] [CrossRef] [PubMed]
- Yonemura, S.; Matsui, T.; Tsukita, S.; Tsukita, S. Rho-dependent and -independent activation mechanisms of ezrin/radixin/moesin proteins: An essential role for polyphosphoinositides in vivo. J. Cell Sci. 2002, 115, 2569–2580. [Google Scholar]
- Hao, J.J.; Liu, Y.; Kruhlak, M.; Debell, K.E.; Rellahan, B.L.; Shaw, S. Phospholipase C-mediated hydrolysis of PIP2 releases ERM proteins from lymphocyte membrane. J. Cell Biol. 2009, 184, 451–462. [Google Scholar] [CrossRef]
- Ben-Aissa, K.; Patino-Lopez, G.; Belkina, N.V.; Maniti, O.; Rosales, T.; Hao, J.-J.; Kruhlak, M.J.; Knutson, J.R.; Picart, C.; Shaw, S. Activation of moesin, a protein that links actin cytoskeleton to the plasma membrane, occurs by phosphatidylinositol 4,5-bisphosphate (PIP2) binding sequentially to two sites and releasing an autoinhibitory linker. J. Biol. Chem. 2012, 287, 16311–16323. [Google Scholar] [CrossRef]
- Senju, Y.; Kalimeri, M.; Koskela, E.V.; Somerharju, P.; Zhao, H.; Vattulainen, I.; Lappalainen, P. Mechanistic principles underlying regulation of the actin cytoskeleton by phosphoinositides. Proc. Natl. Acad. Sci. USA 2017, 114, E8977–E8986. [Google Scholar] [CrossRef]
- Barret, C.; Roy, C.; Montcourrier, P.; Mangeat, P.; Niggli, V. Mutagenesis of the phosphatidylinositol 4,5-bisphosphate (PIP(2)) binding site in the NH(2)-terminal domain of ezrin correlates with its altered cellular distribution. J. Cell Biol. 2000, 151, 1067–1080. [Google Scholar] [CrossRef]
- Carvalho, K.; Khalifat, N.; Maniti, O.; Nicolas, C.; Arold, S.; Picart, C.; Ramos, L. Phosphatidylinositol 4,5-bisphosphate-induced conformational change of ezrin and formation of ezrin oligomers. Biochemistry 2010, 49, 9318–9327. [Google Scholar] [CrossRef]
- Maniti, O.; Khalifat, N.; Goggia, K.; Dalonneau, F.; Guérin, C.; Blanchoin, L.; Ramos, L.; Picart, C. Binding of moesin and ezrin to membranes containing phosphatidylinositol (4,5) bisphosphate: A comparative study of the affinity constants and conformational changes. Biochim. Biophys. Acta 2012, 1818, 2839–2849. [Google Scholar] [CrossRef]
- Jayasundar, J.J.; Ju, J.H.; He, L.; Liu, D.; Meilleur, F.; Zhao, J.; Callaway, D.J.E.; Bu, Z. Open conformation of ezrin bound to phosphatidylinositol 4,5-bisphosphate and to F-actin revealed by neutron scattering. J. Biol. Chem. 2012, 287, 37119–37133. [Google Scholar] [CrossRef] [PubMed]
- Heiska, L.; Alfthan, K.; Grönholm, M.; Vilja, P.; Vaheri, A.; Carpén, O. Association of ezrin with intercellular adhesion molecule-1 and -2 (ICAM-1 and ICAM-2). Regulation by phosphatidylinositol 4, 5-bisphosphate. J. Biol. Chem. 1998, 273, 21893–21900. [Google Scholar] [CrossRef]
- Hirao, M.; Sato, N.; Kondo, T.; Yonemura, S.; Monden, M.; Sasaki, T.; Takai, Y.; Tsukita, S.; Tsukita, S. Regulation mechanism of ERM (ezrin/radixin/moesin) protein/plasma membrane association: Possible involvement of phosphatidylinositol turnover and Rho-dependent signaling pathway. J. Cell Biol. 1996, 135, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Solinet, S.; Mahmud, K.; Stewman, S.F.; Ben El Kadhi, K.; Decelle, B.; Talje, L.; Ma, A.; Kwok, B.H.; Carreno, S. The actin-binding ERM protein Moesin binds to and stabilizes microtubules at the cell cortex. J. Cell Biol. 2013, 202, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.M.; Gutmann, D.H. Merlin differentially associates with the microtubule and actin cytoskeleton. J. Neurosci. Res. 1998, 51, 403–415. [Google Scholar] [CrossRef]
- Bensenor, L.B.; Barlan, K.; Rice, S.E.; Fehon, R.G.; Gelfand, V.I. Microtubule-mediated transport of the tumor-suppressor protein Merlin and its mutants. Proc. Natl. Acad. Sci. USA 2010, 107, 7311–7316. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Cooper, J.; Zhou, L.; Yang, C.; Erdjument-Bromage, H.; Zagzag, D.; Snuderl, M.; Ladanyi, M.; Hanemann, C.O.; Zhou, P.; et al. Merlin/NF2 Loss-Driven Tumorigenesis Linked to CRL4DCAF1-Mediated Inhibition of the Hippo Pathway Kinases Lats1 and 2 in the Nucleus. Cancer Cell 2014, 26, 48–60. [Google Scholar] [CrossRef]
- Fung, H.Y.J.; Fu, S.C.; Chook, Y.M. Nuclear export receptor CRM1 recognizes diverse conformations in nuclear export signals. Elife 2017, 6, 1–13. [Google Scholar] [CrossRef]
- Kressel, M.; Schmucker, B. Nucleocytoplasmic transfer of the NF2 tumor suppressor protein merlin is regulated by exon 2 and a CRM1-dependent nuclear export signal in exon 15. Hum. Mol. Genet. 2002, 11, 2269–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, K.T.; Yamashita, K.; Sakurai, N.; Ohno, S. The Epithelial Circumferential Actin Belt Regulates YAP/TAZ through Nucleocytoplasmic Shuttling of Merlin. Cell Rep. 2017, 20, 1435–1447. [Google Scholar] [CrossRef]
- Turunen, O.; Wahlström, T.; Vaheri, A. Ezrin has a COOH-terminal actin-binding site that is conserved in the ezrin protein family. J. Cell Biol. 1994, 126, 1445–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gary, R.; Bretscher, A. Ezrin self-association involves binding of an N-terminal domain to a normally masked C-terminal domain that includes the F-actin binding site. Mol. Biol. Cell 1995, 6, 1061–1075. [Google Scholar] [CrossRef]
- Saleh, H.S.; Merkel, U.; Geissler, K.J.; Sperka, T.; Sechi, A.; Breithaupt, C.; Morrison, H. Properties of an Ezrin Mutant Defective in F-actin Binding. J. Mol. Biol. 2009, 385, 1015–1031. [Google Scholar] [CrossRef] [PubMed]
- Shuster, C.B.; Herman, I.M. Indirect association of ezrin with F-actin: Isoform specificity and calcium sensitivity. J. Cell Biol. 1995, 128, 837–848. [Google Scholar] [CrossRef]
- Bretscher, A. Purification of an 80,000-dalton protein that is a component of the isolated microvillus cytoskeleton, and its localization in nonmuscle cells. J. Cell Biol. 1983, 97, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Pestonjamasp, K.; Amieva, M.R.; Strassel, C.P.; Nauseef, W.M.; Furthmayr, H.; Luna, E.J. Moesin, ezrin, and p205 are actin-binding proteins associated with neutrophil plasma membranes. Mol. Biol. Cell 1995, 6, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, F.; Huang, L.; Pestonjamasp, K.; Luna, E.J.; Furthmayr, H. Regulation of F-actin binding to platelet moesin in vitro by both phosphorylation of threonine 558 and polyphosphatidylinositides. Mol. Biol. Cell 1999, 10, 2669–2685. [Google Scholar] [CrossRef] [PubMed]
- Janke, M.; Herrig, A.; Austermann, J.; Gerke, V.; Steinem, C.; Janshoff, A. Actin binding of ezrin is activated by specific recognition of PIP2-functionalized lipid bilayers. Biochemistry 2008, 47, 3762–3769. [Google Scholar] [CrossRef]
- Sainio, M.; Zhao, F.; Heiska, L.; Turunen, O.; den Bakker, M.; Zwarthoff, E.; Lutchman, M.; Rouleau, G.A.; Jääskeläinen, J.; Vaheri, A.; et al. Neurofibromatosis 2 tumor suppressor protein colocalizes with ezrin and CD44 and associates with actin-containing cytoskeleton. J. Cell Sci. 1997, 110, 2249–2260. [Google Scholar] [PubMed]
- James, M.F.; Manchanda, N.; Gonzalez-Agosti, C.; Hartwig, J.H.; Ramesh, V. The neurofibromatosis 2 protein product merlin selectively binds F-actin but not G-actin, and stabilizes the filaments through a lateral association. Biochem. J. 2001, 356, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Baboolal, T.G.; Siththanandan, V.; Chen, M.; Walker, M.L.; Knight, P.J.; Peckham, M.; Sellers, J.R. A FERM domain autoregulates Drosophila myosin 7a activity. Proc. Natl. Acad. Sci. USA 2009, 106, 4189–4194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-S.; Bellin, R.M.; Walker, D.L.; Patel, B.; Powers, P.; Liu, H.; Garcia-Alvarez, B.; de Pereda, J.M.; Liddington, R.C.; Volkmann, N.; et al. Characterization of an actin-binding site within the talin FERM domain. J. Mol. Biol. 2004, 343, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Scoles, D.R.; Huynh, D.P.; Morcos, P.A.; Coulsell, E.R.; Robinson, N.G.G.; Tamanoi, F.; Pulst, S.M. Neurofibromatosis 2 tumour suppressor schwannomin interacts with βII- spectrin. Nat. Genet. 1998, 18, 354–359. [Google Scholar] [CrossRef]
- Neill, G.W.; Crompton, M.R. Binding of the merlin-I product of the neurofibromatosis type 2 tumour suppressor gene to a novel site in beta-fodrin is regulated by association between merlin domains. Biochem J. 2001, 358, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Yogesha, S.D.; Sharff, A.J.; Giovannini, M.; Bricogne, G.; Izard, T. Unfurling of the band 4.1, ezrin, radixin, moesin (FERM) domain of the merlin tumor suppressor. Protein Sci. 2011, 20, 2113–2120. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Merlin-ERM | ERM only | |
|---|---|---|
| Full-length | 37% (226/606) | 67% (394/586) |
| FERM domain (Merlin: 1–312; ERM: 1–296) | 54% (169/313) | 81% (240/296) |
| Helical domain (Merlin: 313–478; ERM: 297–469) | 23% (39/173) | 56% (99/173) |
| C-terminal domain (including polyPro tract) (Merlin: 479–595; ERM: 470-C-terminus) | 15% (15/126) | 48% (57/118) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michie, K.A.; Bermeister, A.; Robertson, N.O.; Goodchild, S.C.; Curmi, P.M.G. Two Sides of the Coin: Ezrin/Radixin/Moesin and Merlin Control Membrane Structure and Contact Inhibition. Int. J. Mol. Sci. 2019, 20, 1996. https://doi.org/10.3390/ijms20081996
Michie KA, Bermeister A, Robertson NO, Goodchild SC, Curmi PMG. Two Sides of the Coin: Ezrin/Radixin/Moesin and Merlin Control Membrane Structure and Contact Inhibition. International Journal of Molecular Sciences. 2019; 20(8):1996. https://doi.org/10.3390/ijms20081996
Chicago/Turabian StyleMichie, Katharine A., Adam Bermeister, Neil O. Robertson, Sophia C. Goodchild, and Paul M. G. Curmi. 2019. "Two Sides of the Coin: Ezrin/Radixin/Moesin and Merlin Control Membrane Structure and Contact Inhibition" International Journal of Molecular Sciences 20, no. 8: 1996. https://doi.org/10.3390/ijms20081996
APA StyleMichie, K. A., Bermeister, A., Robertson, N. O., Goodchild, S. C., & Curmi, P. M. G. (2019). Two Sides of the Coin: Ezrin/Radixin/Moesin and Merlin Control Membrane Structure and Contact Inhibition. International Journal of Molecular Sciences, 20(8), 1996. https://doi.org/10.3390/ijms20081996