The Interaction between 30b-5p miRNA and MBNL1 mRNA is Involved in Vascular Smooth Muscle Cell Differentiation in Patients with Coronary Atherosclerosis

 , ,
, ,
Abstract
:1. Introduction
2. Results
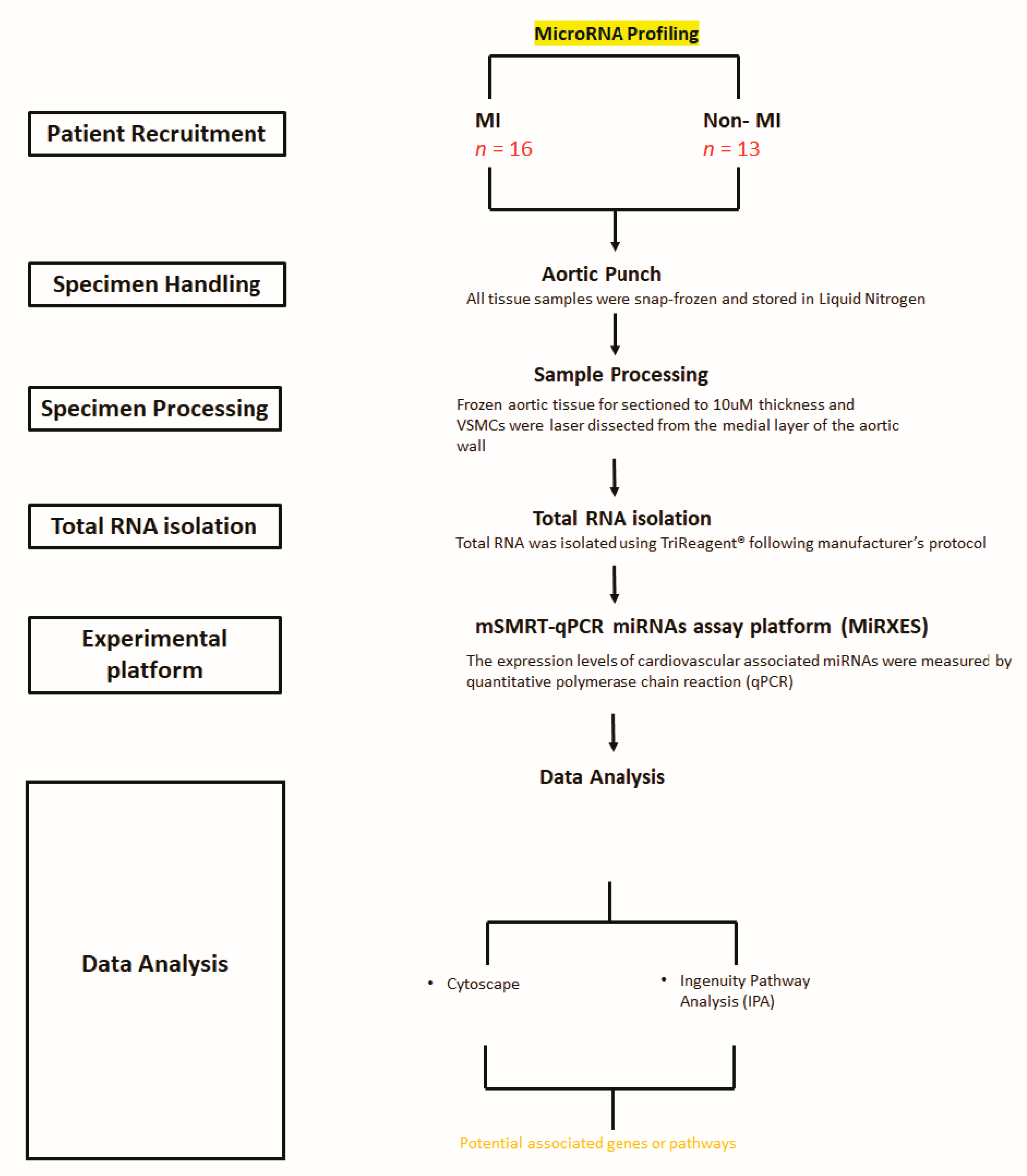
2.1. Demographic Characteristics of the Patients Recruited for This Study
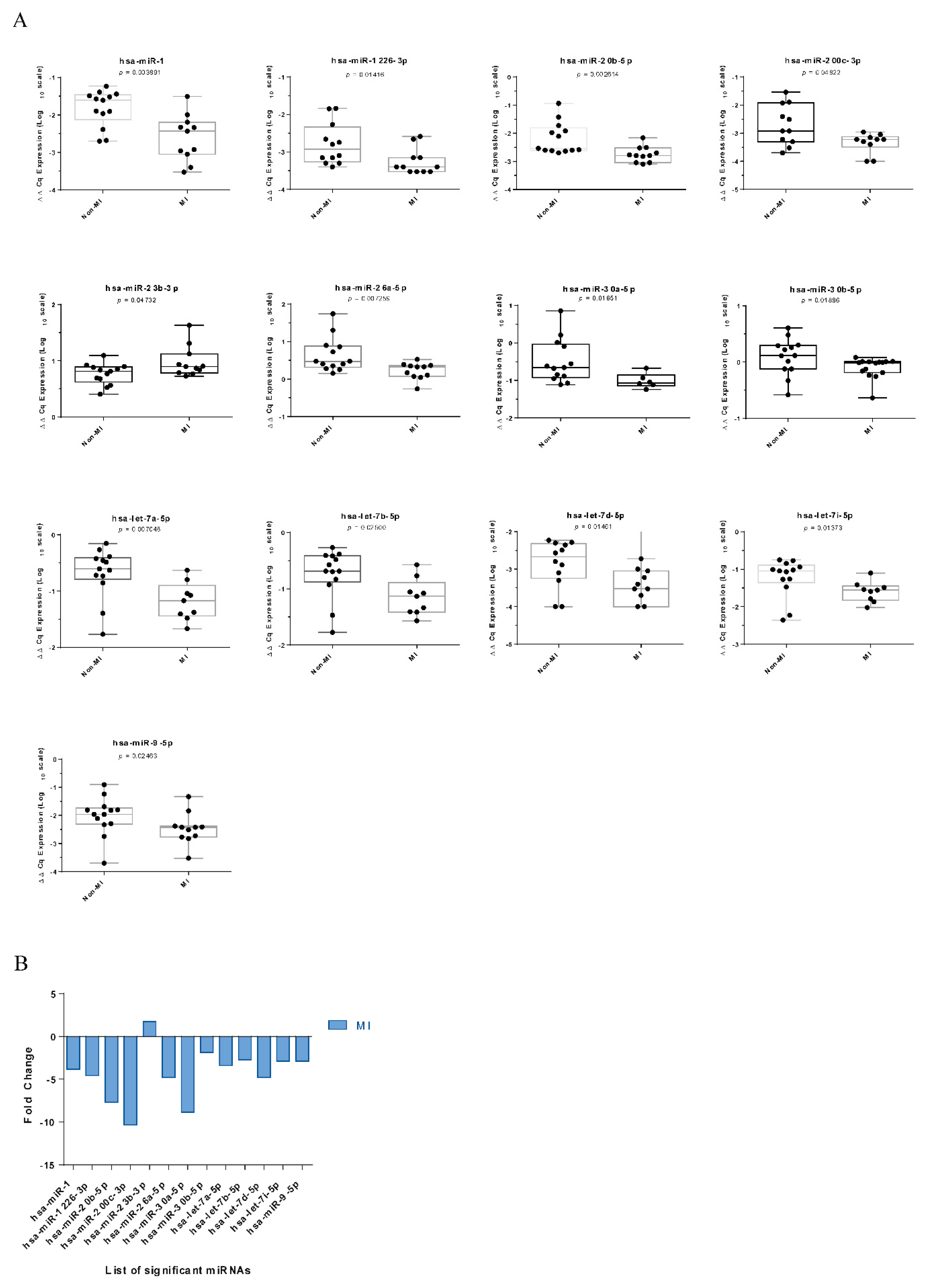
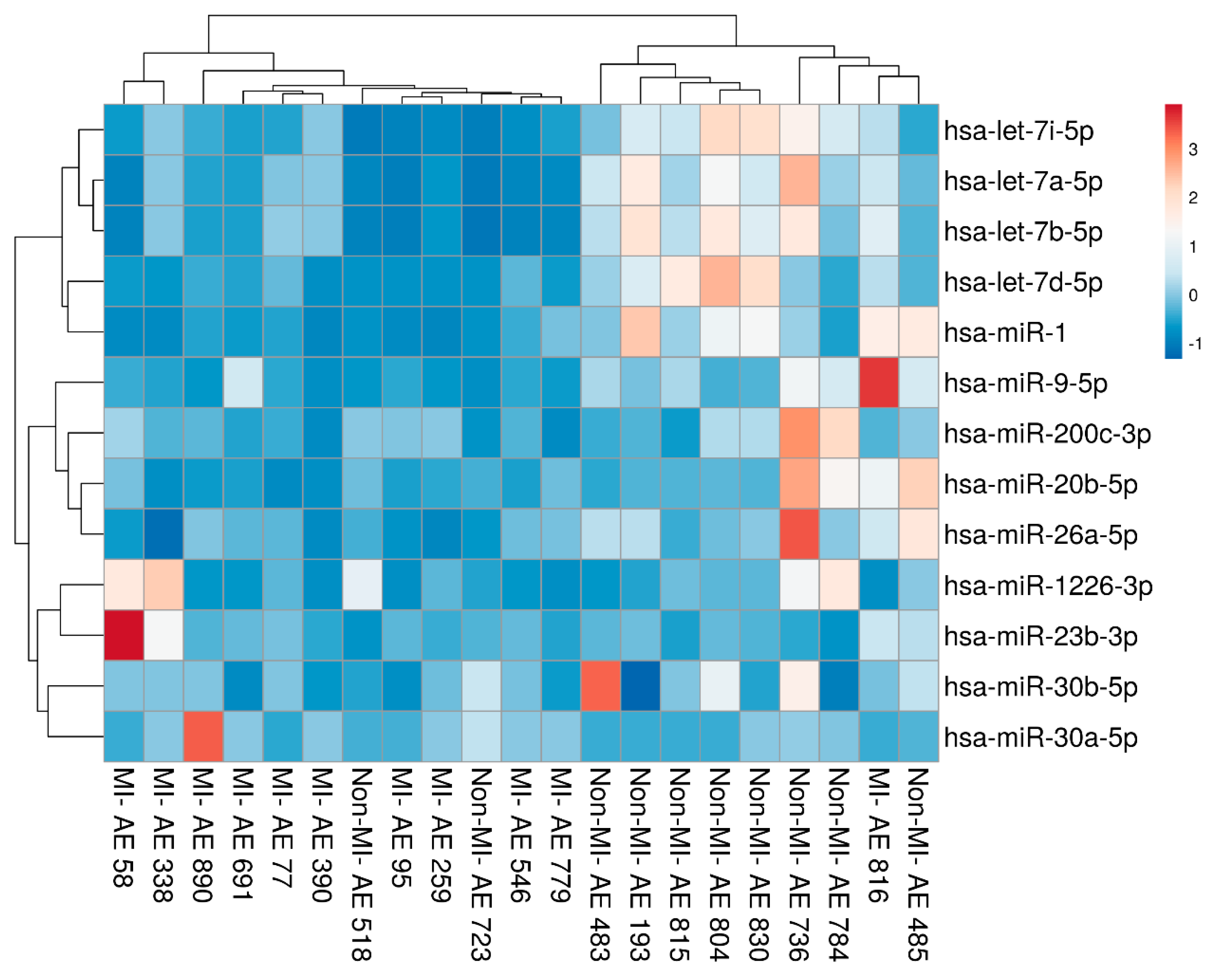
2.2. Characteristics of miRNAs in VSMCs from the Atherosclerotic Wall
2.3. Target mRNA Prediction for Differentially Expressed miRNAs
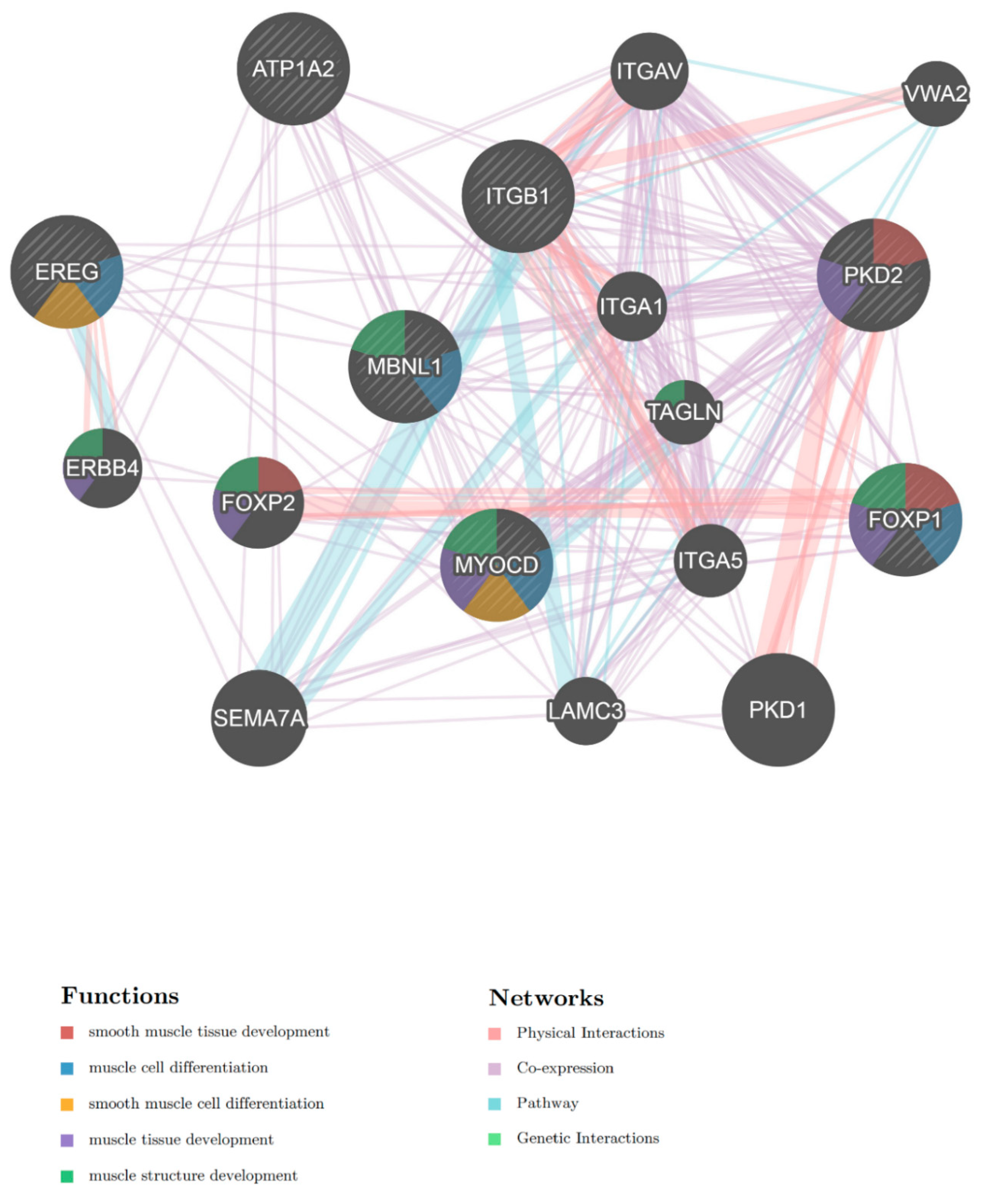
2.4. Biological Function Analysis of Predicted miRNA Target Genes
2.5. Analysis of Predicted miRNA Target mRNAs Associated with Muscle Cell Differentiation
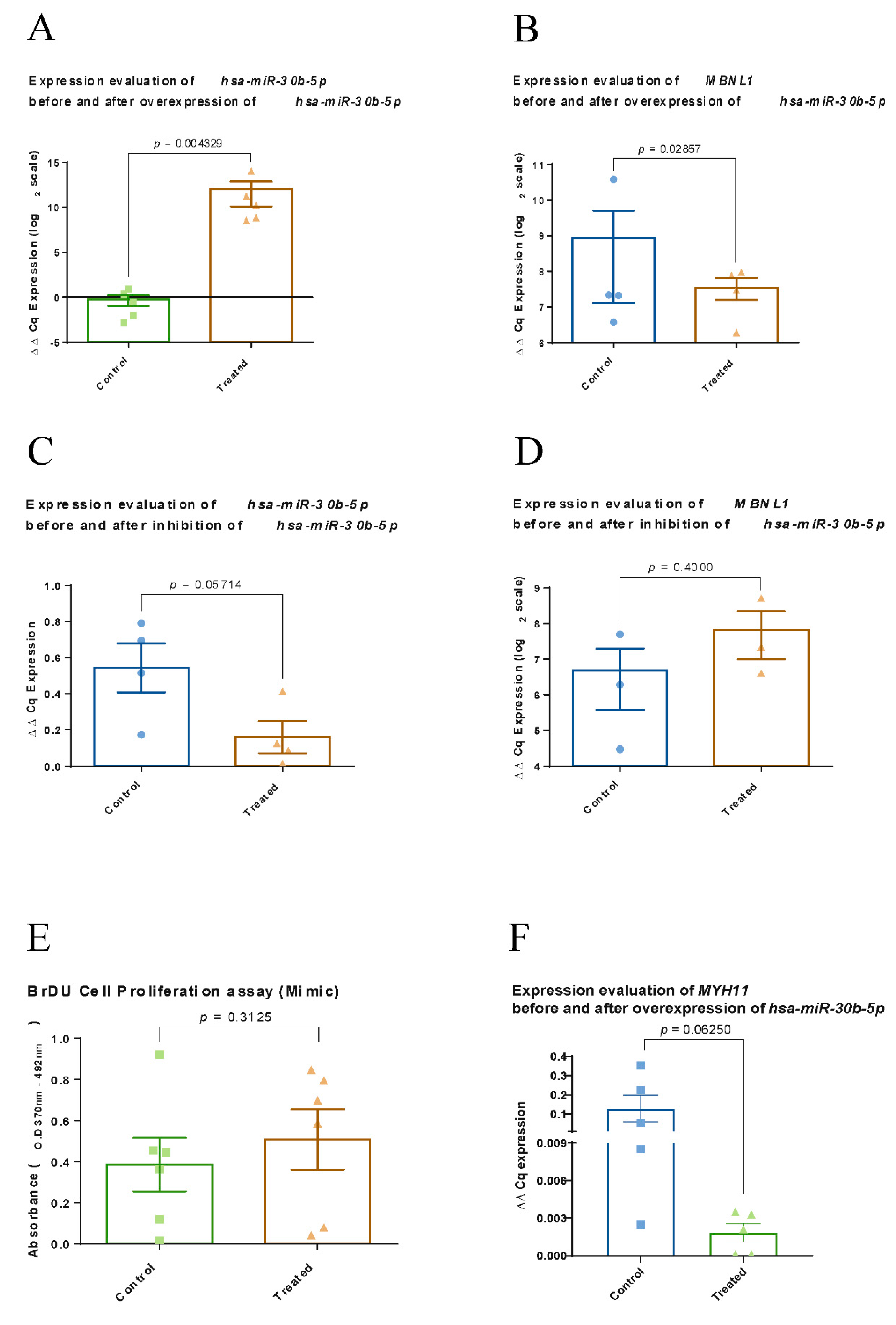
2.6. Experimental Evaluation of Regulatory Function of hsa-miR-30b-5p on MBNL1 in Human VSMC
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Aortic Wall Tissue Collection
4.3. Cryosectioning, Staining, and LCM of VSMC
4.4. RNA Processing and miRNA Profiling, Heatmapping, and Clustering
4.5. Transfection of miRNA Oligonucleotides
4.6. Data Processing and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| VSMC | Vascular smooth muscle cells |
| miRNAs | microRNAs |
| SM | Smooth muscle |
| CAD | Coronary artery disease |
| MI | Myocardial infarction |
| RT-qPCR | Real-time polymerase chain reaction |
| IPA | Ingenuity Pathway Analysis |
| DIANA | DNA Intelligent Analysis |
| miEAA | miRNA Enrichment Analysis and Annotation |
| GO | Gene ontology |
| SMC | Smooth muscle cell |
| BrdU | Bromodeoxyuridine |
| MYOCD | Myocardin |
| MBNL1 | Muscleblind-like splicing regulator 1 |
| LCM | Laser capture microdissection |
References
- Michel, J.B.; Li, Z.; Lacolley, P. Smooth muscle cells and vascular diseases. Cardiovasc. Res. 2012, 95, 135–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Vascular smooth muscle cell in atherosclerosis. Acta Physiol. 2015, 214, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Hata, A. MicroRNA regulation of smooth muscle gene expression and phenotype. Curr. Opin. Hematol. 2012, 19, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.C.; Wongsurawat, T.; Soong, R.; Lee, C.N.; Richards, M.; Kuznetsov, V.; Sorokin, V. Distinctive pattern of LET-7B and MIR-30B in human aortic smooth muscle cells of myocardial infarction patients. Atherosclerosis 2017, 263, e63. [Google Scholar] [CrossRef]
- Wongsurawat, T.; Woo, C.C.; Giannakakis, A.; Lin, X.Y.; Cheow, E.S.H.; Lee, C.N.; Richards, M.; Sze, S.K.; Nookaew, I.; Kuznetsov, V.A.; et al. Distinctive molecular signature and activated signaling pathways in aortic smooth muscle cells of patients with myocardial infarction. Atherosclerosis 2018, 271, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.N.; Xie, B.D.; Sun, L.; Chen, W.; Jiang, S.L.; Liu, W.; Bian, F.; Tian, H.; Li, R.K. Phenotypic switching of vascular smooth muscle cells in the ‘normal region’ of aorta from atherosclerosis patients is regulated by miR-145. J. Cell Mol. Med. 2016, 20, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Gomez, D.; Owens, G.K. Smooth muscle cell phenotypic switching in atherosclerosis. Cardiovasc. Res. 2012, 95, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Rensen, S.S.M.; Doevendans, P.A.; van Eys, G.J.J.M. Regulation and characteristics of vascular smooth muscle cell phenotypic diversity. Neth. Heart J. 2007, 15, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.W.; Cai, H.F.; Wei, X.F.; Sun, J.J.; Lan, X.Y.; Lei, C.Z.; Lin, F.P.; Qi, X.L.; Plath, M.; Chen, H. miR-30-5p regulates muscle differentiation and alternative splicing of muscle-related genes by targeting MBNL. Int. J. Mol. Sci. 2016, 17, E182. [Google Scholar] [CrossRef] [Green Version]
- Jamaluddin, M.S.; Weakley, S.M.; Zhang, L.; Kougias, P.; Lin, P.H.; Yao, Q.; Chen, C. miRNAs: Roles and clinical applications in vascular disease. Expert Rev. Mol. Diagn. 2011, 11, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, M.D.; Georgakilas, G.; Kostoulas, N.; Vlachos, I.S.; Vergoulis, T.; Reczko, M.; Filippidis, C.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-microT web server v5.0: Service integration into miRNA functional analysis workflows. Nucleic Acids Res. 2013, 41, W169–W173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backes, C.; Khaleeq, Q.T.; Meese, E.; Keller, A. miEAA: microRNA enrichment analysis and annotation. Nucleic Acids Res. 2016, 44, W110–W116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Song, Z.; Li, G. Role of specific microRNAs in regulation of vascular smooth muscle cell differentiation and the response to injury. J. Cardiovasc. Transl. Res. 2010, 3, 246–250. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Chen, Q.; Wen, G.; Zhang, C.; Luong, L.A.; Zhu, J.; Zhang, L.; Xiao, Q. Functional role of miRNA-34A in smooth muscle cell functions and neointima formation. Atherosclerosis 2016, 244, e3–e4. [Google Scholar] [CrossRef]
- Davis-Dusenbery, B.N.; Wu, C.; Hata, A. Micromanaging vascular smooth muscle cell differentiation and phenotypic modulation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2370–2377. [Google Scholar] [CrossRef] [Green Version]
- Albinsson, S.; Sessa, W.C. Can microRNAs control vascular smooth muscle phenotypic modulation and the response to injury? Physiol. Genom. 2011, 43, 529–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabas, I. Macrophage death and defective inflammation resolution in atherosclerosis. Nat. Rev. Immunol. 2010, 10, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular smooth muscle cells in atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Makki, N.; Thiel, W.K.; Miller, F.J., Jr. The epidermal growth factor receptor and its ligands in cardiovascular disease. Int. J. Mol. Sci. 2013, 14, 20597–20613. [Google Scholar] [CrossRef] [Green Version]
- Dixon, D.M.; Choi, J.; El-Ghazali, A.; Park, S.Y.; Roos, K.P.; Jordan, M.C.; Fishbein, M.C.; Comai, L.; Reddy, S. Loss of muscleblind-like 1 results in cardiac pathology and persistence of embryonic splice isoforms. Sci. Rep. 2015, 5, 9042. [Google Scholar] [CrossRef]
- Takahashi, M.; Hayashi, K.; Yoshida, K.; Ohkawa, Y.; Komurasaki, T.; Kitabatake, A.; Ogawa, A.; Nishida, W.; Yano, M.; Monden, M.; et al. Epiregulin as a major autocrine/paracrine factor released from ERK- and p38MAPK-activated vascular smooth muscle cells. Circulation 2003, 108, 2524–2529. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, D.Z.; Tegs Pipes, G.C.; Olson, E.N. Myocardin is a master regulator of smooth muscle gene expression. Proc. Natl. Acad. Sci. USA 2003, 100, 7129–7134. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.L. Myocardin and smooth muscle differentiation. Arch. Biochem. Biophys. 2014, 543, 48–56. [Google Scholar] [CrossRef]
- Yoshida, T.; Sinha, S.; Dandré, F.; Wamhoff, B.R.; Hoofnagle, M.H.; Kremer, B.E.; Wang, D.Z.; Olson, E.N.; Owens, G.K. Myocardin is a key regulator of CArG-dependent transcription of multiple smooth muscle marker genes. Circ. Res. 2003, 92, 856–864. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, S.; Yuan, L.; Tian, Y.; Weidenfeld, J.; Yang, J.; Liu, F.; Chokas, A.L.; Morrisey, E.E. Foxp1 coordinates cardiomyocyte proliferation through both cell-autonomous and nonautonomous mechanisms. Genes Dev. 2010, 24, 1746–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arunachalam, G.; Upadhyay, R.; Ding, H.; Triggle, C.R. MicroRNA signature and cardiovascular dysfunction. J. Cardiovasc. Pharmacol. 2015, 65, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, D.Z. microRNAs in cardiovascular development. J. Mol. Cell Cardiol. 2012, 52, 949–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Liu, X.; Yang, J.; Lin, Y.; Xu, D.Z.; Lu, Q.; Deitch, E.A.; Huo, Y.; Delphin, E.S.; Zhang, C. MicroRNA-145, a novel smooth muscle cell phenotypic marker and modulator, controls vascular neointimal lesion formation. Circ. Res. 2009, 105, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Ranganna, K.; Mathew, O.P.; Milton, S.G.; Hayes, B.E. MicroRNAome of vascular smooth muscle cells: Potential for microRNA-based vascular therapies. In Current Trends in Atherogenesis; Rezzani, R., Ed.; IntechOpen Limited: London, UK, 2013; pp. 147–164. [Google Scholar]
- Pujol-López, M.; Ortega-Paz, L.; Garabito, M.; Brugaletta, S.; Sabaté, M.; Dantas, A.P. miRNA update: A review focus on clinical implications of miRNA in vascular remodeling. AIMS Med Sci. 2017, 4, 99–112. [Google Scholar] [CrossRef]
- Suzuki, R.; Shimodaira, H. Pvclust: An R package for assessing the uncertainty in hierarchical clustering. Bioinformatics 2006, 22, 1540–1542. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. Clustvis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | MI (n = 16) | Non-MI (n = 13) | p-Value | |
|---|---|---|---|---|
| Ethnicity | Chinese (%) | 12 (75.0%) | 7 (53.8%) | 0.363 |
| Malay (%) | 3 (18.8%) | 3 (23.1%) | ||
| Others (%) | 1 (6.3%) | 3 (23.1%) | ||
| Gender | Male (%) | 14 (87.5%) | 13 (100%) | 0.186 |
| Female (%) | 2 (12.5%) | 0 (0%) | ||
| Age (Mean ± SD) | 62.19 ± 9.614 | 55.85 ± 6.401 | 0.052 | |
| Diabetes Mellitus | No (%) | 6 (37.5%) | 5 (38.5%) | 0.958 |
| Yes (%) | 10 (62.5%) | 8 (61.5%) | ||
| Hypertension | No (%) | 1 (6.3%) | 1 (7.7%) | 0.879 |
| Yes (%) | 15 (93.8%) | 12 (92.3%) | ||
| Hyperlipidemia | No (%) | 0 (0%) | 1 (7.7%) | 0.259 |
| Yes (%) | 16 (100%) | 12 (92.3%) | ||
| Smoking | No (%) | 7 (43.8%) | 6 (46.2%) | 0.897 |
| Yes (%) | 9 (56.2%) | 7 (53.8%) | ||
| Ejection Fraction | Good (>45%) | 8 (50.0%) | 9 (69.2%) | 0.579 |
| Fair (30–45%) | 6 (37.5%) | 3 (23.1%) | ||
| Poor (<30%) | 2 (12.5%) | 1 (7.7%) | ||
| Troponin I (µg/L) (Mean ± SD) | 13.73 ± 4.768 | 3.377 ± 1.834 | 0.003 | |
| GOID | GO Term | Term p-value | Term p-value Corrected with Benjamini-Hochberg | % Associated | Associated mRNA |
|---|---|---|---|---|---|
| GO:0060537 | muscle tissue development | 0.000000 | 0.000021 | 5.56 | ACADM, AKAP6, ARID2, COL11A1, CREB1, DDX5, FOXP1, HNRNPU, HOMER1, IGF1, ITGB1, LEMD2, MEF2D, MYOCD, NLN, NR1D2, PDGFRA, PKD2, PRKAA1, RBFOX1, RPS6KB1, SKIL] |
| GO:0060537 | muscle tissue development | 0.000000 | 0.000021 | 5.56 | [ACADM, AKAP6, ARID2, COL11A1, CREB1, DDX5, FOXP1, HNRNPU, HOMER1, IGF1, ITGB1, LEMD2, MEF2D, MYOCD, NLN, NR1D2, PDGFRA, PKD2, PRKAA1, RBFOX1, RPS6KB1, SKIL] |
| GO:0014706 | striated muscle tissue development | 0.000000 | 0.000022 | 5.56 | [ACADM, AKAP6, ARID2, COL11A1, CREB1, DDX5, FOXP1, HNRNPU, HOMER1, IGF1, ITGB1, LEMD2, MEF2D, MYOCD, NLN, NR1D2, PDGFRA, PRKAA1, RBFOX1, RPS6KB1, SKIL] |
| GO:0014706 | striated muscle tissue development | 0.000000 | 0.000022 | 5.56 | [ACADM, AKAP6, ARID2, COL11A1, CREB1, DDX5, FOXP1, HNRNPU, HOMER1, IGF1, ITGB1, LEMD2, MEF2D, MYOCD, NLN, NR1D2, PDGFRA, PRKAA1, RBFOX1, RPS6KB1, SKIL |
| GO:0060509 | type I pneumocyte differentiation | 0.000000 | 0.000025 | 80.00 | CREB1, NFIB, THRA, THRB |
| GO:0061061 | muscle structure development | 0.000002 | 0.000156 | 4.05 | ACADM, AKAP6, ANKRD17, BASP1, COL11A1, CREB1, DDX5, EREG, ETV1, FOXP1, HNRNPU, HOMER1, IGF1, ITGB1, LEMD2, MBNL1, MEF2D, MYOCD, NLN, NR1D2, PDGFRA, PRKAA1, RBFOX1, RPS6KB1, SKIL, THRA |
| GO:0061061 | muscle structure development | 0.000002 | 0.000156 | 4.05 | ACADM, AKAP6, ANKRD17, BASP1, COL11A1, CREB1, DDX5, EREG, ETV1, FOXP1, HNRNPU, HOMER1, IGF1, ITGB1, LEMD2, MBNL1, MEF2D, MYOCD, NLN, NR1D2, PDGFRA, PRKAA1, RBFOX1, RPS6KB1, SKIL, THRA |
| GO:0060538 | skeletal muscle organ development | 0.000002 | 0.000158 | 7.14 | BASP1, DDX5, FOXP1, HOMER1, LEMD2, MEF2D, MYOCD, NLN, NR1D2, PRKAA1, RBFOX1, RPS6KB1, SKIL |
| GO:0016202 | regulation of striated muscle tissue development | 0.000003 | 0.000170 | 8.40 | AKAP6, CREB1, DDX5, FOXP1, IGF1, MYOCD, NLN, NR1D2, PRKAA1, RBFOX1, RPS6KB1 |
| GO:1901861 | regulation of muscle tissue development | 0.000003 | 0.000169 | 8.27 | AKAP6, CREB1, DDX5, FOXP1, IGF1, MYOCD, NLN, NR1D2, PRKAA1, RBFOX1, RPS6KB1 |
| miRNA | Target Gene Symbol | Description | Biological Processes |
|---|---|---|---|
| hsa-miR-1, hsa-let-7b-5p | FOXP1 | forkhead box P1 | heart development |
| hsa-miR-1 | MYOCD | myocardin | regulation of smooth muscle contraction |
| hsa-miR-20b-5p | PKD2 | polycystic kidney disease 2 | heart development |
| hsa-miR-30b-5p | MBNL1 | muscleblind-like | striated muscle tissue development |
| hsa-miR-20b-5p | ATP1A2 | ATPase, Na+/K+ transporting, alpha 2 (+) polypeptide | regulation of smooth muscle contraction |
| hsa-miR-20b-5p, hsa-miR-26a-5p | EREG | epiregulin | regulation of muscle cell differentiation |
| hsa-miR-1226-3p | ITGB1 | integrin, beta 1 | heart development |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woo, C.C.; Liu, W.; Lin, X.Y.; Dorajoo, R.; Lee, K.W.; Richards, A.M.; Lee, C.N.; Wongsurawat, T.; Nookaew, I.; Sorokin, V. The Interaction between 30b-5p miRNA and MBNL1 mRNA is Involved in Vascular Smooth Muscle Cell Differentiation in Patients with Coronary Atherosclerosis. Int. J. Mol. Sci. 2020, 21, 11. https://doi.org/10.3390/ijms21010011
Woo CC, Liu W, Lin XY, Dorajoo R, Lee KW, Richards AM, Lee CN, Wongsurawat T, Nookaew I, Sorokin V. The Interaction between 30b-5p miRNA and MBNL1 mRNA is Involved in Vascular Smooth Muscle Cell Differentiation in Patients with Coronary Atherosclerosis. International Journal of Molecular Sciences. 2020; 21(1):11. https://doi.org/10.3390/ijms21010011
Chicago/Turabian StyleWoo, Chin Cheng, Wenting Liu, Xiao Yun Lin, Rajkumar Dorajoo, Kee Wah Lee, A Mark Richards, Chuen Neng Lee, Thidathip Wongsurawat, Intawat Nookaew, and Vitaly Sorokin. 2020. "The Interaction between 30b-5p miRNA and MBNL1 mRNA is Involved in Vascular Smooth Muscle Cell Differentiation in Patients with Coronary Atherosclerosis" International Journal of Molecular Sciences 21, no. 1: 11. https://doi.org/10.3390/ijms21010011
APA StyleWoo, C. C., Liu, W., Lin, X. Y., Dorajoo, R., Lee, K. W., Richards, A. M., Lee, C. N., Wongsurawat, T., Nookaew, I., & Sorokin, V. (2020). The Interaction between 30b-5p miRNA and MBNL1 mRNA is Involved in Vascular Smooth Muscle Cell Differentiation in Patients with Coronary Atherosclerosis. International Journal of Molecular Sciences, 21(1), 11. https://doi.org/10.3390/ijms21010011