Influence of Acrylamide Administration on the Neurochemical Characteristics of Enteric Nervous System (ENS) Neurons in the Porcine Duodenum


Abstract
:1. Introduction
2. Results
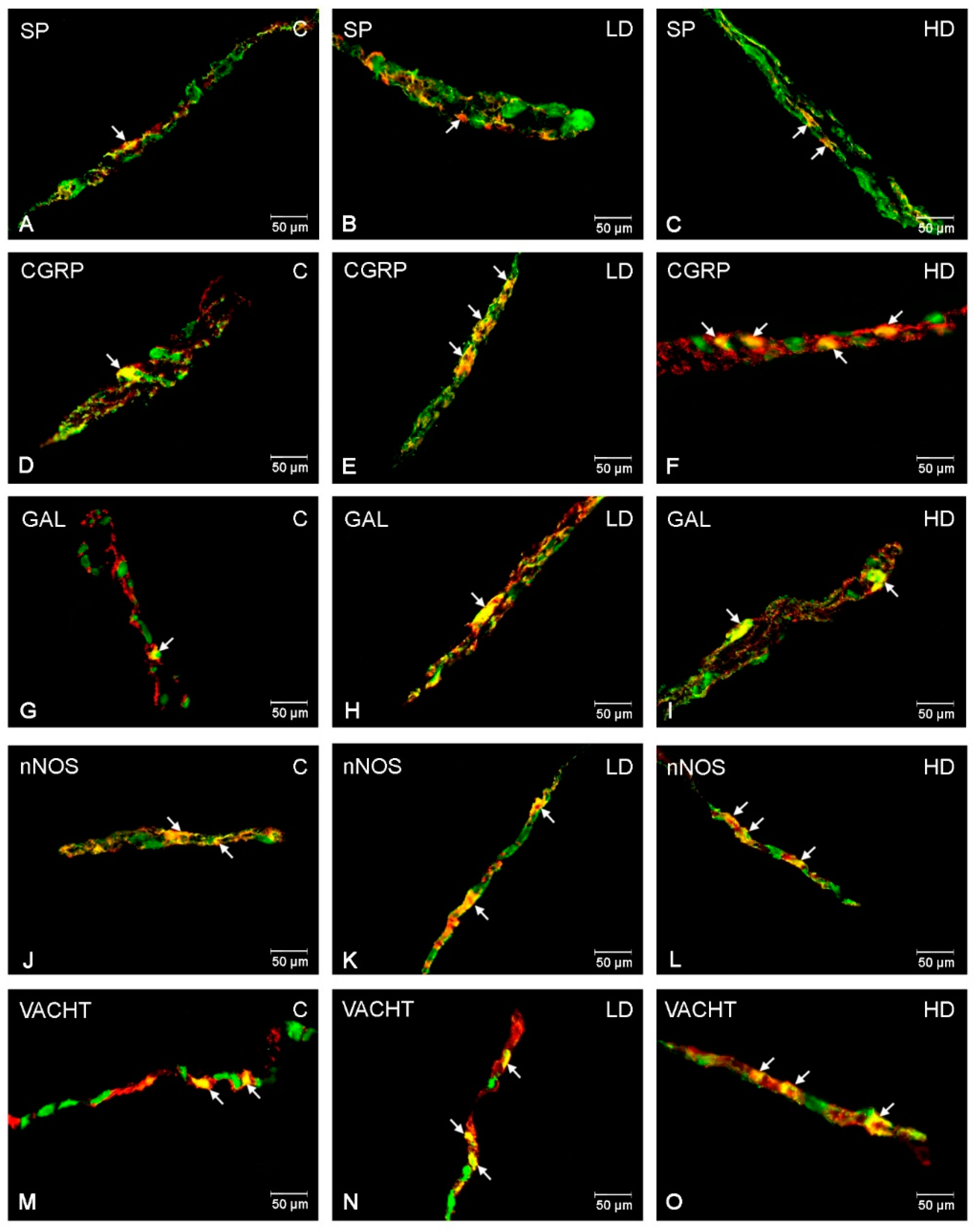
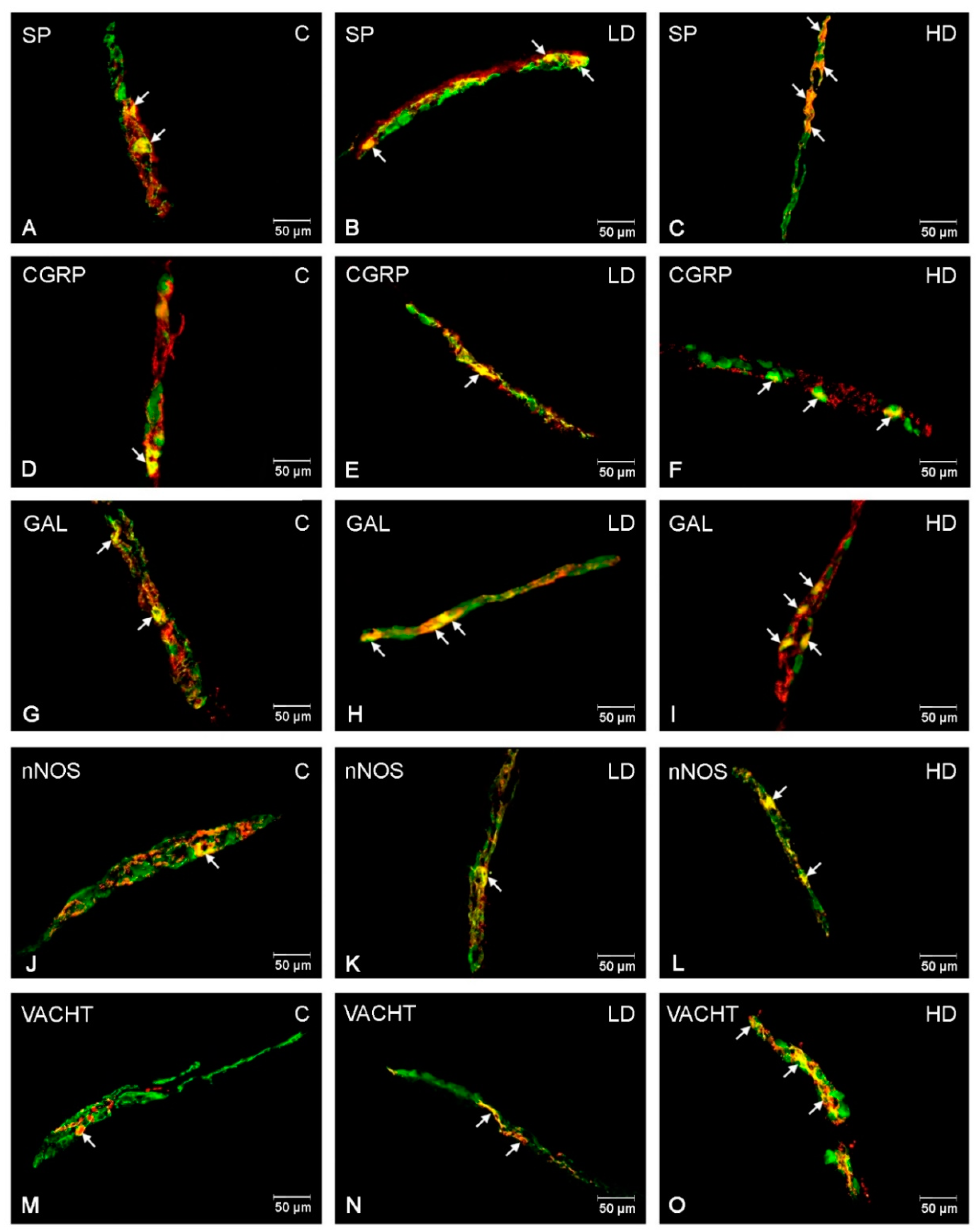
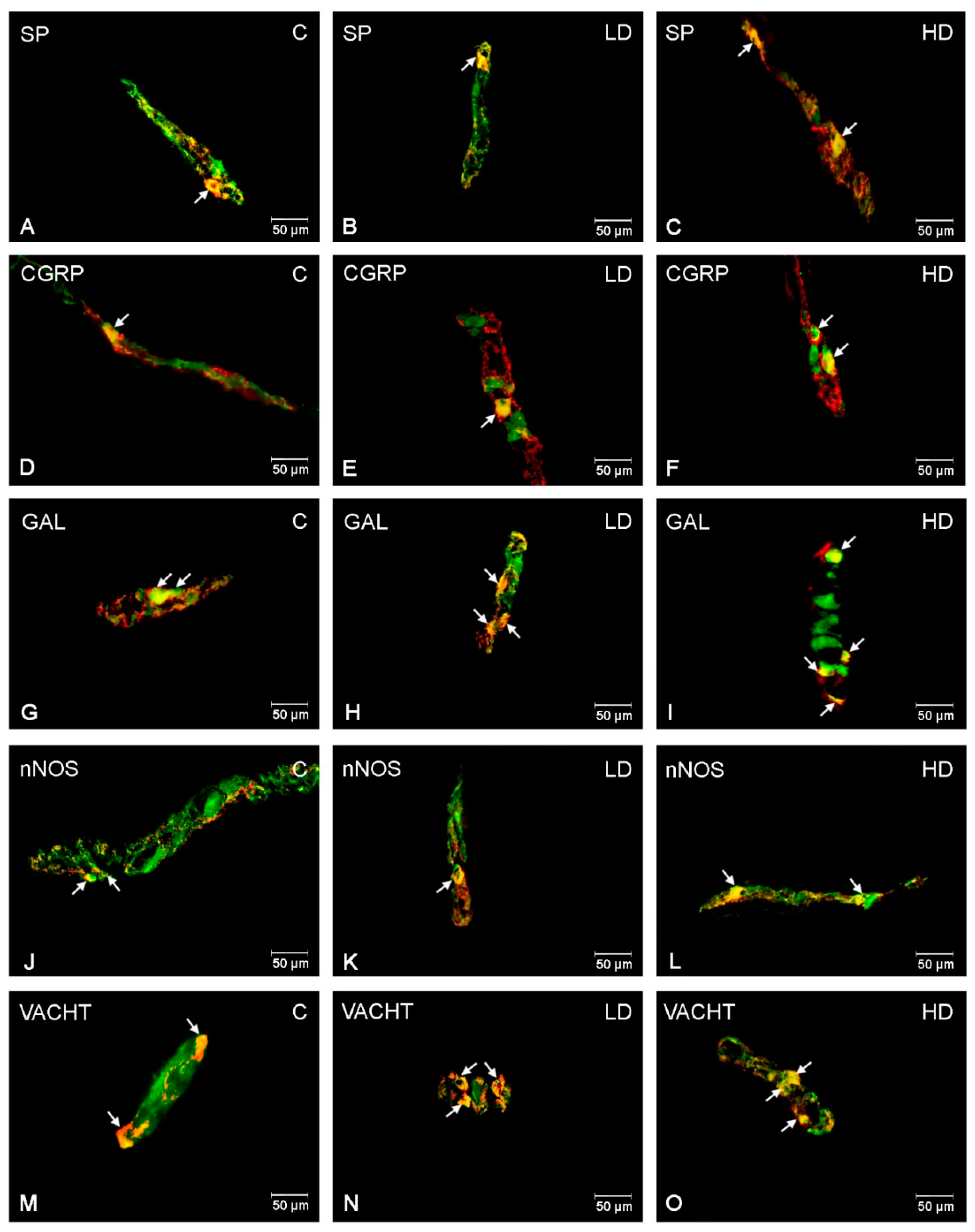
2.1. Myenteric Plexus (MP)
2.2. Outer Submucous Plexus (OSP)
2.3. Inner Submucous Plexus (ISP)
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Procedures
4.2. Double-Labelling Immunofluorescence
4.3. Counting and Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Van Lancker, F.; Adams, A.; De Kimpe, N. Chemical modifications of peptides and their impact on food properties. Chem. Rev. 2011, 111, 7876–7903. [Google Scholar] [CrossRef]
- Dybing, E.; Farmer, P.B.; Andersen, M.; Fennell, T.R.; Lalljie, S.P.; Muller, D.J.; Olin, S.; Petersen, B.J.; Schlatter, J.; Scholz, G.; et al. Human exposure and internal dose assessments of acrylamide in food. Food Chem. Toxicol. 2005, 43, 365–410. [Google Scholar] [CrossRef] [PubMed]
- Shipp, A.; Lawrence, G.; Gentry, R.; McDonald, T.; Bartow, H.; Bounds, J.; Macdonald, N.; Clewell, H.; Allen, B.; van Landingham, C. Acrylamide: Review of toxicity data and dose-response analyses for cancer and noncancer effects. Crit. Rev. Toxicol. 2006, 36, 481–608. [Google Scholar] [CrossRef] [PubMed]
- Zödl, B.; Schmid, D.; Wassler, G.; Gundacker, C.; Leibetseder, V.; Thalhammer, T.; Ekmekcioglu, C. Intestinal transport and metabolism of acrylamide. Toxicology 2007, 232, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Besaratinia, A.; Pfeifer, G.P. A review of mechanisms of acrylamide carcinogenicity. Carcinogenesis 2007, 28, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Pachin, R.M. The changing view of acrylamide neurotoxicity. Toxicol. In Vitro 2010, 25, 573–579. [Google Scholar]
- Szczerbina, T.; Banach, Z.; Tylko, G.; Pyza, E. Toxic effects of acrylamide on survival, development and haemocytes of Musca domestica. Food Chem. Toxicol. 2008, 46, 2316–2319. [Google Scholar] [CrossRef]
- Ewaleifoh, O.; Trinh, M.; Griffin, J.W.; Nguyen, T. A novel system to accelerate the progression of nerve degeneration in transgenic mouse models of neuropathies. Exp. Neurol. 2012, 237, 153–159. [Google Scholar] [CrossRef] [Green Version]
- WHO. Health Implications of Acrylamide in Food; FAO/WHO: Geneva, Switzerland, 2002; Available online: http://apps.who.int/iris/handle/10665/42563 (accessed on 15 November 2016).
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. Adv. Exp. Med. Biol. 2014, 817, 39–71. [Google Scholar]
- Palus, K.; Bulc, M.; Całka, J. Changes in somatostatin-like immunoreactivity in the sympathetic neurons projecting to the prepyloric area of the porcine stomach induced by selected pathological conditions. Biomed. Res. Int. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Khurana, R.K.; Petras, J.M. Sensory innervation of the canine esophagus, stomach, and duodenum. Am. J. Anat. 1991, 192, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Gonkowski, S. Substance P as a neuronal factor in the enteric nervous system of the porcine descending colon in physiological conditions and during selected pathogenic processes. Biofactors 2013, 39, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Nezami, B.G.; Srinivasan, S. Enteric nervous system in the small intestine: Pathophysiology and clinical implications. Curr. Gastroenterol. Rep. 2010, 12, 358–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonkowski, S.; Rowniak, M.; Wojtkiewicz, J. Zinc transporter 3 (ZnT3) in the enteric nervous system of the porcine ileum in physiological conditions and during experimental inflammation. Int. J. Mol. Sci. 2017, 18, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielefeldt, K.; Davis, B.; Binion, D.G. Pain and inflammatory bowel disease. Inflamm. Bowel Dis. 2009, 15, 778–788. [Google Scholar] [CrossRef]
- Delafoy, L.; Gelot, A.; Ardid, D.; Eschalier, A.; Bertrand, C.; Doherty, A.M.; Diop, L. Interactive involvement of brain derived neurotrophic factor, nerve growth factor, and calcitonin gene related peptide in colonic hypersensitivity in the rat. Gut 2006, 55, 940–945. [Google Scholar] [CrossRef] [Green Version]
- Szymanska, K.; Gonkowski, S. Bisphenol A-Induced changes in the enteric nervous system of the porcine duodenum. Neurotoxicology 2018, 66, 78–86. [Google Scholar] [CrossRef]
- Bulc, M.; Palus, K.; Dąbrowski, M.; Całka, J. Hyperglycaemia-induced downregulation in expression of nNOS intramural neurons of the small intestine in the pig. Int. J. Mol. Sci. 2019, 20, 1681. [Google Scholar] [CrossRef] [Green Version]
- Kasparek, M.S.; Fatima, J.; Iqbal, C.W.; Duenes, J.A.; Sarr, M.G. Role of VIP and Substance P in NANC innervation in the longitudinal smooth muscle of the rat jejunum-influence of extrinsic denervation. J. Surg. Res. 2007, 141, 22–30. [Google Scholar] [CrossRef]
- Bulc, M.; Palus, K.; Zielonka, Ł.; Gajęcka, M.; Całka, J. Changes in expression of inhibitory substances in the intramural neurons of the stomach following streptozotocin- induced diabetes in the pig. World J. Gastroenterol. 2017, 23, 6088–6099. [Google Scholar] [CrossRef]
- Costa, M.; Furness, J.B.; Llewellyn-Smith, I.J.; Cello, A.C. Projections of substance P neurons within the guinea-pig small intestine. Neuroscience 1981, 6, 411–424. [Google Scholar] [CrossRef]
- Hoyle, C.H.; Burnstock, G. Galanin-like immunoreactivity in enteric neurons of the human colon. J. Anat. 1989, 166, 23–33. [Google Scholar] [PubMed]
- Palus, K.; Całka, J. Neurochemical plasticity of the coeliac-superior mesenteric ganglion complex neurons projecting to the prepyloric area of the porcine stomach following hyperacidity. Neural Plast. 2016, 2016, 8596214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polidoro, G.; Giancola, F.; Fracassi, F.; Pietra, M.; Bettini, G.; Asti, M.; Chiocchetti, R. Substance P and the neurokinin-1 receptor expression in dog ileum with and without inflammation. Res. Vet. Sci. 2017, 114, 297–307. [Google Scholar] [CrossRef]
- Harrison, S.; Geppetti, P. Substance P. Int. J. Biochem. Cell Biol. 2001, 33, 555–576. [Google Scholar] [CrossRef]
- Evangelista, S. Role of calcitonin gene-related peptide in gastric mucosal defence and healing. Curr. Pharm. Des. 2009, 15, 3571–3576. [Google Scholar] [CrossRef]
- Wolf, M.; Schrödl, F.; Neuhuber, W.; Brehmer, A. Calcitonin gene-related peptide: A marker for putative primary afferent neurons in the pig small intestinal myenteric plexus? Anat. Rec. (Hoboken) 2007, 290, 1273–1279. [Google Scholar] [CrossRef]
- Lang, R.; Gundlach, A.L.; Kofler, B. The galanin peptide family: Receptor pharmacology, pleiotropic biological actions, and implications in health and disease. Pharm. Ther. 2007, 115, 177–207. [Google Scholar] [CrossRef]
- Shah, V.; Lyford, G.; Gores, G.; Farrugia, G. Nitric oxide in gastrointestinal health and disease. Gastroenterology 2004, 126, 903–913. [Google Scholar] [CrossRef]
- Arvidsson, U.; Riedl, M.; Elde, R.; Meister, B. Vesicular acetylcholine transporter (VAChT) protein: A novel and unique marker for cholinergic neurons in the central and peripheral nervous systems. J. Comp. Neurol. 1997, 378, 454–467. [Google Scholar] [CrossRef]
- Vasina, V.; Barbara, G.; Talamonti, L.; Stanghellini, V.; Corinaldesi, R.; Tonini, M.; De Ponti, F.; De Giorgio, R. Enteric neuroplasticity evoked by inflammation. Auton. Neurosci. 2006, 126, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Erin, N.; Türker, S.; Elpek, O.; Yıldırım, B. Differential changes in Substance P, VIP as well as neprilysin levels in patients with gastritis or ulcer. Peptides 2012, 35, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Pelucchi, C.; Bosetti, C.; Galeone, C.; La Vecchia, C. Dietary acrylamide and cancer risk: An updated meta-analysis. Int. J. Cancer 2015, 136, 2912–2922. [Google Scholar] [CrossRef] [PubMed]
- Sickles, D.W.; Stone, J.D.; Friedman, M.A. Fast axonal transport: A site of acrylamide neurotoxicity? Neurotoxicology 2002, 23, 223–251. [Google Scholar] [CrossRef]
- Erdemli, M.E.; Turkoz, Y.; Altinoz, E.; Elibol, E.; Dogan, Z. Investigation of the effects of acrylamide applied during pregnancy on fetal brain development in rats and protective role of the vitamin E. Hum. Exp. Toxicol. 2016, 35, 1337–1344. [Google Scholar] [CrossRef]
- LoPachin, R.M.; Gavin, T. Toxic neuropathies: Mechanistic insights based on a chemical perspective. Neurosci. Lett. 2015, 596, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Martyniuk, C.J.; Feswick, A.; Fang, B.; Koomen, J.M.; Barber, D.S.; Gavin, T.; LoPachin, R.M. Protein targets of acrylamide adduct formation in cultured rat dopaminergic cells. Toxicol. Lett. 2013, 219, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Park, H.R.; Lee, J.Y.; Cho, J.H.; Song, H.M.; Kim, A.H.; Lee, W.; Lee, Y.; Chang, S.C.; Kim, H.S.; et al. Learning, memory deficits, and impaired neuronal maturation attributed to acrylamide. J. Toxicol. Environ. Health A 2018, 81, 254–265. [Google Scholar] [CrossRef]
- Kwolek-Mirek, M.; Zadrag-Tecza, R.; Bednarska, S.; Bartosz, G. Yeast Saccharomyces cerevisiae devoid of Cu,Zn-superoxide dismutase as a cellular model to study acrylamide toxicity. Toxicol. In Vitro 2011, 25, 573–579. [Google Scholar] [CrossRef]
- Cao, J.; Liu, Y.; Jia, L.; Jiang, L.I.P.; Geng, C.-Y.; Yao, X.-F.; Kong, Y.; Jiang, B.-N.; Zhong, L.-F. Curcumin attenuates acrylamide-induced cytotoxicity and genotoxicity in HepG2 cells by ROS scavenging. J. Agric. Food Chem. 2008, 56, 12059–12063. [Google Scholar] [CrossRef]
- Catalgol, B.; Ozhan, G.; Alpertunga, B. Acrylamide-induced oxidative stress in human erythrocytes. Hum. Exp. Toxicol. 2009, 28, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Krishna, G.; Muralidhara. Oral supplements of combined fructo- and xylo-oligosaccharides during perinatal period significantly offsets acrylamide-induced oxidative impairments and neurotoxicity in rats. J. Physiol. Pharmacol. 2018, 69. [Google Scholar] [CrossRef]
- Zong, C.; Hasegawa, R.; Urushitani, M.; Zhang, L.; Nagashima, D.; Sakurai, T.; Ichihara, S.; Ohsako, S.; Ichihara, G. Role of microglial activation and neuroinflammation in neurotoxicity of acrylamide in vivo and in vitro. Arch. Toxicol. 2019, 93, 2007–2019. [Google Scholar] [CrossRef]
- Dortaj, H.; Yadegari, M.; Hosseini Sharif Abad, M.; Abbasi Sarcheshmeh, A.; Anvari, M. Stereological Method for Assessing the Effect of Vitamin C Administration on the Reduction of Acrylamide-induced Neurotoxicity. Basic Clin. Neurosci. 2018, 9, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Imam, R.A.; Gadallah, H.N. Acrylamide-induced adverse cerebellar changes in rats: Possible oligodendrogenic effect of omega 3 and green tea. Folia Morphol. (Warsz) 2019, 78, 564–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdemli, Z.; Erdemli, M.E.; Turkoz, Y.; Gul, M.; Yigitcan, B.; Gozukara Bag, H. The effects of acrylamide and Vitamin E administration during pregnancy on adult rats testis. Andrologia 2019, 51, e13292. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, E.; Dobrowolski, P.; Puzio, I.; Prost, L.; Kurlak, P.; Sawczuk, P.; Badzian, B.; Hulas-Stasiak, M.; Kostro, K. Acrylamide-induced prenatal programming of intestine structure in guinea pig. J. Physiol. Pharmacol. 2014, 65, 107–115. [Google Scholar] [PubMed]
- Sandgren, K.; Lin, Z.; Fex Svenningsen, A.; Ekblad, E. Vasoactive intestinal peptide and nitric oxide promote survival of adult rat myenteric neurons in culture. J. Neurosci. Res. 2003, 72, 595–602. [Google Scholar] [CrossRef]
- McQuade, R.M.; Stojanovska, V.; Donald, E.L.; Rahman, A.A.; Campelj, D.G.; Abalo, R.; Rybalka, E.; Bornstein, J.C.; Nurgali, K. Irinotecan-Induced Gastrointestinal Dysfunction Is Associated with Enteric Neuropathy, but Increased Numbers of Cholinergic Myenteric Neurons. Front. Physiol. 2017, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Thompson, S.A.; Smith, O.; Linn, D.M.; Linn, C.L. Acetylcholine neuroprotection against glutamate-induced excitotoxicity in adult pig retinal ganglion cells is partially mediated through alpha4 nAChRs. Exp. Eye Res. 2006, 83, 1135–1145. [Google Scholar] [CrossRef]
- Santhanasabapathy, R.; Vasudevan, S.; Anupriya, K.; Pabitha, R.; Sudhandiran, G. Farnesol quells oxidative stress, reactive gliosis and inflammation during acrylamid-induced neurotoxicity: Behavioral and biochemical evidence. Neuroscience 2015, 308, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Naruszewicz, M.; Zapolska-Downar, D.; Kósmider, A.; Nowicka, G.; Kozłowska-Wojciechowska, M.; Vikström, A.S.; Törnqvist, M. Chronic intake of potato chips in humans increases the production of reactive oxygen radicals by leukocytes and increases plasma C-reactive protein: A pilot study. Am. J. Clin. Nutr. 2009, 89, 773–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palus, K.; Obremski, K.; Bulc, M.; Całka, J. The impact of low and high doses of acrylamide on the intramural neurons of the porcine ileum. Food Chem. Toxicol. 2019, 132, 110673. [Google Scholar] [CrossRef]
- Mantyh, C.R.; Vigna, S.R.; Maggio, J.E.; Mantyh, P.W.; Bollinger, R.R.; Pappas, T.N. Substance P binding sites on intestinal lymphoid aggregates and blood vessels in inflammatory bowel disease correspond to authentic NK-1 receptors. Neurosci. Lett. 1994, 178, 255–259. [Google Scholar] [CrossRef]
- Islek, A.; Yilmaz, A.; Elpek, G.O.; Erin, N. Childhood chronic gastritis and duodenitis: Role of altered sensory neuromediators. World J. Gastroenterol. 2016, 22, 8349–8360. [Google Scholar] [CrossRef] [PubMed]
- Brancati, S.B.; Zádori, Z.S.; Németh, J.; Gyires, K. Substance P induces gastric mucosal protection at supraspinal level via increasing the level of endomorphin-2 in rats. Brain Res Bull. 2013, 91, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mönnikes, H.; van der Voort, I.R.; Wollenberg, B.; Heymann-Monnikes, I.; Tebbe, J.J.; Alt, W.; Arnold, R.; Klapp, B.F.; Wiedenmann, B.; McGregor, G.P. Gastric perception thresholds are low and sensory neuropeptide levels high in helicobacter pyloripositive functional dyspepsia. Digestion 2005, 71, 111–123. [Google Scholar] [CrossRef]
- Azzolina, A.; Bongiovanni, A.; Lampiasi, N. Substance P induces TNF-alpha and IL- 6 production through NF kappa B in peritoneal mast cells. Biochim. Biophys. Acta 2003, 7, 1–3. [Google Scholar]
- Li, W.; Hou, L.; Hua, Z.; Wang, X. Interleukin-1β induces β-calcitonin gene-related peptide secretion in human type II alveolar epithelial cells. FASEB J. 2004, 18, 1603–1605. [Google Scholar] [CrossRef] [Green Version]
- Ohno, T.; Hattori, Y.; Komine, R.; Ae, T.; Mizuguchi, S.; Arai, K.; Saeki, T.; Suzuki, T.; Hosono, K.; Hayashi, I.; et al. Roles of calcitonin gene-related peptide in maintenance of gastric mucosal integrity and in enhancement of ulcer healing and angiogenesis. Gastroenterology 2008, 134, 215–225. [Google Scholar] [CrossRef]
- Makowska, K.; Gonkowski, S. The influence of inflammation and nerve damage on the neurochemical characterization of calcitonin gene-related peptide-like immunoreactive (CGRP-LI) neurons in the enteric nervous system of the porcine descending colon. Int. J. Mol. Sci. 2018, 19, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.J.; Zou, Y.Y.; Cui, Y.; Yin, Y.; Guo, G.; Lu, F.G. Calcitonin gene-related peptide is a promising marker in ulcerative colitis. Dig. Dis. Sci. 2013, 58, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Matkowskyj, K.A.; Nathaniel, R.; Prasad, R.; Weihrauch, D.; Rao, M.; Benya, R.V. Galanin contributes to the excess colonic fluid secretion observed in dextran sulfate sodium murine colitis. Inflamm. Bowel Dis. 2004, 10, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Zalecki, M.; Pidsudko, Z.; Franke-Radowiecka, A.; Wojtkiewicz, J.; Kaleczyc, J. Galaninergic intramural nerve and tissue reaction to antral ulcerations. Neurogastroenterol. Motil. 2018, 30, e13360. [Google Scholar] [CrossRef] [PubMed]
- Matkowskyj, K.; Royan, S.V.; Blunier, A.; Hecht, G.; Rao, M.; Benya, R.V. Age-dependent differences in galanin-dependent colonic fluid secretion after infection with Salmonella typhimurium. Gut 2009, 58, 1201–1206. [Google Scholar] [CrossRef]
- Dallos, A.; Kiss, M.; Polyánka, H.; Dobozy, A.; Kemény, L.; Husz, S. Galanin receptor expression in cultured human keratinocytes and in normal human skin. J. Peripher. Nerv. Syst. 2006, 11, 156–164. [Google Scholar] [CrossRef]
- Leite, H.R.; Oliveira-Lima, O.C.; Pereira, L.M.; Oliveira, V.E.M.; Prado, V.F.; Prado, M.A.M.; Pereira, G.S.; Massensini, A.R. Vesicular acetylcholine transporter knock down-mice are more susceptible to inflammation, c-Fos expression and sickness behavior induced by lipopolysaccharide. Brain Behav. Immun. 2016, 57, 282–292. [Google Scholar] [CrossRef]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef]
- Winston, J.H.; Li, Q.; Sarna, S.K. Paradoxical regulation of ChAT and nNOS expression in animal models of Crohn’s colitis and ulcerative colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G295–G302. [Google Scholar] [CrossRef] [Green Version]
- Rychlik, A.; Gonkowski, S.; Nowicki, M.; Calka, J. Inflammatory bowel disease affects density of nitrergic nerve fibers in the mucosal layer of the canine gastrointestinal tract. Can. J. Vet. Res. 2017, 81, 129–136. [Google Scholar]
- Rivera, L.R.; Poole, D.P.; Thacker, M.; Furness, J.B. The involvement of nitric oxide synthase neurons in enteric neuropathies. Neurogastroenterol. Motil. 2011, 23, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T. Pathophysiological significance of neuronal nitric oxide synthase in the gastrointestinal tract. J. Gastrenterol. 2003, 38, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Kararli, T.T. Comparison of the gastrointestinal anatomy, physiology, and biochemistry of humans and commonly used laboratory animals. Biopharm. Drug Dispos. 1995, 16, 351–380. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Timmermans, J.P. Lessons from the porcine enteric nervous system. Neurogastroenterol. Motil. 2004, 16, 50–54. [Google Scholar] [CrossRef]
- Kobayashi, E.; Hanazono, Y.; Kunita, S. Swine used in the medical university: Overview of 20 years of experience. Exp. Anim. 2018, 67, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Palus, K.; Makowska, K.; Całka, J. Acrylamide-induced alterations in the cocaine- and amphetamine-regulated peptide transcript (CART)-like immunoreactivity within the enteric nervous system of the porcine small intestines. Ann. Anat. 2018, 219, 94–101. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| MP | OSP | ISP | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Experimental Group | C Group | LD Group | HD Group | C Group | LD Group | HD Group | C Group | LD Group | HD Group |
| SP | 0.69 ± 0.14 | 1.05 ± 0.25 (**) | 2.67 ± 0.44 (**) | 21.47 ± 1.19 | 25.83 ± 1.28 (***) | 38.50 ± 1.23 (***) | 11.57± 1.17 | 23.51± 1.18 (**) | 26.77± 1.75 (***) |
| CGRP | 12.38 ± 1.02 | 21.75 ± 0.90 (***) | 31.54 ±0.70 (***) | 14.62 ± 1.20 | 16.15 ± 0.68 | 27.44 ± 0.99 (***) | 20.14 ± 0.69 | 22.59 ± 0.65 (*) | 28.64 ± 0.77 (***) |
| GAL | 2.87 ± 0.41 | 6.45 ± 0.70 (***) | 10.24 ± 0.32 (***) | 31.75 ± 1.41 | 39.10 ± 0.81 (***) | 49.78 ± 0.64 (***) | 38.86 ± 1.53 | 49.12 ± 1.15 (***) | 54.45± 1.35 (***) |
| nNOS | 29.34 ± 1.78 | 31.55 ± 1.12 | 37.39 ± 0.98 (**) | 2.41 ± 0.60 | 4.62 ± 0.28 (*) | 9.20 ± 0.46 (***) | 1.53 ± 0.19 | 2.84 ± 0.26 | 7.46 ± 0.65 (***) |
| VACHT | 13.92 ± 0.91 | 20.22 ± 0.46 (**) | 24.89 ± 1.50 (***) | 20.80 ± 1.00 | 26.88 ± 0.69 (**) | 33.60 ± 1.48 (***) | 42.74 ± 1.75 | 52.81 ± 1.43 (**) | 61.96 ± 1.06 (***) |
| Antigen | Host Species | Cat No. | Dilution | Supplier |
|---|---|---|---|---|
| Primary Antibodies | ||||
| PGP 9.5 | Mouse | 7863-2004 | 1:1000 | Bio-Rad, Hercules, CA, USA |
| SP | Rat | 8450-0505 | 1:150 | AbD Serotec, Raleigh, NC, USA |
| CGRP | Rabbit | MAB317 | 1:4000 | Millipore, Burlington, MA, USA |
| GAL | Rabbit | RIN7153 | 1:3000 | Peninsula, San Carlos, CA, USA, |
| nNOS | Rabbit | AB5380 | 1:2000 | Sigma-Aldrich, Saint Louis, MO, USA |
| VACHT | Rabbit | H-V007 | 1:2000 | Phoenix Pharmaceuticals, Burlingame, CA, USA |
| Secondary Antibodies | ||||
| Alexa Fluor 488 donkey anti-mouse IgG | A21202 | 1:1000 | Thermo Fisher Scientific, Waltham, MA, USA | |
| Alexa Fluor 546 goat anti-rabbit IgG | A11010 | 1:1000 | Thermo Fisher Scientific, Waltham, MA, USA | |
| Alexa Fluor 546 goat anti-rat IgG | A11081 | 1:1000 | Thermo Fisher Scientific, Waltham, MA, USA | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palus, K.; Całka, J. Influence of Acrylamide Administration on the Neurochemical Characteristics of Enteric Nervous System (ENS) Neurons in the Porcine Duodenum. Int. J. Mol. Sci. 2020, 21, 15. https://doi.org/10.3390/ijms21010015
Palus K, Całka J. Influence of Acrylamide Administration on the Neurochemical Characteristics of Enteric Nervous System (ENS) Neurons in the Porcine Duodenum. International Journal of Molecular Sciences. 2020; 21(1):15. https://doi.org/10.3390/ijms21010015
Chicago/Turabian StylePalus, Katarzyna, and Jarosław Całka. 2020. "Influence of Acrylamide Administration on the Neurochemical Characteristics of Enteric Nervous System (ENS) Neurons in the Porcine Duodenum" International Journal of Molecular Sciences 21, no. 1: 15. https://doi.org/10.3390/ijms21010015
APA StylePalus, K., & Całka, J. (2020). Influence of Acrylamide Administration on the Neurochemical Characteristics of Enteric Nervous System (ENS) Neurons in the Porcine Duodenum. International Journal of Molecular Sciences, 21(1), 15. https://doi.org/10.3390/ijms21010015