Crossbridge Recruitment Capacity of Wild-Type and Hypertrophic Cardiomyopathy-Related Mutant Troponin-T Evaluated by X-ray Diffraction and Mechanical Study of Cardiac Skinned Fibers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
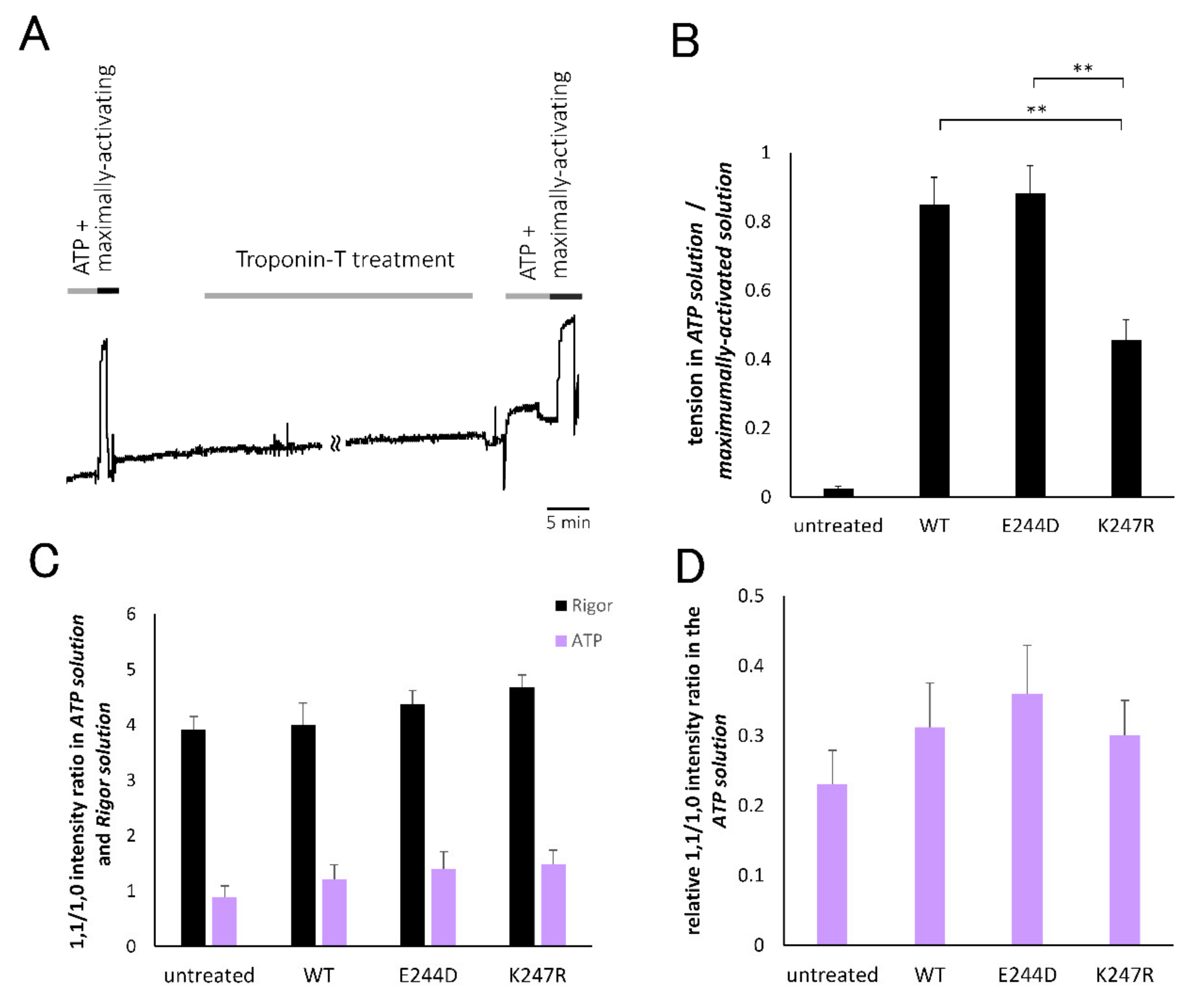
2. Results
2.1. Intensity Change of Troponin Reflection of Cardiac Muscle Fibers with No Treatment
2.2. Intensity Change of Troponin Reflection of Cardiac Muscle Fibers after Treatment with Wild-Type TnT
2.3. Intensity Change of Troponin Reflection of Cardiac Muscle Fibers after Treatment with The Mutant TnT
2.4. Evaluation of Actomyosin Interaction of Cardiac Muscle Fibers with or without TnT-Treatment Based on Tension Development and Equatorial Reflection Measurement
3. Discussion
3.1. Intensity Drop of Troponin Reflection Triggered by Contractile Crossbridge Formation
3.2. Implication for the Pathogenesis of the K247R Mutant Cardiac Fibers
3.3. Pathogenesis of the E244E Mutant Cardiac Fibers
4. Materials and Methods
4.1. Preparation of Skinned Cardiac Fiber from Rat Ventricle
4.2. Replacement of Endogenous Troponin T/I/C Complex with Exogenous TnT
4.3. X-ray Diffraction Experiments
4.4. Measurement of Tension Development in Cardiac Muscle Skinned Fibers
4.5. Analysis of Diffraction Patterns
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TnT | Troponin T |
| TnI | Troponin I |
| TnC | Troponin C |
| HCM | Hypertrophic cardiomyopathy |
| S.E.M. | Standard error of the mean |
References
- Perz-Edwardsa, R.J.; Irving, T.C.; Baumann, B.A.J.; Gore, D.; Hutchinson, D.C.; Uroš Kržičd Porter, R.L.; Ward, A.B.; Reedy, M.K. X-ray diffraction evidence for myosin-troponin connections and tropomyosin movement during stretch activation of insect flight muscle. Proc. Natl. Acad. Sci. USA 2011, 108, 120–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, T.; Iwamoto, H.; Yagi, N. Monitoring the structural behavior of troponin and myoplasmic free Ca2+ concentration during twitch of frog skeletal muscle. Biophys. J. 2010, 99, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, T.; Ueno, Y.; Takezawa, Y.; Sugimoto, T.; Oda, T.; Wakabayashi, K. X-ray fiber diffraction modeling of the thin filament upon activation of live vertebrate skeletal muscle. Biophysics 2010, 6, 13–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, T.; Wakayama, J.; Inoue, K.; Yagi, N.; Iwamoto, H. Dynamics of thin-filament activation in rabbit skeletal muscle fibers examined by time-resolved X-ray diffraction. Biophys. J. 2009, 96, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, T.; Yagi, N. Structural changes in the muscle thin filament during contractions caused by single and double electrical pulses. J. Mol. Biol. 2008, 383, 1019–1036. [Google Scholar] [CrossRef]
- Yagi, N. An X-ray diffraction study on early structural changes in skeletal muscle contraction. Biophys. J. 2003, 84, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Koubassova, N.A.; Bershitsky, S.Y.; Ferenczi, M.A.; Narayanan, T.; Tsaturyan, A.K. Tropomyosin movement is descriibed by a quantitative high-resolution model of X-ray diffraction of contracting muscle. Eur. Biophys. J. 2017, 46, 335–342. [Google Scholar] [CrossRef]
- Behrman, E.; Muller, M.; Penczek, P.A.; Mannherz, H.G.; Manstein, D.J.; Raunser, S. Structure of the rigor actin-tropomyosin-myosin complex. Cell 2012, 150, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Maytum, R.; Lehrer, S.S.; Geeves, M.A. Cooperativity and switching within the three-state model of muscle regulation. Biochemistry 1999, 38, 1102–1110. [Google Scholar] [CrossRef]
- Hatakenaka, M.; Ohtsuki, I. Effect of removal and reconstitution of troponins C and I on the Ca2+-activated tension development of single glycerinated rabbit skeletal muscle fibers. Eur. J. Biochem. 1992, 205, 985–993. [Google Scholar] [CrossRef]
- Lu, Q.W.; Wu, X.Y.; Morimoto, S. Inherited cardiomyopathies caused by troponin mutations. J. Geriatr. Cardiol. 2013, 10, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Takemori, S. Activating efficiency of Ca2+ and cross-bridges as measured by phosphate analog release. Biophys. J. 2001, 80, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Güth, K.; Potter, J.D. Effect of rigor and cycling cross-bridges on the structure of troponin C and on the Ca2+ affinity of the Ca2+-specific regulatory sites in skinned rabbit psoas fibers. J. Biol. Chem. 1987, 262, 13627–13635. [Google Scholar]
- Watkins, H.; McKenna, W.J.; Thierfelder, L.; Suk, H.J.; Anan, R.; O’Donoghue, A.; Spirito, P.; Matsumori, A.; Moravec, C.S.; Seidman, J.G.; et al. Mutations in the genes for cardiac troponin T and alpha-tropomyosin in hypertrophic cardiomyopathy. N. Engl. J. Med. 1995, 332, 1058–1064. [Google Scholar] [CrossRef]
- Garcia-Castro, M.; Rreguero, J.R.; Batalla, B.A.; Diaz-molina, B.; Gonzalez, P.; Alvarez, V.; Cortina, A.; Cubero, G.I.; Coto, E. Hypertrophic Cardiomyopathy: Low Frequency of mutations in the β-myosin heavy chain (MYH7) and cardiac TnT (TNNT2) genes among Spanish patients. Clin. Chem. Mol. Diagn. Genet. 2003, 49, 1279–1285. [Google Scholar] [CrossRef]
- Møller DVAndersen PSHedley, P.; Ersboll, M.K.; Bundgaard, H.; Smook JMChristiansen, M.; Kober, L. The role of sarcomere gene mutations in patients with idiopathic dilated cardiomyopathy. Eur. J. Human Genet. 2009, 17, 1241–1249. [Google Scholar] [CrossRef] [Green Version]
- Nakaura, H.; Yanaga, F.; Ohtsuki, I.; Morinoto, S. Effects of missense mutations Phe110Ile and Glu244Asp in human cardiac troponin T on force generation in skinned cardac muscle fibers. J. Biochem. 1999, 126, 457–460. [Google Scholar] [CrossRef]
- Harada, K.; Potters, J.D. Familial hypertrophic myopathy mutations from different functional regions of troponinT result in different effects on the pH and Ca2+ sensitivity of cardiac muscle contraction. J. Biol. Chem. 2004, 279, 14488–14495. [Google Scholar] [CrossRef] [Green Version]
- Martin-Fernandez, M.L.; Bordas, J.; Diakun, G.; Harries, J.; Lowy, J.; Mant, G.R.; Svensson, A.; Towns-Andrews, E. Time-resolved x-ray diffraction studies of myosin head movements in live frog sartorius muscle during isometric and isotonic contractions. J. Muscle Res. Cell Motil. 1994, 15, 319–348. [Google Scholar] [CrossRef]
- Matsumoto, F.; Maeda, K.; Chatake, T.; Maeda, Y. Functional aberration of myofibrils by cardiomyopathy-causing mutations in the coiled-coil region of the troponin-core domain. Biochem. Biophys. Res. Commun. 2009, 382, 205–209. [Google Scholar] [CrossRef]
- Ford, S.V.; Mallampalli, S.; Chandra, M. Cardiomyopathy-related mutations (E244D, K247R, D270N, and K273E) in the H2-Helix of cardiac troponin T have varied effectsi on myofilament responsiveness to calcium and crossbridge recruitment dynamics. Biophys. J. 2012, 102, 357a. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Kimura, M.; Takemori, S.; Ohno, T.; Ohtsuka, Y.; Akiyama, N.; Yagi, N. The molecular mechanism of cardiomyopathy caused by mutant troponin; a molecular dynamics study tested by and X-ray diffraction experiment. J. Physiol. Sci. 2009, 59 (Suppl. 1), 60. [Google Scholar]
- Matsuo, T.; Tominaga, T.; Kono, F.; Shibata, K.; Fujiwara, S. Modulation of the picosecond dynamics of troponin by the cardiomyopathy-causing mutation K247R of troponin T observed by quasielastic neutron scattering. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Namba, K.; Fujii, T. Cardiac muscle thin filament structures reveal calcium regulatory mechanism. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Kono, F.; Fujiwara, S. Effects of cardiomyopathy-causing E244D mutation of tropninT on the structures of cardiac thin filaments studied by small-angle X-ray scattering. J. Struct. Biol. 2019, 205, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Yanaga, F.; Morimoto, S.; Ohtsuki, I. Ca2+ sensitization and potentiation of the maximum level of myofibrillar ATPase activity caused by mutations of troponin T found in familial hypertrophic cardiomyopathy. J. Biol. Chem. 1999, 274, 8806–8812. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Kimura, M.; Li, Z.B.; Ohno, T.; Takemori, S.; Hoh, J.F.Y.; Yagi, N. X-ray diffraction analysis of the effects of myosin regulatory light chain phosphorylation and butanedione monoxime on skinned skeletal muscle fibers. Am. J. Physiol. 2016, 310, C692–C700. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamaguchi, M.; Kimura, M.; Ohno, T.; Nakahara, N.; Akiyama, N.; Takemori, S.; Yagi, N. Crossbridge Recruitment Capacity of Wild-Type and Hypertrophic Cardiomyopathy-Related Mutant Troponin-T Evaluated by X-ray Diffraction and Mechanical Study of Cardiac Skinned Fibers. Int. J. Mol. Sci. 2020, 21, 3520. https://doi.org/10.3390/ijms21103520
Yamaguchi M, Kimura M, Ohno T, Nakahara N, Akiyama N, Takemori S, Yagi N. Crossbridge Recruitment Capacity of Wild-Type and Hypertrophic Cardiomyopathy-Related Mutant Troponin-T Evaluated by X-ray Diffraction and Mechanical Study of Cardiac Skinned Fibers. International Journal of Molecular Sciences. 2020; 21(10):3520. https://doi.org/10.3390/ijms21103520
Chicago/Turabian StyleYamaguchi, Maki, Masako Kimura, Tetsuo Ohno, Naoya Nakahara, Nobutake Akiyama, Shigeru Takemori, and Naoto Yagi. 2020. "Crossbridge Recruitment Capacity of Wild-Type and Hypertrophic Cardiomyopathy-Related Mutant Troponin-T Evaluated by X-ray Diffraction and Mechanical Study of Cardiac Skinned Fibers" International Journal of Molecular Sciences 21, no. 10: 3520. https://doi.org/10.3390/ijms21103520
APA StyleYamaguchi, M., Kimura, M., Ohno, T., Nakahara, N., Akiyama, N., Takemori, S., & Yagi, N. (2020). Crossbridge Recruitment Capacity of Wild-Type and Hypertrophic Cardiomyopathy-Related Mutant Troponin-T Evaluated by X-ray Diffraction and Mechanical Study of Cardiac Skinned Fibers. International Journal of Molecular Sciences, 21(10), 3520. https://doi.org/10.3390/ijms21103520