Becoming Stressed: Does the Age Matter? Reviewing the Neurobiological and Socio-Affective Effects of Stress throughout the Lifespan


 ,
,  and
and 
Abstract
:1. Introduction
2. Stress Exposure during the Prenatal Period: Prenatal Stress
3. Stress Exposure during the Perinatal Period: Perinatal Stress
4. Stress Exposure during Late Childhood and Adolescence: Peripubertal Stress
5. Stress Exposure during Adulthood: Adult Stress
6. Stress Exposure during Old Age: Elderly Stress
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| ACTH | Adrenocorticotropic hormone |
| AEA | Anandamide |
| ASD | Autism spectrum disorders |
| Bdnf | Brain derived neurotrophic factor |
| BMA | Basomedial nucleus of the amygdala |
| BLA | Basolateral nucleus of the amygdala |
| CB1-R | Cannabinoid type 1 receptor |
| CeA | Central nucleus of the amygdala |
| CeM | Centromedial nucleus of the amygdala |
| CORT | Corticosterone |
| CRS | Chronic restraint stress |
| CRF | Corticotrophin releasing factor |
| CSDS | Chronic social defeat stress |
| CUS | Chronic unpredictable stress |
| Dnmt | DNA methyltransferase |
| eCB | Endocannabinoid |
| FAAH | Fatty acid amine hydrolase |
| GAD67 | Glutamic acid decarboxylase enzime, 67KDa |
| GR | Gucocorticoid receptor |
| HPA | Hypothalamic pituitary adrenal |
| LA | Lateral nucleus of the amygdala |
| MeA | Medial nucleus of the amygdala |
| NR-1 | Subunit 1 of the N-Metyl-D-Aspartate (NMDA) receptor |
| OXTR | Oxytocin receptor |
| POMC | Proopiomelano-cortin |
| PSA-NCAM | Polysialylated form of the neural cell adhesion molecule (NCAM) |
| PTSD | Post-traumatic stress disorder |
| PV | Parvalbumin |
| PWSI | Post-weaning social isolation |
| RORA | RAR related orphan receptor A (circadian rhythm-related gene) |
| UCMS | Unpredictable chronic mild stress |
| VGAT | Vesicular GABA transporter |
| VGLUT1 | Vesicular glutamate transporter type 1 |
| WHO | World Health Organization |
| 2-AG | 2-arachidonoyl-glycerol |
| Δ9-THC | Δ9-Tetrahydrocannabinol |
References
- Young, S.N. The neurobiology of human social behaviour: An important but neglected topic. J. Psychiatry Neurosci. 2008, 33, 391–392. [Google Scholar] [PubMed]
- Kennedy, D.P.; Adolphs, R. The social brain in psychiatric and neurological disorders. Trends Cogn. Sci. 2012, 16, 559–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.S.; Beery, A.K. Neural Circuits Underlying Rodent Sociality: A Comparative Approach. In Current Topics in Behavioral Neurosciences; Springer: Berlin/Heidelberg, Germany, 2019; Volume 43, pp. 211–238. [Google Scholar]
- Insel, T.R.; Fernald, R.D. How the brain processes social information: Searching for the social brain. Annu. Rev. Neurosci. 2004, 27, 697–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzanoulinou, S.; Sandi, C. The Programming of the Social Brain by Stress During Childhood and Adolescence: From Rodents to Humans. In Social Behavior from Rodents to Humans; Springer: Cham, Switzerland, 2015; pp. 411–429. [Google Scholar]
- Hensch, T.K.; Bilimoria, P.M. Re-opening Windows: Manipulating Critical Periods for Brain Development. Cerebrum 2012, 2012, 11. [Google Scholar]
- Marco, E.M.; MacRì, S.; Laviola, G. Critical age windows for neurodevelopmental psychiatric disorders: Evidence from animal models. Neurotox. Res. 2011, 19, 286–307. [Google Scholar] [CrossRef]
- Tóth, M.; Halász, J.; Mikics, É.; Barsy, B.; Haller, J. Early Social Deprivation Induces Disturbed Social Communication and Violent Aggression in Adulthood. Behav. Neurosci. 2008, 122, 849–854. [Google Scholar] [CrossRef]
- Toth, M.; Mikics, E.; Tulogdi, A.; Aliczki, M.; Haller, J. Post-weaning social isolation induces abnormal forms of aggression in conjunction with increased glucocorticoid and autonomic stress responses. Horm. Behav. 2011, 60, 28–36. [Google Scholar] [CrossRef]
- Sandi, C.; Haller, J. Stress and the Social Brain: Behavioural Effects and Neurobiological Mechanisms. Nat. Rev. Neurosci. 2015, 16. [Google Scholar] [CrossRef] [Green Version]
- Paus, T.; Keshavan, M.; Giedd, J.N. Why Do Many Psychiatric Disorders Emerge During Adolescence? Nat. Rev. Neurosci. 2008, 9. [Google Scholar] [CrossRef] [Green Version]
- Bale, T.L.; Epperson, C.N. Sex differences and stress across the lifespan. Nat. Neurosci. 2015, 18, 1413–1420. [Google Scholar] [CrossRef]
- Novais, A.; Monteiro, S.; Roque, S.; Correia-Neves, M.; Sousa, N. How age, sex and genotype shape the stress response. Neurobiol. Stress 2017, 6, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Sousa, N.; Almeida, O.F.X. Disconnection and reconnection: The morphological basis of (mal)adaptation to stress. Trends Neurosci. 2012, 35, 742–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beery, A.K.; Kaufer, D. Stress, social behavior, and resilience: Insights from rodents. Neurobiol. Stress 2015, 1, 116–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S. Neurobiological and Systemic Effects of Chronic Stress. Chronic Stress 2017, 1, 2470547017692328. [Google Scholar] [CrossRef] [Green Version]
- Yehuda, R.; Schmeidler, J.; Wainberg, M.; Binder-Brynes, K.; Duvdevani, T. Vulnerability to Posttraumatic Stress Disorder in Adult Offspring of Holocaust Survivors. Am. J. Psychiatry 1998, 155, 1163–1171. [Google Scholar] [CrossRef]
- Franklin, T.; Russig, H.; Weiss, I.; Gräff, J.; Linder, N.; Michalon, A.; Vizi, S.; Mansuy, I. Epigenetic Transmission of the Impact of Early Stress Across Generations. Biol. Psychiatry 2010, 68. [Google Scholar] [CrossRef]
- Matthews, S.; Phillips, D. Transgenerational Inheritance of Stress Pathology. Exp. Neurol. 2012, 233. [Google Scholar] [CrossRef]
- Hillis, S.; Mercy, J.; Amobi, A.; Kress, H. Global prevalence of past-year violence against children: A systematic review and minimum estimates. Pediatrics 2016, 137, e20154079. [Google Scholar] [CrossRef] [Green Version]
- Stöckl, H.; March, L.; Pallitto, C.; Garcia-Moreno, C. Intimate partner violence among adolescents and young women: Prevalence and associated factors in nine countries: A cross-sectional study. BMC Public Health 2014, 14, 751. [Google Scholar] [CrossRef] [Green Version]
- Burkle, F.; Argent, A.; Kissoon, N. The Reality of Pediatric Emergency Mass Critical Care in the Developing World. Pediatr. Crit. Care Med. 2011, 12. [Google Scholar] [CrossRef]
- Wilkinson, R. Why Is Violence More Common Where Inequality Is Greater? Ann. N. Y. Acad. Sci. 2004, 1036. [Google Scholar] [CrossRef] [PubMed]
- De Boer, S.F. Animal models of excessive aggression: Implications for human aggression and violence. Curr. Opin. Psychol. 2018, 19, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Hong, W. Neural Circuit Mechanisms of Social Behavior. Neuron 2018, 98, 16–30. [Google Scholar] [CrossRef] [Green Version]
- Tottenham, N.; Sheridan, M.A. A review of adversity, the amygdala and the hippocampus: A consideration of developmental timing. Front. Hum. Neurosci. 2010, 3, 68. [Google Scholar] [CrossRef] [PubMed]
- Wellman, L.L.; Forcelli, P.A.; Aguilar, B.L.; Malkova, L. Bidirectional control of social behavior by activity within basolateral and central amygdala of primates. J. Neurosci. 2016, 36, 8746–8756. [Google Scholar] [CrossRef] [Green Version]
- Adolphs, R. What does the amygdala contribute to social cognition? Ann. N. Y. Acad. Sci. 2010, 1191, 42–61. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S.; Bowles, N.P.; Gray, J.D.; Hill, M.N.; Hunter, R.G.; Karatsoreos, I.N.; Nasca, C. Mechanisms of stress in the brain. Nat. Neurosci. 2015, 18, 1353–1363. [Google Scholar] [CrossRef]
- McEwen, B.S.; Nasca, C.; Gray, J.D. Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology 2016, 41, 3–23. [Google Scholar] [CrossRef] [Green Version]
- Charil, A.; Laplante, D.P.; Vaillancourt, C.; King, S. Prenatal stress and brain development. Brain Res. Rev. 2010, 65, 56–79. [Google Scholar] [CrossRef]
- Coussons-Read, M.E. Effects of prenatal stress on pregnancy and human development: Mechanisms and pathways. Obstet. Med. 2013, 6, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Huizink, A.C.; De Rooij, S.R. Prenatal stress and models explaining risk for psychopathology revisited: Generic vulnerability and divergent pathways. Dev. Psychopathol. 2018, 30, 1041–1062. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, M. Prenatal stressors in rodents: Effects on behavior. Neurobiol. Stress 2017, 6, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, S.; Dancause, K.; Turcotte-Tremblay, A.-M.; Veru, F.; Laplante, D.P. Using natural disasters to study the effects of prenatal maternal stress on child health and development. Birth Defects Res. C Embryo Today 2012, 96, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Lipner, E.; Murphy, S.K.; Ellman, L.M. Prenatal Maternal Stress and the Cascade of Risk to Schizophrenia Spectrum Disorders in Offspring. Curr. Psychiatry Rep. 2019, 21, 99. [Google Scholar] [CrossRef]
- Ellman, L.M.; Murphy, S.K.; Maxwell, S.D.; Calvo, E.M.; Cooper, T.; Schaefer, C.A.; Bresnahan, M.A.; Susser, E.S.; Brown, A.S. Maternal cortisol during pregnancy and offspring schizophrenia: Influence of fetal sex and timing of exposure. Schizophr. Res. 2019, 213, 15–22. [Google Scholar] [CrossRef]
- Levine, S.Z.; Levav, I.; Goldberg, Y.; Pugachova, I.; Becher, Y.; Yoffe, R. Exposure to genocide and the risk of schizophrenia: A population-based study. Psychol. Med. 2016, 46, 855–863. [Google Scholar] [CrossRef]
- Beversdorf, D.Q.; Stevens, H.E.; Margolis, K.G.; Van de Water, J. Prenatal Stress and Maternal Immune Dysregulation in Autism Spectrum Disorders: Potential Points for Intervention. Curr. Pharm. Des. 2019, 25, 4331–4343. [Google Scholar] [CrossRef]
- Walder, D.J.; Laplante, D.P.; Sousa-Pires, A.; Veru, F.; Brunet, A.; King, S. Prenatal maternal stress predicts autism traits in 61/2 year-old children: Project Ice Storm. Psychiatry Res. 2014, 219, 353–360. [Google Scholar] [CrossRef]
- Van Den Bergh, B.R.H.; Van Calster, B.; Smits, T.; Van Huffel, S.; Lagae, L. Antenatal maternal anxiety is related to HPA-axis dysregulation and self-reported depressive symptoms in adolescence: A prospective study on the fetal origins of depressed mood. Neuropsychopharmacology 2008, 33, 536–545. [Google Scholar] [CrossRef] [Green Version]
- Thomason, M.E. Development of Brain Networks In Utero: Relevance for Common Neural Disorders. Biol. Psychiatry 2020, 88. [Google Scholar] [CrossRef]
- Scheinost, D.; Kwon, S.H.; Lacadie, C.; Sze, G.; Sinha, R.; Constable, R.T.; Ment, L.R. Prenatal stress alters amygdala functional connectivity in preterm neonates. NeuroImage Clin. 2016, 12, 381–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ancatén González, C.; Gutiérrez-Rojas, C.; Bustamante Valdés, C. Maternal exercise reverses morphologic changes in amygdala neurons produced by prenatal stress. Neurol. Psychiatry Brain Res. 2017, 24, 36–42. [Google Scholar] [CrossRef]
- Gur, T.L.; Palkar, A.V.; Rajasekera, T.; Allen, J.; Niraula, A.; Godbout, J.; Bailey, M.T. Prenatal stress disrupts social behavior, cortical neurobiology and commensal microbes in adult male offspring. Behav. Brain Res. 2019, 359, 886. [Google Scholar] [CrossRef] [PubMed]
- Xing, G.; Carlton, J.; Jiang, X.; Wen, J.; Jia, M.; Li, H. Differential Expression of Brain Cannabinoid Receptors between Repeatedly Stressed Males and Females may Play a Role in Age and Gender-Related Difference in Traumatic Brain Injury: Implications from Animal Studies. Front. Neurol. 2014, 5, 161. [Google Scholar] [CrossRef] [PubMed]
- Voiculescu, S.E.; Le Duc, D.; Roșca, A.E.; Zeca, V.; Chiţimuș, D.M.; Arsene, A.L.; Drăgoi, C.M.; Nicolae, A.C.; Zăgrean, L.; Schöneberg, T.; et al. Behavioral and molecular effects of prenatal continuous light exposure in the adult rat. Brain Res. 2016, 1650, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.L.; Smith, R.M.; Edwards, K.S.; Givens, B.; Tilley, M.R.; Beversdorf, D.Q. Combined effect of maternal serotonin transporter genotype and prenatal stress in modulating offspring social interaction in mice. Int. J. Dev. Neurosci. 2010, 28, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Boersma, G.J.; Lee, R.S.; Cordner, Z.A.; Ewald, E.R.; Purcell, R.H.; Moghadam, A.A.; Tamashiro, K.L. Prenatal stress decreases Bdnf expression and increases methylation of Bdnf exon IV in rats. Epigenetics 2014, 9, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, D.E.; Rainnie, D.G. Prenatal Stress Alters the Development of Socioemotional Behavior and Amygdala Neuron Excitability in Rats. Neuropsychopharmacology 2015, 40, 2135–2145. [Google Scholar] [CrossRef] [Green Version]
- Kraszpulski, M.; Dickerson, P.A.; Salm, A.K. Prenatal stress affects the developmental trajectory of the rat amygdala. Stress 2006, 9, 85–95. [Google Scholar] [CrossRef]
- Lee, P.R.; Brady, D.L.; Shapiro, R.A.; Dorsa, D.M.; Koenig, J.I. Prenatal stress generates deficits in rat social behavior: Reversal by oxytocin. Brain Res. 2007, 1156, 152–167. [Google Scholar] [CrossRef] [Green Version]
- Roque, S.; Oliveira, T.G.; Nobrega, C.; Barreira-Silva, P.; Nunes-Alves, C.; Sousa, N.; Palha, J.A.; Correia-Neves, M. Interplay between Depressive-Like Behavior and the Immune System in an Animal Model of Prenatal Dexamethasone Administration. Front. Behav. Neurosci. 2011, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, S.; Coimbra, B.; Soares-Cunha, C.; Miguel Pêgo, J.; Sousa, N.; João Rodrigues, A. Dopaminergic modulation of affective and social deficits induced by prenatal glucocorticoid exposure. Neuropsychopharmacology 2013, 38, 2068–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuloaga, D.G.; Carbone, D.L.; Handa, R.J. Prenatal dexamethasone selectively decreases calretinin expression in the adult female lateral amygdala. Neurosci. Lett. 2012, 521, 109–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, J.; Jin, Y.; Choi, J.; Park, S.; Lee, T.H.; Lee, S.-R.; Chang, K.-T.; Hong, Y. Melatonin as a Novel Interventional Candidate for Fragile X Syndrome with Autism Spectrum Disorder in Humans. Int. J. Mol. Sci. 2017, 18, 1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Quera Salva, M.; Hartley, S.; Barbot, F.; C Alvarez, J.; Lofaso, F.; Guilleminault, C. Circadian Rhythms, Melatonin and Depression. Curr. Pharm. Des. 2011, 17, 1459–1470. [Google Scholar] [CrossRef]
- Ehrlich, D.E.; Neigh, G.N.; Bourke, C.H.; Nemeth, C.L.; Hazra, R.; Ryan, S.J.; Rowson, S.; Jairam, N.; Sholar, C.A.; Rainnie, D.G.; et al. Prenatal stress, regardless of concurrent escitalopram treatment, alters behavior and amygdala gene expression of adolescent female rats. Neuropharmacology 2015, 97, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Salari, A.-A.; Fatehi-Gharehlar, L.; Motayagheni, N.; Homberg, J.R. Fluoxetine normalizes the effects of prenatal maternal stress on depression- and anxiety-like behaviors in mouse dams and male offspring. Behav. Brain Res. 2016, 311, 354–367. [Google Scholar] [CrossRef]
- Pilkay, S.R.; Combs-Orme, T.; Tylavsky, F.; Bush, N.; Smith, A.K. Maternal trauma and fear history predict BDNF methylation and gene expression in newborns. PeerJ 2020, 8, e8858. [Google Scholar] [CrossRef]
- Franks, A.; Berry, K.; DeFranco, D. Prenatal Drug Exposure and Neurodevelopmental Programming of Glucocorticoid Signalling. J. Neuroendocrinol. 2020, 32. [Google Scholar] [CrossRef]
- Dow-Edwards, D.; Frank, A.; Wade, D.; Weedon, J.; Izenwasser, S. Sexually-dimorphic alterations in cannabinoid receptor density depend upon prenatal/early postnatal history. Neurotoxicol. Teratol. 2016, 58, 31–39. [Google Scholar] [CrossRef] [Green Version]
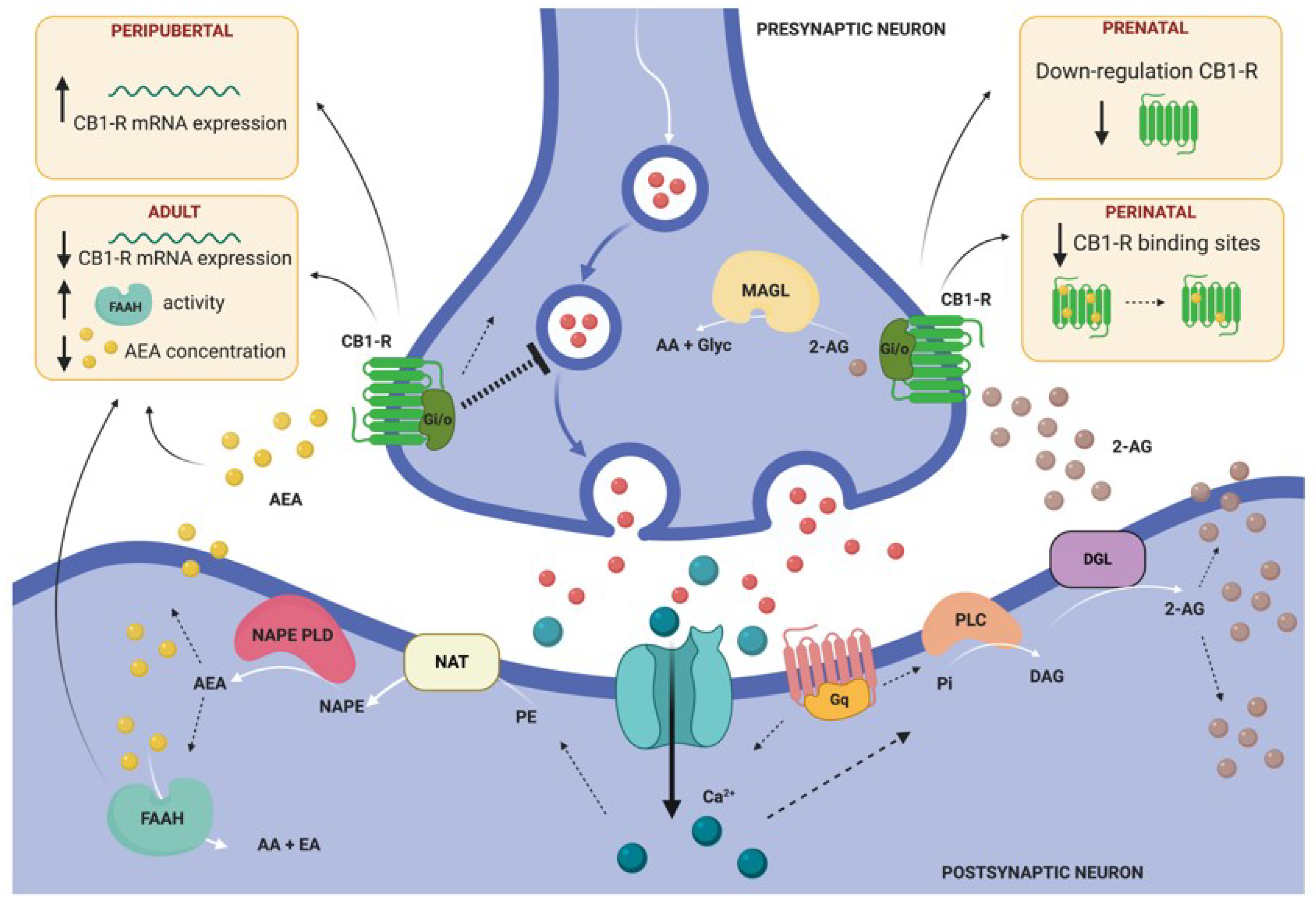
- Morena, M.; Patel, S.; Bains, J.S.; Hill, M.N. Neurobiological Interactions Between Stress and the Endocannabinoid System. Neuropsychopharmacology 2016, 41, 80–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Huppertz-Kessler, C.J.; Poeschl, J.; Hertel, R.; Unsicker, K.; Schenkel, J. Effects of a new postnatal stress model on monoaminergic neurotransmitters in rat brains. Brain Dev. 2012, 34, 274–279. [Google Scholar] [CrossRef]
- Guadagno, A.; Wong, T.P.; Walker, C.-D. Morphological and functional changes in the preweaning basolateral amygdala induced by early chronic stress associate with anxiety and fear behavior in adult male, but not female rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 81, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Szyf, M. The epigenetics of perinatal stress. Dialogues Clin. Neurosci. 2019, 21, 369. [Google Scholar] [CrossRef] [PubMed]
- Roth, T.L.; Matt, S.; Chen, K.; Blaze, J. Bdnf DNA methylation modifications in the hippocampus and amygdala of male and female rats exposed to different caregiving environments outside the homecage. Dev. Psychobiol. 2014, 56, 1755–1763. [Google Scholar] [CrossRef] [Green Version]
- Doherty, T.S.; Forster, A.; Roth, T.L. Global and gene-specific DNA methylation alterations in the adolescent amygdala and hippocampus in an animal model of caregiver maltreatment. Behav. Brain Res. 2016, 298, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Raineki, C.; Cortés, M.R.; Belnoue, L.; Sullivan, R.M. Effects of early-life abuse differ across development: Infant social behavior deficits are followed by adolescent depressive-like behaviors mediated by the amygdala. J. Neurosci. 2012, 32, 7758–7765. [Google Scholar] [CrossRef] [Green Version]
- Goldstein Ferber, S.; Trezza, V.; Weller, A. Early life stress and development of the endocannabinoid system: A bidirectional process in programming future coping. Dev. Psychobiol. 2019. [Google Scholar] [CrossRef]
- Hill, M.N.; Eiland, L.; Lee, T.T.; Hillard, C.J.; McEwen, B.S. Early Life Stress Alters the Developmental Trajectory of Corticolimbic Endocannabinoid Signaling in Male Rats. Neuropharmacology 2019, 146, 154. [Google Scholar] [CrossRef]
- Berman, A.K.; Lott, R.B.; Donaldson, S.T. Periodic maternal deprivation may modulate offspring anxiety-like behavior through mechanisms involving neuroplasticity in the amygdala. Brain Res. Bull. 2014, 101, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, E.K.Y.; Bian, Z.X.; Xu, H.X.; Sung, J.J.Y. Neonatal maternal separation increases brain-derived neurotrophic factor and tyrosine kinase receptor B expression in the descending pain modulatory system. NeuroSignals 2009, 17, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Karen, C.; Rajan, K.E. Social Behaviour and Epigenetic Status in Adolescent and Adult Rats: The Contribution of Early-Life Stressful Social Experience. Cell. Mol. Neurobiol. 2019, 39, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Poeggel, G.; Helmeke, C.; Abraham, A.; Schwabe, T.; Friedrich, P.; Braun, K. Juvenile emotional experience alters synaptic composition in the rodent cortex, hippocampus, and lateral amygdala. Proc. Natl. Acad. Sci. USA 2003, 100, 16137–16142. [Google Scholar] [CrossRef] [Green Version]
- Vela, G.; Martín, S.; García-Gil, L.; Crespo, J.A.; Ruiz-Gayo, M.; Javier Fernández-Ruiz, J.; García-Lecumberri, C.; Pélaprat, D.; Fuentes, J.A.; Ramos, J.A.; et al. Maternal exposure to δ9-tetrahydrocannabinol facilitates morphine self-administration behavior and changes regional binding to central μ opioid receptors in adult offspring female rats. Brain Res. 1998, 807, 101–109. [Google Scholar] [CrossRef]
- Keller, S.M.; Nowak, A.; Roth, T.L. Female pups receive more maltreatment from stressed dams. Dev. Psychobiol. 2019, 61, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Raineki, C.; Opendak, M.; Sarro, E.; Showler, A.; Bui, K.; McEwen, B.S.; Wilson, D.A.; Sullivan, R.M. During infant maltreatment, stress targets hippocampus, but stress with mother present targets amygdala and social behavior. Proc. Natl. Acad. Sci. USA 2019, 116, 22821–22832. [Google Scholar] [CrossRef] [PubMed]
- Higuera-Matas, A.; Ucha, M.; Ambrosio, E. Long-term consequences of perinatal and adolescent cannabinoid exposure on neural and psychological processes. Neurosci. Biobehav. Rev. 2015, 55, 119–146. [Google Scholar] [CrossRef] [PubMed]
- Spear, L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2000, 24, 417–463. [Google Scholar] [CrossRef]
- WHO. Definition of Key Terms; WHO: Geneva, Switzerland, 2013; Available online: https://www.who.int/hiv/pub/guidelines/arv2013/intro/keyterms/en/ (accessed on 5 July 2020).
- Tzanoulinou, S.; Riccio, O.; De Boer, M.W.; Sandi, C. Peripubertal stress-induced behavioral changes are associated with altered expression of genes involved in excitation and inhibition in the amygdale. Transl. Psychiatry 2014, 4, e410. [Google Scholar] [CrossRef]
- Stain, H.; Brønnick, K.; Hegelstad, W.T.; Joa, I.; Johannessen, J.; Langeveld, J.; Mawn, L.; Larsen, T. Impact of Interpersonal Trauma on the Social Functioning of Adults With First-Episode Psychosis. Schizophr. Bull. 2014, 40, 1491–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmier-Claus, J.; Berry, K.; Darrell-Berry, H.; Emsley, R.; Parker, S.; Drake, R.; Bucci, S. Childhood adversity and social functioning in psychosis: Exploring clinical and cognitive mediators. Psychiatry Res. 2016, 238, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Kilian, S.; Asmal, L.; Chiliza, B.; Olivier, M.; Phahladira, L.; Scheffler, F.; Seedat, S.; Marder, S.; Green, M.; Emsley, R. Childhood adversity and cognitive function in schizophrenia spectrum disorders and healthy controls: Evidence for an association between neglect and social cognition. Psychol. Med. 2018, 48, 2186–2193. [Google Scholar] [CrossRef] [PubMed]
- Heim, C.; Binder, E. Current Research Trends in Early Life Stress and Depression: Review of Human Studies on Sensitive Periods, Gene-Environment Interactions, and Epigenetics. Exp. Neurol. 2012, 233. [Google Scholar] [CrossRef] [PubMed]
- Widom, C.S.; DuMont, K.; Czaja, S.J. A prospective investigation of major depressive disorder and comorbidity in abused and neglected children grown up. Arch. Gen. Psychiatry 2007, 64, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Widom, C.S.; Maxfield, M.G. A prospective examination of risk for violence among abused and neglected children. Ann. N. Y. Acad. Sci. 1996, 794, 224–237. [Google Scholar] [CrossRef]
- Yeager, C.A.; Lewis, D.O. Mental illness, neuropsychologic deficits, child abuse, and violence. Child Adolesc. Psychiatr. Clin. N. Am. 2000, 9, 793–813. [Google Scholar] [CrossRef]
- Van der Kolk, B.A. The neurobiology of childhood trauma and abuse. Child Adolesc. Psychiatr. Clin. N. Am. 2003, 12, 293–317. [Google Scholar] [CrossRef]
- Malvaso, C.; Day, A.; Casey, S.; Corrado, R. Young Offenders, Maltreatment, and Trauma: A Pilot Study. Psychiatry Psychol. Law 2017, 24, 458–469. [Google Scholar] [CrossRef]
- Brydges, N.M.; Hall, J.; Best, C.; Rule, L.; Watkin, H.; Drake, A.J.; Lewis, C.; Thomas, K.L.; Hall, J. Childhood stress impairs social function through AVP-dependent mechanisms. Transl. Psychiatry 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Mikics, É.; Guirado, R.; Umemori, J.; Tóth, M.; Biró, L.; Miskolczi, C.; Balázsfi, D.; Zelena, D.; Castrén, E.; Haller, J.; et al. Social learning requires plasticity enhanced by fluoxetine through prefrontal Bdnf-TrkB signaling to limit aggression induced by post-weaning social isolation. Neuropsychopharmacology 2018, 43, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rau, A.R.; Chappell, A.M.; Butler, T.R.; Ariwodola, O.J.; Weiner, J.L. Increased Basolateral Amygdala Pyramidal Cell Excitability May Contribute to the Anxiogenic Phenotype Induced by Chronic Early-Life Stress. J. Neurosci. 2015, 35, 9730–9740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilabert-Juan, J.; Moltó, M.D.; Nacher, J. Post-weaning social isolation rearing influences the expression of molecules related to inhibitory neurotransmission and structural plasticity in the amygdala of adult rats. Brain Res. 2012, 1448, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Gómez, E.; Pérez-Rando, M.; Bellés, M.; Gilabert-Juan, J.; Llorens, J.V.; Carceller, H.; Bueno-Fernández, C.; García-Mompó, C.; Ripoll-Martínez, B.; Curto, Y.; et al. Early social isolation stress and perinatal nmda receptor antagonist treatment induce changes in the structure and neurochemistry of inhibitory neurons of the adult amygdala and prefrontal cortex. eNeuro 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Márquez, C.; Poirier, G.; Cordero, M.; Larsen, M.; Groner, A.; Marquis, J.; Magistretti, P.; Trono, D.; Sandi, C. Peripuberty Stress Leads to Abnormal Aggression, Altered Amygdala and Orbitofrontal Reactivity and Increased Prefrontal MAOA Gene Expression. Transl. Psychiatry 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Cordero, M.I.; Poirier, G.L.; Marquez, C.; Veenit, V.; Fontana, X.; Salehi, B.; Ansermet, F.; Sandi, C. Evidence for biological roots in the transgenerational transmission of intimate partner violence. Transl. Psychiatry 2012, 2, e106. [Google Scholar] [CrossRef] [Green Version]
- Tzanoulinou, S.; García-Mompó, C.; Castillo-Gómez, E.; Veenit, V.; Nacher, J.; Sandi, C. Long-Term Behavioral Programming Induced by Peripuberty Stress in Rats Is Accompanied by GABAergic-Related Alterations in the Amygdala. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Papilloud, A.; Veenit, V.; Tzanoulinou, S.; Riccio, O.; Zanoletti, O.; de Suduiraut, I.G.; Grosse, J.; Sandi, C. Peripubertal stress-induced heightened aggression: Modulation of the glucocorticoid receptor in the central amygdala and normalization by mifepristone treatment. Neuropsychopharmacology 2019, 44, 674. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Rosenkranz, J.A. Effects of Repeated Stress on Age-Dependent GABAergic Regulation of the Lateral Nucleus of the Amygdala. Neuropsychopharmacology 2016, 41, 2309–2323. [Google Scholar] [CrossRef]
- Jiang, X.; Xing, G.; Yang, C.; Verma, A.; Zhang, L.; Li, H. Stress Impairs 5-HT2A Receptor-Mediated Serotonergic Facilitation of GABA Release in Juvenile Rat Basolateral Amygdala. Neuropsychopharmacology 2009, 34. [Google Scholar] [CrossRef]
- Ferrari, A.J.; Somerville, A.J.; Baxter, A.J.; Norman, R.; Patten, S.B.; Vos, T.; Whiteford, H.A. Global variation in the prevalence and incidence of major depressive disorder: A systematic review of the epidemiological literature. Psychol. Med. 2013, 43, 471–481. [Google Scholar] [CrossRef]
- Tang, S.; Lu, L.; Zhang, L.; Hu, X.; Bu, X.; Li, H.; Hu, X.; Gao, Y.; Zeng, Z.; Gong, Q.; et al. Abnormal amygdala resting-state functional connectivity in adults and adolescents with major depressive disorder: A comparative meta-analysis. EBioMedicine 2018, 36, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Mothersill, O.; Donohoe, G. Neural effects of social environmental stress—An activation likelihood estimation meta-analysis. Psychol. Med. 2016, 46, 2015–2023. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Li, M.-X.; Xu, C.; Chen, H.-B.; An, S.-C.; Ma, X.-M.; Qiao, H.; Li, M.-X.; Xu, C.; Chen, H.-B.; et al. Dendritic Spines in Depression: What We Learned from Animal Models. Neural Plast. 2016, 2016, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestler, E.J.; Hyman, S.E. Animal models of neuropsychiatric disorders. Nat. Neurosci. 2010, 13, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Bennur, S.; Shankaranarayana Rao, B.S.; Pawlak, R.; Strickland, S.; McEwen, B.S.; Chattarji, S. Stress-induced spine loss in the medial amygdala is mediated by tissue-plasminogen activator. Neuroscience 2007, 144, 8–16. [Google Scholar] [CrossRef]
- Lau, T.; Bigio, B.; Zelli, D.; McEwen, B.; Nasca, C. Stress-induced structural plasticity of medial amygdala stellate neurons and rapid prevention by a candidate antidepressant. Mol. Psychiatry 2017, 22, 227. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Rosenkranz, J.A. Repeated restraint stress increases basolateral amygdala neuronal activity in an age-dependent manner. Neuroscience 2012, 226, 459. [Google Scholar] [CrossRef] [Green Version]
- Gilabert-Juan, J.; Castillo-Gomez, E.; Pérez-Rando, M.; Moltó, M.D.; Nacher, J. Chronic stress induces changes in the structure of interneurons and in the expression of molecules related to neuronal structural plasticity and inhibitory neurotransmission in the amygdala of adult mice. Exp. Neurol. 2011, 232, 33–40. [Google Scholar] [CrossRef]
- Pesarico, A.P.; Bueno-Fernandez, C.; Guirado, R.; Gómez-Climent, M.Á.; Curto, Y.; Carceller, H.; Nacher, J. Chronic Stress Modulates Interneuronal Plasticity: Effects on PSA-NCAM and Perineuronal Nets in Cortical and Extracortical Regions. Front. Cell. Neurosci. 2019, 13, 197. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.N.; Kumar, S.A.; Filipski, S.B.; Iverson, M.; Stuhr, K.L.; Keith, J.M.; Cravatt, B.F.; Hillard, C.J.; Chattarji, S.; McEwen, B.S. Disruption of fatty acid amide hydrolase activity prevents the effects of chronic stress on anxiety and amygdalar microstructure. Mol. Psychiatry 2013, 18, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.R.; Thakur, M.K. Correlation of ERα/ERβ expression with dendritic and behavioural changes in CUMS mice. Physiol. Behav. 2015, 145, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, Y.S.; Misquitta, K.A.; Rocco, B.R.; Prevot, T.D.; Knodt, A.R.; Ellegood, J.; Voineskos, A.N.; Lerch, J.P.; Hariri, A.R.; Sibille, E.; et al. Shifting priorities: Highly conserved behavioral and brain network adaptations to chronic stress across species. Transl. Psychiatry 2018, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachs, B.D.; Tran, H.L.; Folse, E.; Caron, M.G. Brain-region-specific molecular responses to maternal separation and social defeat stress in mice. Neuroscience 2018, 373, 122. [Google Scholar] [CrossRef]
- Nowacka-Chmielewska, M.M.; Kasprowska-Liśkiewicz, D.; Barski, J.J.; Obuchowicz, E.; Małecki, A. The behavioral and molecular evaluation of effects of social instability stress as a model of stress-related disorders in adult female rats. Stress 2017, 20, 549–561. [Google Scholar] [CrossRef]
- Mitra, R.; Sapolsky, R.M. Acute corticosterone treatment is sufficient to induce anxiety and amygdaloid dendritic hypertrophy. Proc. Natl. Acad. Sci. USA 2008, 105, 5573–5578. [Google Scholar] [CrossRef] [Green Version]
- Mitra, R.; Jadhav, S.; McEwen, B.S.; Vyas, A.; Chattarji, S. Stress duration modulates the spatiotemporal patterns of spine formation in the basolateral amygdala. Proc. Natl. Acad. Sci. USA 2005, 102, 9371–9376. [Google Scholar] [CrossRef] [Green Version]
- Varea, E.; Guirado, R.; Gilabert-Juan, J.; Martí, U.; Castillo-Gomez, E.; Blasco-Ibáñez, J.M.; Crespo, C.; Nacher, J. Expression of PSA-NCAM and synaptic proteins in the amygdala of psychiatric disorder patients. J. Psychiatr. Res. 2012, 46, 189–197. [Google Scholar] [CrossRef]
- Ancelin, M.-L.; Scali, J.; Norton, J.; Ritchie, K.; Dupuy, A.-M.; Chaudieu, I.; Ryan, J. Heterogeneity in HPA axis dysregulation and serotonergic vulnerability to depression. Psychoneuroendocrinology 2017, 77, 90–94. [Google Scholar] [CrossRef]
- Du, X.; Pang, T.Y. Is Dysregulation of the HPA-Axis a Core Pathophysiology Mediating Co-Morbid Depression in Neurodegenerative Diseases? Front. Psychiatry 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Lukas, M.; Neumann, I. Oxytocin and Vasopressin in Rodent Behaviors Related to Social Dysfunctions in Autism Spectrum Disorders. Behav. Brain Res. 2013, 251. [Google Scholar] [CrossRef] [PubMed]
- WHO. Proposed Working Definition of an Older Person in Africa for the MDS Project; WHO: Geneva, Switzerland, 2016; Available online: https://www.who.int/healthinfo/survey/ageingdefnolder/en/ (accessed on 7 July 2020).
- Sapolsky, R.M.; Krey, L.C.; McEwen, B.S. The neuroendocrinology of stress and aging: The glucocorticoid cascade hypothesis. Endocr. Rev. 1986, 7, 284–301. [Google Scholar] [CrossRef] [PubMed]
- Mcewen, B.S. Re-examination of the glucocorticoid hypothesis of stress and aging. Prog. Brain Res. 1992, 93, 365–383. [Google Scholar] [PubMed]
- Everaerd, D.; Klumpers, F.; Oude Voshaar, R.; Fernández, G.; Tendolkar, I. Acute Stress Enhances Emotional Face Processing in the Aging Brain. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 591–598. [Google Scholar] [CrossRef]
- Rubinow, M.J.; Drogos, L.L.; Juraska, J.M. Age-related dendritic hypertrophy and sexual dimorphism in rat basolateral amygdala. Neurobiol. Aging 2009, 30, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Sartin, J.; Mulchahey, J.J.; Segar, T.; Sheriff, S.; Herman, J.P.; Kasckow, J.W. Aging associated changes in amygdalar corticotropin-releasing hormone (CRH) and CRH-binding protein in Fischer 344 rats. Brain Res. 2006, 1073, 325–331. [Google Scholar] [CrossRef]
- Oizumi, H.; Kuriyama, N.; Imamura, S.; Tabuchi, M.; Omiya, Y.; Mizoguchi, K.; Kobayashi, H. Influence of aging on the behavioral phenotypes of C57BL/6J mice after social defeat. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Buechel, H.M.; Popovic, J.; Staggs, K.; Anderson, K.L.; Thibault, O.; Blalock, E.M. Aged rats are hypo-responsive to acute restraint: Implications for psychosocial stress in aging. Front. Aging Neurosci. 2014, 6. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Stressor | Stress Protocol | Molecular/Cellular | Behavior | References | ||
|---|---|---|---|---|---|---|
| Assessment | Age | Assessment | Age | |||
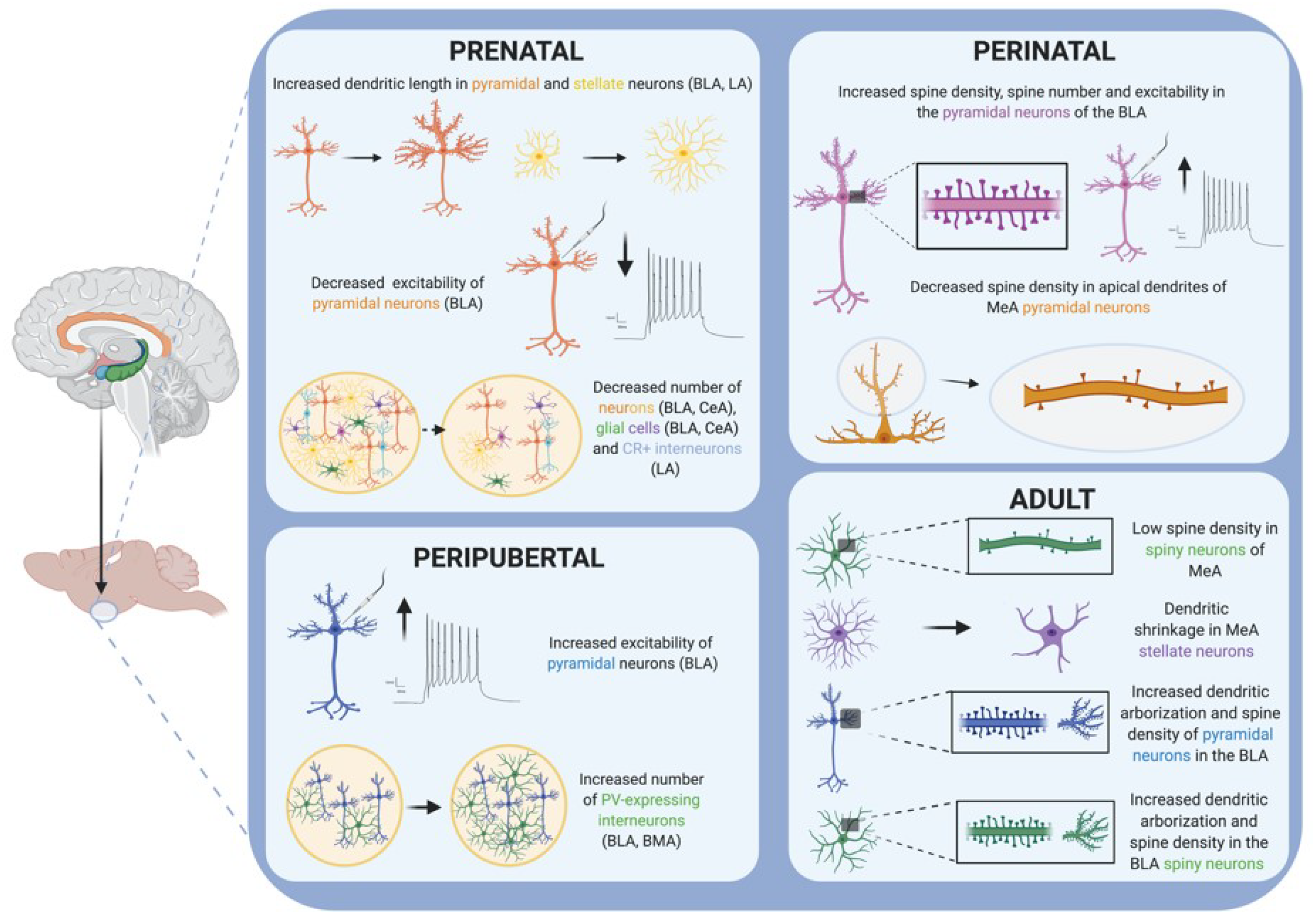
| Restraint or immobilization | From G14 to G21 (45 min; 3 times/day) | ↑ increased dendritic length of pyramidal and stellate neurons (BLA and LA) | P52 | [44] | ||
| From G10 to G16 (2 h/day) | ↓ serotonergic metabolism, ↑ CORT response to social interaction | P70 | ↓ social interaction | P60 to P70 | [45] | |
| 3 days (2 h/day) + tail-shocks | ↓ CB1-R in the amygdala | P60 | [46] | |||
| Continuous light exposure | From G12 to G21 | ↑ RORA in the amygdala | P72 | ↓ mobility and exploration | P60 to P72 | [47] |
| Unpredictable stress | Constant light exposure, fox-odor, restraint, cage changing, noise, wet bedding (from G6 to G21 in 5-HTT heterozygous mice) | ↓ social interaction,↓ social interest | P60 to P62 | [48] | ||
| Restraint, social defeat, cold exposure, forced swim, open field, lights-on overnight (from G14 to G21) | ↓ Bdnf expression, ↑ DNA methylation Bdnf exon IV, ↑ DNA methyltransferases in the amygdala | P21, P80 | [49] | |||
| Foot shocks (from G17 to G20) | ↓ excitability of BLA pyramidal neurons | P10, P14, P17, P21, P28, P60 | ↓ sociability, ↓ anxiety | P7, P17, P60 | [50] | |
| cage change and saline s.c. injection (from G14 to G21) | ↓ number of neurons and glia in BLA and CeA | P7, P45 P60 | [51] | |||
| Restraint, social defeat, cold exposure, forced swim, open field, lights-on overnight (from G14 to G21) | ↑ oxytocin receptor (OXTR) binding in the CeA | ↓ social interaction, = anxiety | P56 to P64 | [52] | ||
| Glucocorticoid administration | dexamethasone injections (0.4 mg/kg from G14 to G17 or 1 mg/kg from G18 to G19) | ↓ number of CR+ interneurons in the LA | P60 | depressive-like behavior, ↓ social interaction | P39, P50, P90, P120 | [53,54,55] |
| Stressor | Stress Protocol | Molecular/Cellular | Behavior | References | ||
|---|---|---|---|---|---|---|
| Assessment | Age | Assessment | Age | |||
| Bad caregiving conditions | Limited bedding (P1–P9) + 60 min immobilization (P10) or restraint (P20) | ↑ spine density, ↑ spine number and ↑ excitability (pyramidal neurons BLA) ↑ BLA volume | P10, P20, P18 to P22 | ↑ anxiety ↓ social contact | P60 to P78 | [66] |
| Caregiver maltreatment (stressed dam) outside the home cage (30 min/day from P1 to P7) | ↑ DNA methylation Bdnf exon IV (amygdala females) ↓ 5-hmC levels in (amygdala males) | P8, P30, P90 | ↑anxiety | P1 to P7 | [67,68,69] | |
| Insufficient bedding (from P8 to P12) | ↑ amygdala neuronal activity | P20, P45 | ↓ social behavior, depressive-like symptoms | P20, P45 | [70] | |
| Maternal separation | 3 h/day from P2 to P12 in the home cage (dam removed) | ↓ CB1-R binding sites in the amygdala (P40, P70), ↓ AEA content (P12, P14), ↑ 2-AG content (P12, P14) | P2, P12, P14, P40, P70 | [71,72] | ||
| 30 min/day on P5, P16 and P21 outside the home cage (pups removed) | ↑ BDNF protein levels in the amygdala | adult | ↓ anxiety | Infant, Adult | [73] | |
| 180 min/day from P2 to P14 outside the home cage (pups removed) | ↑ BDNF expression in the CeA ↑ TrkB expression in the CeA | adult | [74] | |||
| 3 h/day from P5 to P10 outside the home cage (pups removed) | ↑ DNA methyltransferases in the amygdala (P31) ↓ BDNF in the amygdala (P31, P86) | P31, P86 | ↓ social interaction | P30 to P31, P85 to P86 | [75] | |
| 1 h/day from P1 to P21 outside the home cage (pups removed and placed individually) | ↓ spine density in apical dendrites (pyramidal neurons MeA) | P21 | [76] | |||
| Drug administration (Δ9-THC) | 5 mg/kg (oral in mothers) from G5 to weaning age (P24) | ↓ density μ-opioid-R in posteromedial cortical amygdala (males) ↑ density μ-opioid-R in posteromedial cortical amygdala (females) | P70 | [77] | ||
| Stressor | Stress Protocol | Molecular/Cellular | Behavior | References | ||
|---|---|---|---|---|---|---|
| Assessment | Age | Assessment | Age | |||
| Post-weaning social isolation stress model (PWSI) | Individual cages from P21 to P82 | ↓ BDNF expression (amygdala) | P82 | pathological aggression, ↓ social communication | P82 | [9,94] |
| Individual cages from P28 to P109 | ↑ BLA pyramidal cell excitability | P101–P115 | ↑ anxiety | P101–P115 | [95] | |
| Individual cages from P21 to P90 | ↑ GAD67 protein (CeM, MeA, BLA) | P90 | [96] | |||
| Individual cages from P21 to P90 | ↑ BLA, BMA, Ce volume ↑ number of PV+ interneurons (BLA, BMA) ↓ PSA-NCAM protein (amygdala) ↓ VGLUT1/VGAT (LA, BLA) ↑ mRNA CB1-R (amygdala) | P90 | ↑ anxiety | P90 | [97] | |
| Unpredictable stress | fear-inducing stressors: open field, fox-odor, elevated platform (P28–P30, P34, P36, P40, P42) | ↑ mRNA NR1 (amygdala) ↓ mRNA GAD67 (amygdala) ↑ VGLUT1/VGAT (CeA) ↑ mRNA encoding GR and ↓ number of GR + cells (CeA) ↓ GAD and GABA-A receptor α3 (LA, BLA, BMA, MeA, CeA) | P90 | ↓ sociability, ↑ anxiety, ↑ novelty reactivity, pathological aggression | P90 | [83,98,99,100,101] |
| Repeated restraint stress | 20 min/day from P29 to P37 | ↓ GABAergic inhibition of LA projection neurons ↓ presynaptic GABA function and interneuron activity (LA) | P39 | ↑ anxiety | P38 | [102] |
| 2 h/day restraint session + 40 tail shocks /day (from P22 to P24) | ↓ serotoninergic modulation of GABAergic transmission (BLA), amygdala hyperexcitability | P24–P25 | [103] | |||
| Stressor | Stress Protocol | Molecular/Cellular | Behavior | References | ||
|---|---|---|---|---|---|---|
| Assessment | Age | Assessment | Age | |||
| CRS | 6 h/day for 21 days | ↓ spine density of spiny neurons in the MeA (GABAergic) | P90 | depression-like behavior | P90 | [107,109] |
| 2 h/day for 21 days | Dendritic shrinkage in MeA stellate neurons (↓ arborization, ↓ dendrite length) | P77 | ↓ social interaction ↑ anxiety-like behavior | P77 | [110] | |
| 1 h/day for 7 days | ↑ firing rate of BLA projecting neurons | P65 | ↑ anxiety-like behavior | P64 | [111] | |
| 1 h/day for 21 days | ↓ GAD67, synaptophysin and PSA-NCAM in the amygdala ↓ dendritic arborization of interneurons in the BLA | P112 | [112] | |||
| 6 h/day for 10 days | ↑ number of PV + neurons in the BLA | P112 | [113] | |||
| 6 h/day for 21 days | ↓ CB1-R expression in the amygdala ↑ FAAH activity, ↓ AEA amygdala ↑ dendritic arborization, complexity and spine density of pyramidal neurons in the BLA | P112 | ↑ anxiety-like behavior | P112 | [114] | |
| CUS (Forced swim, restraint, lights-on overnight, aversive smell, wet bedding, no bedding) | 2 weeks | ↑ dendritic arborization and spine density in the BLA spiny neurons (glutamatergic neurons) | P98 | depression-like behavior | P98 | [107,115] |
| 5 weeks | ↑ postsynaptic density-95 protein level in the amygdala and synaptic strengthening | P90 | ↑ behavioral emotionality | P90 | [116] | |
| Chronic social stress | Chronic defeat stress (5 min/day for 5 days) | ↑ POMC in the amygdala | P77 | [117] | ||
| Unpredictable chronic social instability (isolation and crowding, 3 h or 6 h/day for 28 days) | ↑ POMC, ↑ OXTR and ↓ AVPR1a in the amygdala | P88 | ↑ anxiety-like behavior | P88 | [118] | |
| Chronic exposure to exogenous corticosterone | 10 mg/kg (s.c.) for 1 or 10 days | ↑ dendritic length ↑ spine density (pyramidal neurons BLA) | P85 | ↑ anxiety-like behavior | P85 | [119] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mañas-Ojeda, A.; Ros-Bernal, F.; Olucha-Bordonau, F.E.; Castillo-Gómez, E. Becoming Stressed: Does the Age Matter? Reviewing the Neurobiological and Socio-Affective Effects of Stress throughout the Lifespan. Int. J. Mol. Sci. 2020, 21, 5819. https://doi.org/10.3390/ijms21165819
Mañas-Ojeda A, Ros-Bernal F, Olucha-Bordonau FE, Castillo-Gómez E. Becoming Stressed: Does the Age Matter? Reviewing the Neurobiological and Socio-Affective Effects of Stress throughout the Lifespan. International Journal of Molecular Sciences. 2020; 21(16):5819. https://doi.org/10.3390/ijms21165819
Chicago/Turabian StyleMañas-Ojeda, Aroa, Francisco Ros-Bernal, Francisco E. Olucha-Bordonau, and Esther Castillo-Gómez. 2020. "Becoming Stressed: Does the Age Matter? Reviewing the Neurobiological and Socio-Affective Effects of Stress throughout the Lifespan" International Journal of Molecular Sciences 21, no. 16: 5819. https://doi.org/10.3390/ijms21165819
APA StyleMañas-Ojeda, A., Ros-Bernal, F., Olucha-Bordonau, F. E., & Castillo-Gómez, E. (2020). Becoming Stressed: Does the Age Matter? Reviewing the Neurobiological and Socio-Affective Effects of Stress throughout the Lifespan. International Journal of Molecular Sciences, 21(16), 5819. https://doi.org/10.3390/ijms21165819