Connexin Mutants Compromise the Lens Circulation and Cause Cataracts through Biomineralization



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Lens and Its Specialized Cells
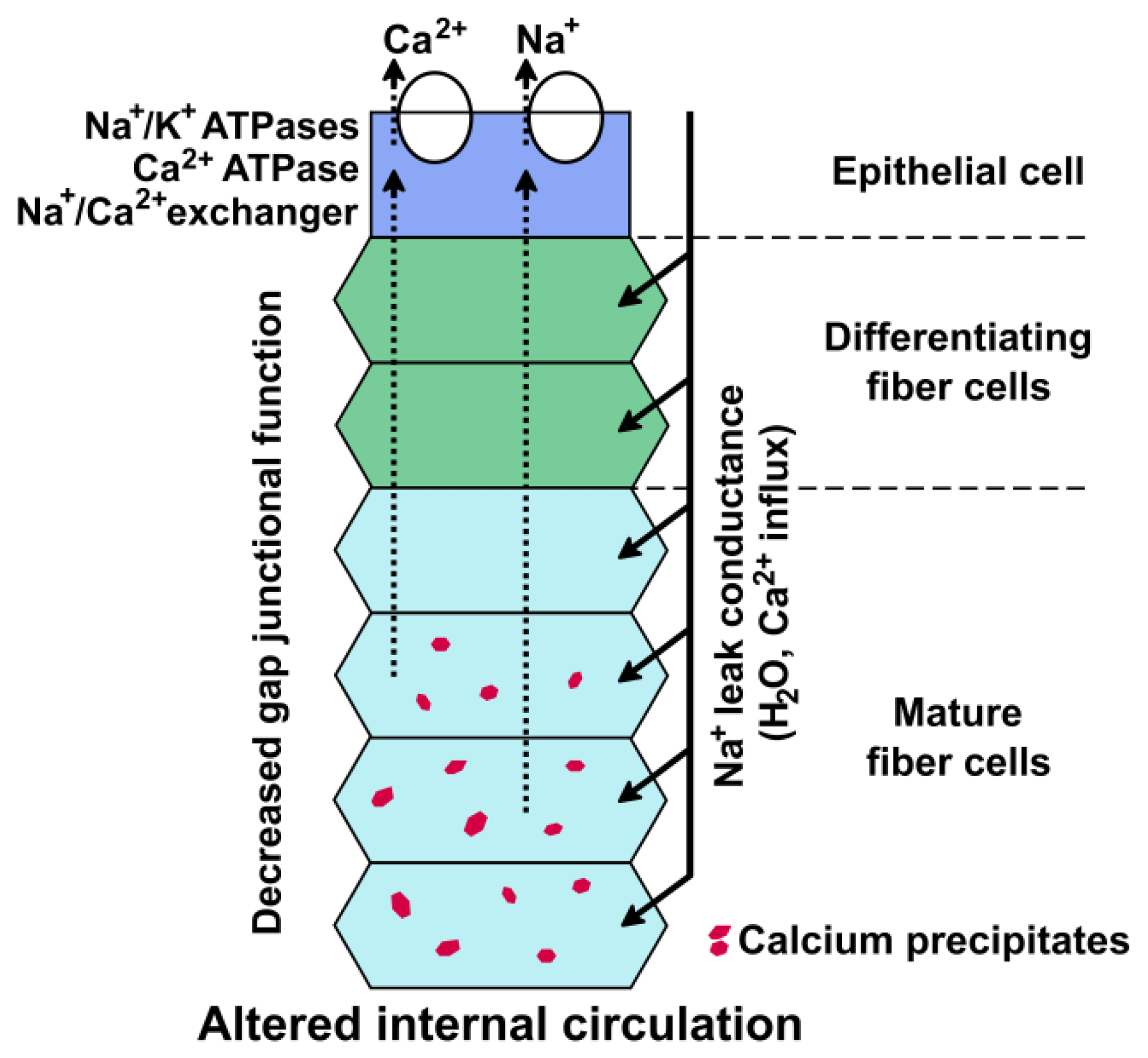
3. Intercellular Communication and Healthy Survival of the Lens
4. Lens Disease and Connexins
5. Calcium Ions in the Normal and Cataractous Lens
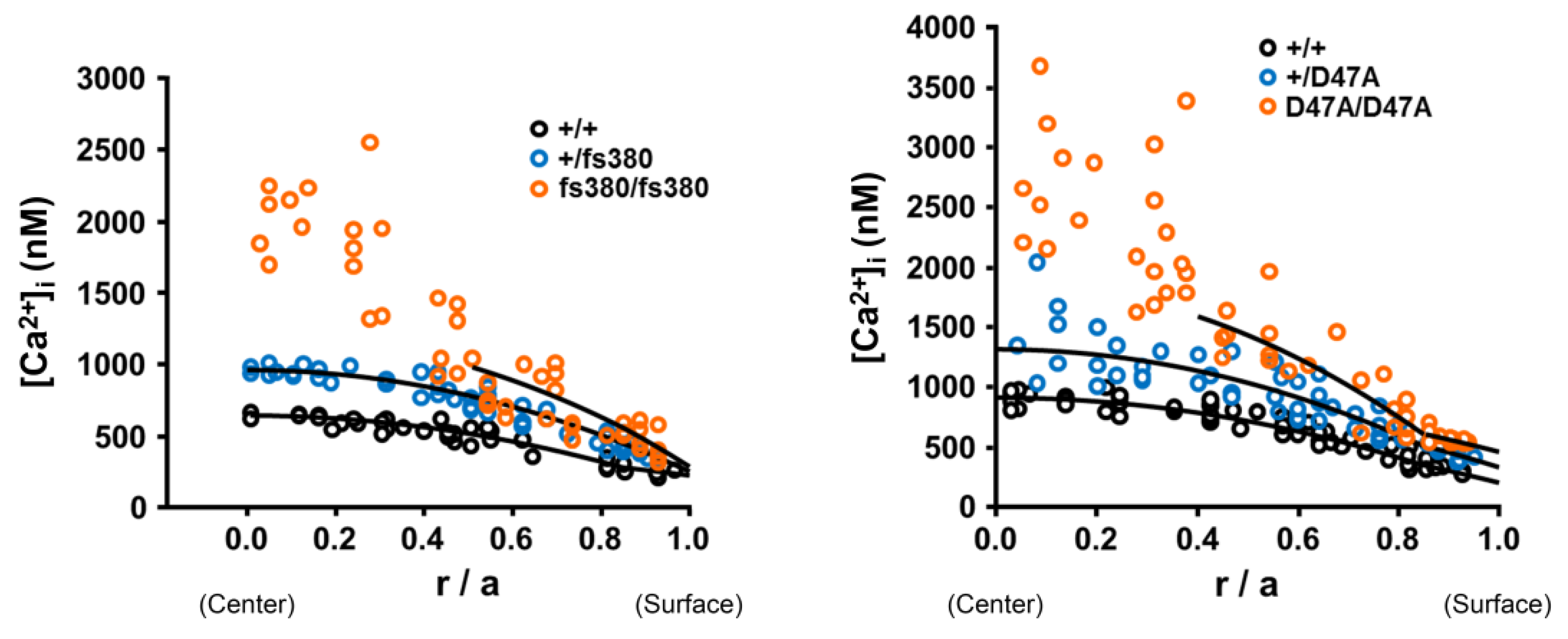
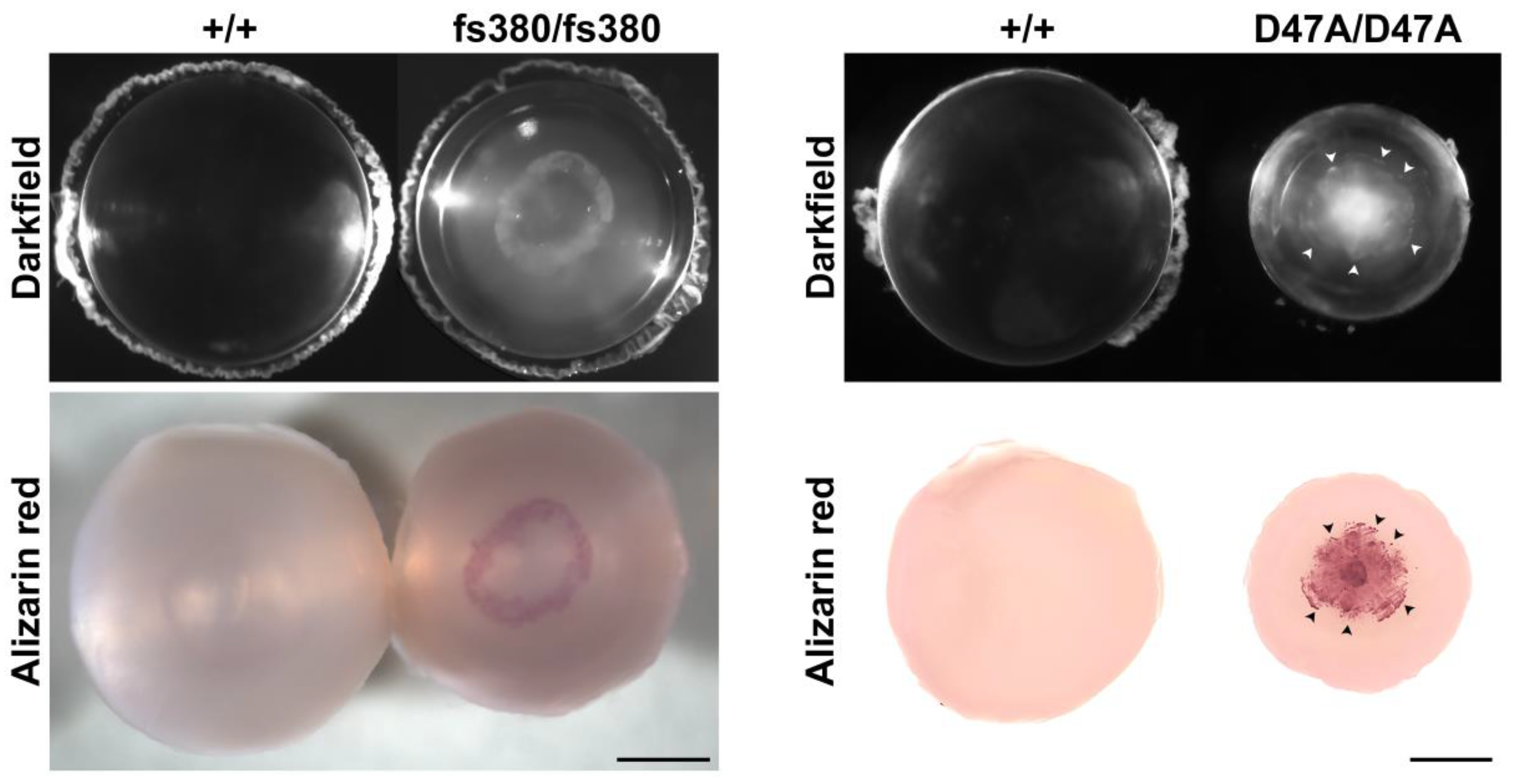
6. The Cataractous Lens: A Mineralized Organ?
7. Cataracts May Result from Biomineralization in the Lens
8. Potential Generality of the Biomineralization Model for Cataract Formation
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| [Ca2+]i | Intracellular concentration of calcium ions |
| Cx43 | Connexin43 |
| Cx46 | Connexin46 |
| Cx50 | Connexin50 |
| K6W-Ub | Ubiquitin K6W mutant |
| Ksp | Solubility product constant |
| NO | Nitric oxide |
References
- Delaye, M.; Tardieu, A. Short-range order of crystallin proteins accounts for eye lens transparency. Nature 1983, 302, 415–417. [Google Scholar] [CrossRef] [PubMed]
- Ponce, A.; Sorensen, C.; Takemoto, L. Role of short-range protein interactions in lens opacifications. Mol. Vis. 2006, 12, 879–884. [Google Scholar] [PubMed]
- Gao, J.; Minogue, P.J.; Beyer, E.C.; Mathias, R.T.; Berthoud, V.M. Disruption of the lens circulation causes calcium accumulation and precipitates in connexin mutant mice. Am. J. Physiol. Cell Physiol. 2018, 314, C492–C503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathias, R.T.; Kistler, J.; Donaldson, P. The lens circulation. J. Membr. Biol. 2007, 216, 1–16. [Google Scholar] [CrossRef]
- Resnikoff, S.; Pascolini, D.; Etya’ale, D.; Kocur, I.; Pararajasegaram, R.; Pokharel, G.P.; Mariotti, S.P. Global data on visual impairment in the year 2002. Bull. World Health Organ. 2004, 82, 844–851. [Google Scholar]
- Francis, P.J.; Moore, A.T. Genetics of childhood cataract. Curr. Opin. Ophthalmol. 2004, 15, 10–15. [Google Scholar] [CrossRef]
- Shiels, A.; Bennett, T.M.; Hejtmancik, J.F. Cat-Map: Putting cataract on the map. Mol. Vis. 2010, 16, 2007–2015. [Google Scholar]
- Berthoud, V.M.; Ngezahayo, A. Focus on lens connexins. BMC Cell Biol. 2017, 18 (Suppl. 1), 6. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.; Li, E.; Klier, G.; Huang, Q.; Wu, Y.; Lei, H.; Kumar, N.M.; Horwitz, J.; Gilula, N.B. Disruption of α3 connexin gene leads to proteolysis and cataractogenesis in mice. Cell 1997, 91, 833–843. [Google Scholar] [CrossRef] [Green Version]
- White, T.W.; Goodenough, D.A.; Paul, D.L. Targeted ablation of connexin50 in mice results in microphthalmia and zonular pulverulent cataracts. J. Cell Biol. 1998, 143, 815–825. [Google Scholar] [CrossRef]
- Rong, P.; Wang, X.; Niesman, I.; Wu, Y.; Benedetti, L.E.; Dunia, I.; Levy, E.; Gong, X. Disruption of Gja8 (α8 connexin) in mice leads to microphthalmia associated with retardation of lens growth and lens fiber maturation. Development 2002, 129, 167–174. [Google Scholar]
- Steele, E.C., Jr.; Lyon, M.F.; Favor, J.; Guillot, P.V.; Boyd, Y.; Church, R.L. A mutation in the connexin 50 (Cx50) gene is a candidate for the No2 mouse cataract. Curr. Eye Res. 1998, 17, 883–889. [Google Scholar] [CrossRef]
- Graw, J.; Löster, J.; Soewarto, D.; Fuchs, H.; Meyer, B.; Reis, A.; Wolf, E.; Balling, R.; Hrabé de Angelis, M. Characterization of a mutation in the lens-specific MP70 encoding gene of the mouse leading to a dominant cataract. Exp. Eye Res. 2001, 73, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.; Wang, X.; Hawes, N.L.; Ojakian, R.; Davisson, M.T.; Lo, W.-K.; Gong, X. A Gja8 (Cx50) point mutation causes an alteration of α3 connexin (Cx46) in semi-dominant cataracts of Lop10 mice. Hum. Mol. Genet. 2002, 11, 507–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, C.-h.; Chang, B.; DeRosa, A.M.; Cheng, C.; White, T.W.; Gong, X. Cataracts and microphthalmia caused by a Gja8 mutation in extracellular loop 2. PLoS ONE 2012, 7, e52894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoud, V.M.; Minogue, P.J.; Yu, H.; Schroeder, R.; Snabb, J.I.; Beyer, E.C. Connexin50D47A decreases levels of fiber cell connexins and impairs lens fiber cell differentiation. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7614–7622. [Google Scholar] [CrossRef]
- Berthoud, V.M.; Minogue, P.J.; Yu, H.; Snabb, J.I.; Beyer, E.C. Connexin46fs380 causes progressive cataracts. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6639–6648. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.; Baldo, G.J.; Kumar, N.M.; Gilula, N.B.; Mathias, R.T. Gap junctional coupling in lenses lacking α3 connexin. Proc. Natl. Acad. Sci. USA 1998, 95, 15303–15308. [Google Scholar] [CrossRef] [Green Version]
- Baldo, G.J.; Gong, X.; Martinez-Wittinghan, F.J.; Kumar, N.M.; Gilula, N.B.; Mathias, R.T. Gap junctional coupling in lenses from α8 connexin knockout mice. J. Gen. Physiol. 2001, 118, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Minogue, P.J.; Gao, J.; Zoltoski, R.K.; Novak, L.A.; Mathias, R.T.; Beyer, E.C.; Berthoud, V.M. Physiological and optical alterations precede the appearance of cataracts in Cx46fs380 mice. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4366–4374. [Google Scholar] [CrossRef] [Green Version]
- Berthoud, V.M.; Gao, J.; Minogue, P.J.; Jara, O.; Mathias, R.T.; Beyer, E.C. The connexin50D47A mutant causes cataracts by calcium precipitation. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2336–2346. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Lyu, L.; Chin, D.; Gao, J.; Sun, X.; Shang, F.; Caceres, A.; Chang, M.-L.; Rowan, S.; Peng, J.; et al. Altered ubiquitin causes perturbed calcium homeostasis, hyperactivation of calpain, dysregulated differentiation, and cataract. Proc. Natl. Acad. Sci. USA 2015, 112, 1071–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Sun, X.; Moore, L.C.; White, T.W.; Brink, P.R.; Mathias, R.T. Lens intracellular hydrostatic pressure is generated by the circulation of sodium and modulated by gap junction coupling. J. Gen. Physiol. 2011, 137, 507–520. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Sun, X.; Martinez-Wittinghan, F.J.; Gong, X.; White, T.W.; Mathias, R.T. Connections between connexins, calcium, and cataracts in the lens. J. Gen. Physiol. 2004, 124, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Cheng, C.; Xia, C.-h.; White, T.W.; Fletcher, D.A.; Gong, X. Connexin mediated cataract prevention in mice. PLoS ONE 2010, 5, e12624. [Google Scholar] [CrossRef] [Green Version]
- Shahidullah, M.; Mandal, A.; Delamere, N.A. TRPV4 in porcine lens epithelium regulates hemichannel-mediated ATP release and Na-K-ATPase activity. Am. J. Physiol. Cell Physiol. 2012, 302, C1751–C1761. [Google Scholar] [CrossRef] [Green Version]
- Maddala, R.; Nagendran, T.; de Ridder, G.G.; Schey, K.L.; Rao, P.V. L-type calcium channels play a critical role in maintaining lens transparency by regulating phosphorylation of aquaporin-0 and myosin light chain and expression of connexins. PLoS ONE 2013, 8, e64676. [Google Scholar] [CrossRef] [Green Version]
- Martínez-García, M.C.; Martínez, T.; Pañeda, C.; Gallego, P.; Jimenez, A.I.; Merayo, J. Differential expression and localization of transient receptor potential vanilloid 1 in rabbit and human eyes. Histol. Histopathol. 2013, 28, 1507–1516. [Google Scholar]
- Bennett, T.M.; Mackay, D.S.; Siegfried, C.J.; Shiels, A. Mutation of the melastatin-related cation channel, TRPM3, underlies inherited cataract and glaucoma. PLoS ONE 2014, 9, e104000. [Google Scholar] [CrossRef]
- De Maria, A.; Zhao, H.; Bassnett, S. Expression of potassium-dependent sodium-calcium exchanger in the murine lens. Exp. Eye Res. 2018, 167, 18–24. [Google Scholar] [CrossRef]
- Sharma, K.K.; Santhoshkumar, P. Lens aging: Effects of crystallins. Biochim. Biophys. Acta 2009, 1790, 1095–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Zhu, X.; Lu, Y. The proteome of cataract markers: Focus on crystallins. Adv. Clin. Chem. 2018, 86, 179–210. [Google Scholar] [PubMed]
- Truscott, R.J.W.; Friedrich, M.G. Molecular processes implicated in human age-related nuclear cataract. Investig. Ophthalmol. Vis. Sci. 2019, 60, 5007–5021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burge, W.E. Analysis of the ash of the normal and the cataractous lens. Arch. Ophthalmol. 1909, 38, 435–450. [Google Scholar]
- Duncan, G.; van Heyningen, R. Distribution of non-diffusible calcium and sodium in normal and cataractous human lenses. Exp. Eye Res. 1977, 25, 183–193. [Google Scholar] [CrossRef]
- Duncan, G.; Jacob, T.J. Calcium and the physiology of cataract. Ciba Found. Symp. 1984, 106, 132–152. [Google Scholar]
- Hightower, K.R.; Reddy, V.N. Calcium content and distribution in human cataract. Exp. Eye Res. 1982, 34, 413–421. [Google Scholar] [CrossRef]
- Zimmerman, L.E.; Johnson, F.B. Calcium oxalate crystals within ocular tissues. A clinicopathologic and histochemical study. AMA Arch. Ophthalmol. 1958, 60, 372–383. [Google Scholar] [CrossRef]
- Bron, A.J.; Habgood, J.O. Morgagnian cataract. Trans. Ophthalmol. Soc. U.K. 1976, 96, 265–277. [Google Scholar]
- Pau, H. Spheroliths in the lens. Klin. Monbl. Augenheilkd. 1984, 184, 159–162. [Google Scholar] [CrossRef]
- Weiner, S.; Dove, P.M. An overview of biomineralization processes and the problem of the vital effect. Rev. Mineral. Geochem. 2003, 54, 1–29. [Google Scholar] [CrossRef]
- Kawasaki, K.; Buchanan, A.V.; Weiss, K.M. Biomineralization in humans: Making the hard choices in life. Annu. Rev. Genet. 2009, 43, 119–142. [Google Scholar] [CrossRef] [PubMed]
- Shearer, D.; Ens, W.; Standing, K.; Valdimarsson, G. Posttranslational modifications in lens fiber connexins identified by off-line-HPLC MALDI-quadrupole time-of-flight mass spectrometry. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1553–1562. [Google Scholar] [CrossRef]
- Yin, X.; Liu, J.; Jiang, J.X. Lens fiber connexin turnover and caspase-3-mediated cleavage are regulated alternately by phosphorylation. Cell Commun. Adhes. 2008, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Schey, K.L. Phosphorylation and truncation sites of bovine lens connexin 46 and connexin 50. Exp. Eye Res. 2009, 89, 898–904. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Han, J.; David, L.L.; Schey, K.L. Proteomics and phosphoproteomics analysis of human lens fiber cell membranes. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.B.; Haddad, B.G.; O’Neill, S.E.; Chorev, D.S.; Yoshioka, C.C.; Robinson, C.V.; Zuckerman, D.M.; Reichow, S.L. Structure of native lens connexin 46/50 intercellular channels by cryo-EM. Nature 2018, 564, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Retamal, M.A.; Orellana, V.P.; Arévalo, N.J.; Rojas, C.G.; Arjona, R.J.; Alcaíno, C.A.; González, W.; Canan, J.G.; Moraga-Amaro, R.; Stehberg, J.; et al. Cx46 hemichannel modulation by nitric oxide: Role of the fourth transmembrane helix cysteine and its possible involvement in cataract formation. Nitric Oxide 2019, 86, 54–62. [Google Scholar] [CrossRef]
- Retamal, M.A.; Fiori, M.C.; Fernandez-Olivares, A.; Linsambarth, S.; Peña, F.; Quintana, D.; Stehberg, J.; Altenberg, G.A. 4-Hydroxynonenal induces Cx46 hemichannel inhibition through its carbonylation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158705. [Google Scholar] [CrossRef]
- Berthoud, V.M.; Beyer, E.C.; Kurata, W.E.; Lau, A.F.; Lampe, P.D. The gap-junction protein connexin 56 is phosphorylated in the intracellular loop and the carboxy-terminal region. Eur. J. Biochem. 1997, 244, 89–97. [Google Scholar] [CrossRef]
- Cheng, H.-L.; Louis, C.F. Endogenous casein kinase I catalyzes the phosphorylation of the lens fiber cell connexin49. Eur. J. Biochem. 1999, 263, 276–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoud, V.M.; Westphale, E.M.; Grigoryeva, A.; Beyer, E.C. PKC isoenzymes in the chicken lens and TPA-induced effects on intercellular communication. Investig. Ophthalmol. Vis. Sci. 2000, 41, 850–858. [Google Scholar]
- Cheng, H.-L.; Louis, C.F. Functional effects of casein kinase I-catalyzed phosphorylation on lens cell-to-cell coupling. J. Membr. Biol. 2001, 181, 21–30. [Google Scholar] [CrossRef]
- Liu, J.; Ek Vitorin, J.F.; Weintraub, S.T.; Gu, S.; Shi, Q.; Burt, J.M.; Jiang, J.X. Phosphorylation of connexin 50 by protein kinase A enhances gap junction and hemichannel function. J. Biol. Chem. 2011, 286, 16914–16928. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.M.; Wang, H.Z.; Lin, R.Z.; Brink, P.R.; White, T.W. Differential regulation of Connexin50 and Connexin46 by PI3K signaling. FEBS Lett. 2015, 589, 1340–1345. [Google Scholar] [CrossRef] [Green Version]
- Berthoud, V.M.; Bassnett, S.; Beyer, E.C. Cultured chicken embryo lens cells resemble differentiating fiber cells in vivo and contain two kinetic pools of connexin56. Exp. Eye Res. 1999, 68, 475–484. [Google Scholar] [CrossRef]
- Yin, X.; Jedrzejewski, P.T.; Jiang, J.X. Casein kinase II phosphorylates lens connexin 45.6 and is involved in its degradation. J. Biol. Chem. 2000, 275, 6850–6856. [Google Scholar] [CrossRef] [Green Version]
- Eckert, R. pH gating of lens fibre connexins. Pflüg. Arch. 2002, 443, 843–851. [Google Scholar] [CrossRef]
- Lin, J.S.; Eckert, R.; Kistler, J.; Donaldson, P. Spatial differences in gap junction gating in the lens are a consequence of connexin cleavage. Eur. J. Cell Biol. 1998, 76, 246–250. [Google Scholar]
- Stergiopoulos, K.; Alvarado, J.L.; Mastroianni, M.; Ek-Vitorin, J.F.; Taffet, S.M.; Delmar, M. Hetero-domain interactions as a mechanism for the regulation of connexin channels. Circ. Res. 1999, 84, 1144–1155. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Berthoud, V.M.; Beyer, E.C.; Ebihara, L. Functional role of the carboxyl terminal domain of human connexin 50 in gap junctional channels. J. Membr. Biol. 2002, 186, 101–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeRosa, A.M.; Mui, R.; Srinivas, M.; White, T.W. Functional characterization of a naturally occurring Cx50 truncation. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4474–4481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Gu, S.; Jiang, J.X. The development-associated cleavage of lens connexin 45.6 by caspase-3-like protease is regulated by casein kinase II-mediated phosphorylation. J. Biol. Chem. 2001, 276, 34567–34572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Wang, H.; Sun, X.; Varadaraj, K.; Li, L.; White, T.W.; Mathias, R.T. The effects of age on lens transport. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7174–7187. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.S. The biomineralization proteome: Protein complexity for a complex bioceramic assembly process. Proteomics 2019, 19, e1900036. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berthoud, V.M.; Gao, J.; Minogue, P.J.; Jara, O.; Mathias, R.T.; Beyer, E.C. Connexin Mutants Compromise the Lens Circulation and Cause Cataracts through Biomineralization. Int. J. Mol. Sci. 2020, 21, 5822. https://doi.org/10.3390/ijms21165822
Berthoud VM, Gao J, Minogue PJ, Jara O, Mathias RT, Beyer EC. Connexin Mutants Compromise the Lens Circulation and Cause Cataracts through Biomineralization. International Journal of Molecular Sciences. 2020; 21(16):5822. https://doi.org/10.3390/ijms21165822
Chicago/Turabian StyleBerthoud, Viviana M., Junyuan Gao, Peter J. Minogue, Oscar Jara, Richard T. Mathias, and Eric C. Beyer. 2020. "Connexin Mutants Compromise the Lens Circulation and Cause Cataracts through Biomineralization" International Journal of Molecular Sciences 21, no. 16: 5822. https://doi.org/10.3390/ijms21165822
APA StyleBerthoud, V. M., Gao, J., Minogue, P. J., Jara, O., Mathias, R. T., & Beyer, E. C. (2020). Connexin Mutants Compromise the Lens Circulation and Cause Cataracts through Biomineralization. International Journal of Molecular Sciences, 21(16), 5822. https://doi.org/10.3390/ijms21165822