The Anti-Inflammatory Effects of Angiogenin in an Endotoxin Induced Uveitis in Rats


Abstract
:1. Introduction
2. Results
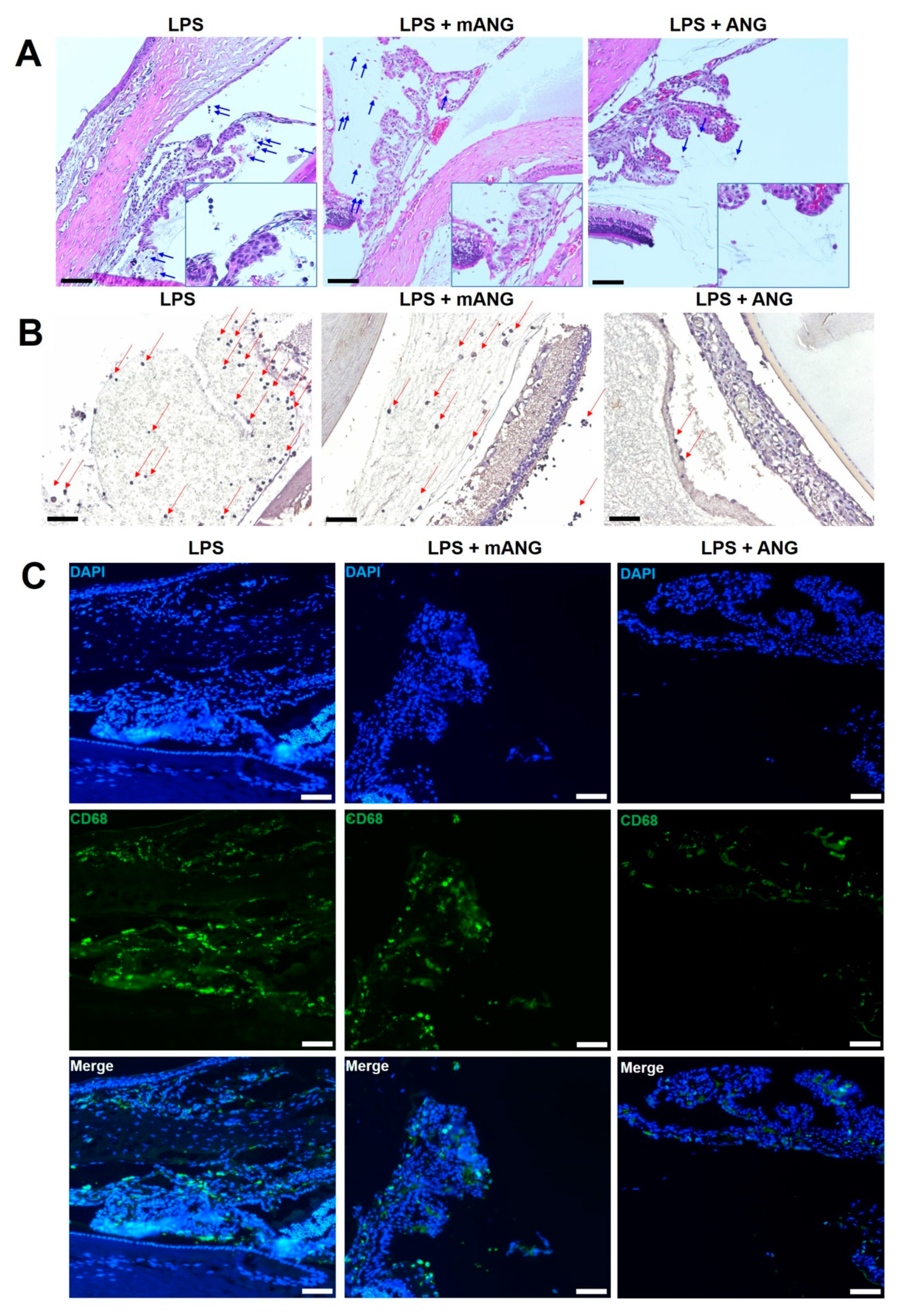
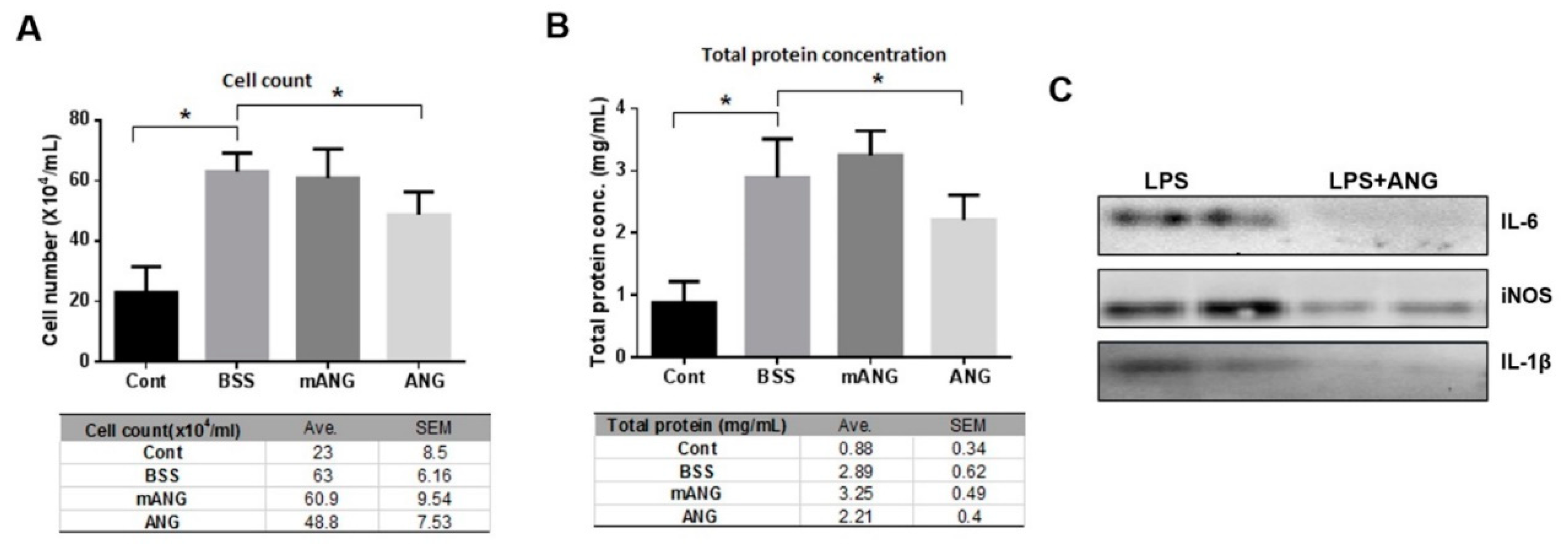
2.1. ANG Reduced Infiltrating Inflammatory Cells, Protein Content, and Proinflammatory Cytokines in the Anterior Chamber
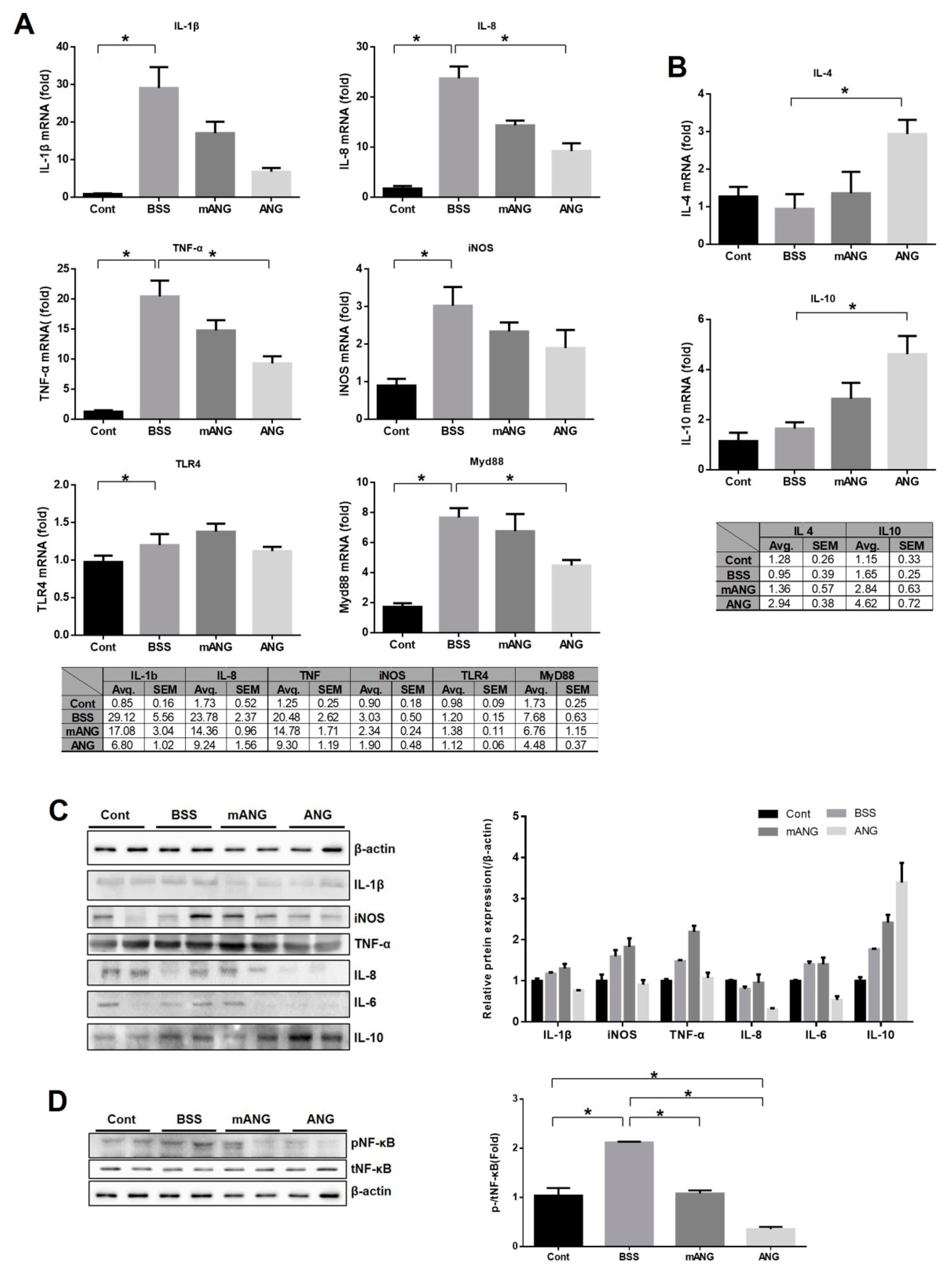
2.2. ANG Inhibits mRNA Expression of Proinflammatory Cytokines and Myd88 While Promoting mRNA Expression of Anti-Inflammatory Cytokines in Ocular Tissues
2.3. ANG Inhibits Nuclear Translocation of NF-κB in the Ciliary Body
3. Discussion
4. Materials and Methods
4.1. Study Approval and Material Preparation
4.2. Induction of Anterior Uveitis (EIU) in Rats and Topical Treatment with ANG Eye Drops
4.3. Cell Counting and Quantification of Protein Concentration in Aqueous Humor of EIU Rats
4.4. Histopathological Evaluation of EIU Rats
4.5. Purification of Total RNA and Real-Time qRT-PCR from Ocular Tissue of EIU Rats
4.6. Western Blots of Eyes from EIU Rats
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mérida, S.; Sancho-Tello, M.; Almansa, I.; Desco, C.; Peris, C.; Moreno, M.L.; Villar, V.M.; Navea, A.; Bosch-Morell, F. Bevacizumab Diminishes Inflammation in an Acute Endotoxin-Induced Uveitis Model. Front Pharm. 2018, 9, 649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, M.C.; Wu, P.C.; Lin, S.P.; Wu, C.Y.; Wang, P.H.; Chen, C.T.; Chen, B.Y. Danshensu Decreases UVB-Induced Corneal Inflammation in an Experimental Mouse Model via Oral Administration. Curr. Eye Res. 2018, 43, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.H.; McCluskey, P.J.; Wakefield, D. Toll-Like Receptors in Ocular Immunity and the Immunopathogenesis of Inflammatory Eye Disease. Brz. J. Ophthalmol. 2006, 90, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trittibach, P.; Barker, S.E.; Broderick, C.A.; Natkunarajah, M.; Duran, Y.; Robbie, S.J.; Bainbridge, J.W.; Smith, A.J.; Sarra, G.M.; Dick, A.D.; et al. Lentiviral-Vector-Mediated Expression of Murine IL-1 Receptor Antagonist or IL-10 Reduces the Severity of Endotoxin-Induced Uveitis. Gene Ther. 2008, 15, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.J.; Chu, K.O.; Yip, Y.W.; Li, W.Y.; Yang, Y.P.; Chan, K.P.; Ren, J.L.; Chan, S.O.; Pang, C.P. Green Tea Extract Treatment Alleviates Ocular Inflammation in a Rat Model of Endotoxin-Induced Uveitis. PloS ONE 2014, 9, e103995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durrani, O.M.; Tehrani, N.N.; Marr, J.E.; Moradi, P.; Stavrou, P.; Murray, P.I. Degree, Duration, and Causes of Visual Loss in Uveitis. Br. J. Ophthalmol. 2004, 88, 1159–1162. [Google Scholar] [CrossRef] [Green Version]
- Nussenblatt, R.B. The Natural History of Uveitis. Int. Ophthalmol. 1990, 14, 303–308. [Google Scholar] [CrossRef]
- Read, R.W. Uveitis: Advances in Understanding of Pathogenesis and Treatment. Curr. Rheumatol. Rep. 2006, 8, 260–266. [Google Scholar] [CrossRef]
- Chylack, L.T., Jr.; Bienfang, D.C.; Bellows, A.R.; Stillman, J.S. Ocular Manifestations of Juvenile Rheumatoid Arthritis. Am. J. Ophthalmol. 1975, 79, 1026–1033. [Google Scholar] [CrossRef]
- Paovic, J.; Paovic, P.; Vukosavljevic, M. Clinical and Immunological Features of Retinal Vasculitis in Systemic Diseases. Vojnosanit. Pregl. 2009, 66, 961–965. [Google Scholar] [CrossRef]
- Gueudry, J.; Wechsler, B.; Terrada, C.; Gendron, G.; Cassoux, N.; Fardeau, C.; Lehoang, P.; Piette, J.C.; Bodaghi, B. Long-Term Efficacy and Safety of Low-Dose Interferon alpha2a Therapy in Severe Uveitis Associated with Behcet Disease. Am. J. Ophthalmol. 2008, 146, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Shil, P.K.; Zhu, P.; Yang, H.; Verma, A.; Lei, B.; Li, Q. Angiotensin-Converting Enzyme 2 (ACE2) Activator Diminazene Aceturate Ameliorates Endotoxin-Induced Uveitis in Mice. Invest Ophthalmol. Vis. Sci. 2014, 55, 3809–3818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, U.C.; Ramana, K.V. Endotoxin-Induced Uveitis in Rodents. Methods Mol. Biol. 2013, 1031, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.; Sieper, J. Ankylosing Spondylitis. Lancet 2007, 369, 1379–1390. [Google Scholar] [CrossRef]
- Gupta, S.K.; Kalaiselvan, V.; Agrawal, S.S.; Srivastava, S.; Saxena, R. Prevention of Endotoxin-Induced Uveitis in Rabbits by Triphala, an Ayurvedic Formulation. Int. J. Cur. Biomed. Phar. Res. 2011, 1, 20–23. [Google Scholar]
- Johnson, A.C.; Heinzel, F.P.; Diaconu, E.; Sun, Y.; Hise, A.G.; Golenbock, D.; Lass, J.H.; Pearlman, E. Activation of Toll-Like Receptor (TLR) 2, TLR4, and TLR9 in the Mammalian Cornea Induces MyD88-Dependent Corneal Inflammation. Investig. Ophthalmol. Vis. Sci. 2005, 46, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Baeuerle, P.A.; Baltimore, D. NF-kappa B: Ten Years After. Cell 1996, 87, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhou, B.P. TNF-alpha/NF-kappaB/Snail Pathway in Cancer Cell Migration and Invasion. Br. J. Cancer 2010, 102, 639–644. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, J.T.; Rosenzweig, H.L.; Smith, J.R.; Martin, T.M.; Planck, S.R. Uveitis Secondary to Bacterial Products. Ophthalmic Res. 2008, 40, 165–168. [Google Scholar] [CrossRef]
- Rosenbaum, J.T.; McDevitt, H.O.; Guss, R.B.; Egbert, P.R. Endotoxin-Induced Uveitis in Rats as a Model for Human Disease. Nature 1980, 286, 611–613. [Google Scholar] [CrossRef]
- Chen, W.; Hu, X.; Zhao, L.; Li, S.; Lu, H. Expression of Toll-Like Receptor 4 in Uvea-Resident Tissue Macrophages during Endotoxin-Induced Uveitis. Mol. Vis. 2009, 15, 619–628. [Google Scholar] [PubMed]
- Girol, A.P.; Mimura, K.K.; Drewes, C.C.; Bolonheis, S.M.; Solito, E.; Farsky, S.H.; Gil, C.D.; Oliani, S.M. Anti-Inflammatory Mechanisms of the Annexin A1 Protein and Its Mimetic Peptide Ac2-26 in Models of Ocular Inflammation In Vivo and In Vitro. J. Immunol. 2013, 190, 5689–5701. [Google Scholar] [CrossRef] [PubMed]
- Gomes Bittencourt, M.; Sepah, Y.J.; Do, D.V.; Agbedia, O.; Akhtar, A.; Liu, H.; Akhlaq, A.; Annam, R.; Ibrahim, M.; Nguyen, Q.D. New Treatment Options for Noninfectious Uveitis. Dev. Ophthalmol. 2012, 51, 134–161. [Google Scholar] [CrossRef] [PubMed]
- Imrie, F.R.; Dick, A.D. Nonsteroidal Drugs for the Treatment of Noninfectious Posterior and Intermediate Uveitis. Curr. Opin. Ophthalmol. 2007, 18, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Shu-Ping, L.I.; Guo-Fu, H.U. Mechanism and Function of Angiogenin in Apoptosis Regulation. Zhongguo Sheng Wu Hua Xue Yu Fen Zi Sheng Wu Xue Bao 2015, 31, 1258–1260. [Google Scholar] [CrossRef]
- Tsuji, T.; Sun, Y.; Kishimoto, K.; Olson, K.A.; Liu, S.; Hirukawa, S.; Hu, G.F. Angiogenin is Translocated to the Nucleus of HeLa Cells and is Involved in Ribosomal RNA Transcription and Cell Proliferation. Cancer Res. 2005, 65, 1352–1360. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Xu, Z. Mechanisms of Action of Angiogenin. Acta Biochim. Biophys. Sin. (Shanghai) 2008, 40, 619–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sack, R.A.; Conradi, L.; Krumholz, D.; Beaton, A.; Sathe, S.; Morris, C. Membrane Array Characterization of 80 Chemokines, Cytokines, and Growth Factors in Open-and Closed-Eye Tears: Angiogenin and Other Defense System Constituents. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Bräuer, L.; Paulsen, F.P. Tear Film and Ocular Surface Surfactants. J. Epithel. Biol. Pharmacol. 2008, 1, 62–67. [Google Scholar] [CrossRef]
- Oikonomou, K.A.; Kapsoritakis, A.N.; Kapsoritaki, A.I.; Manolakis, A.C.; Tiaka, E.K.; Tsiopoulos, F.D.; Tsiompanidis, I.A.; Potamianos, S.P. Angiogenin, Angiopoietin-1, Angiopoietin-2, and Endostatin Serum Levels in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2011, 17, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, N.; Hatzi, E.; Bassaglia, Y.; Frendo, J.L.; Evain Brion, D.; Badet, J. Angiogenin Distribution in Human Term Placenta, and Expression by Cultured Trophoblastic Cells. Angiogenesis 2003, 6, 317–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurie, G.W.; Olsakovsky, L.A.; Conway, B.P.; McKown, R.L.; Kitagawa, K.; Nichols, J.J. Dry Eye and Designer Ophthalmics. Optom. Vis. Sci. 2008, 85, 643–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Kim, K.W.; Min, K.M.; Kim, K.W.; Chang, S.I.; Kim, J.C. Angiogenin Reduces Immune Inflammation via Inhibition of TANK-Binding Kinase 1 Expression in Human Corneal Fibroblast Cells. Mediat. Inflamm. 2014, 2014, 861435. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, K.W.; Joo, K.; Kim, J.C. Angiogenin Ameliorates Corneal Opacity and Neovascularization via Regulating Immune Response in Corneal Fibroblasts. BMC Ophthalmol. 2016, 16, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.R.; Hart, P.H.; Williams, K.A. Basic Pathogenic Mechanisms Operating in Experimental Models of Acute Anterior Uveitis. Immunol. Cell Biol. 1998, 76, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Marie, C.; Pitton, C.; Fitting, C.; Cavaillon, J.M. Regulation by Anti-Inflammatory Cytokines (IL-4, IL-10, IL-13, TGFbeta) of Interleukin-8 Production by LPS-and/ or TNFalpha-Activated Human Polymorphonuclear Cells. Mediators Inflamm. 1996, 5, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.W.; Hong, Y.S.; Chun, C.M.; Moon, J.D.; Kim, S.J.; Jung, I.C.; Yoon, Y.H.; Lee, B.A.; Moon, S.W.; Choi, S.H.; et al. Anti-Inflammatory Effects of IL-4 and IL-10 on Human Polymorphonuclear Leukocytes. J. Korean Med. Sci. 2002, 17, 7–14. [Google Scholar] [CrossRef] [Green Version]
- de Waal Malefyt, R.; Haanen, J.; Spits, H.; Roncarolo, M.G.; te Velde, A.; Figdor, C.; Johnson, K.; Kastelein, R.; Yssel, H.; de Vries, J.E. Interleukin 10 (IL-10) and viral IL-10 Strongly Reduce Antigen-Specific Human T Cell Proliferation by Diminishing the Antigen-Presenting Capacity of Monocytes via Downregulation of Class II Major Histocompatibility Complex Expression. J. Exp. Med. 1991, 174, 915–924. [Google Scholar] [CrossRef]
- Li, S.; Lu, H.; Hu, X.; Chen, W.; Xu, Y.; Wang, J. Expression of TLR4-MyD88 and NF-kappaB in the Iris during Endotoxin-Induced Uveitis. Mediat. Inflamm. 2010, 2010, 748218. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lu, H.; Hu, X.; Chen, W.; Xu, Z.; Li, S.; Xu, Y. Nuclear Factor Translocation and Acute Anterior Uveitis. Mol. Vis. 2011, 17, 170. [Google Scholar]
- Hale, S.; Lightman, S. Anti-TNF Therapies in the Management of Acute and Chronic Uveitis. Cytokine 2006, 33, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Hu, G.F. Emerging role of Angiogenin in Stress Response and Cell Survival under Adverse Conditions. J. Cell Physiol. 2012, 227, 2822–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, J.; Xu, Z. Three Decades of Research on Angiogenin: A Review and Perspective. Acta Biochim. Biophys. Sin. (Shanghai) 2016, 48, 399–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, J.; Huang, M.; Tian, Q.; Shui, Q.; Zhou, Y.; Chen, J. Downregulation of Angiogenin Inhibits the Growth and Induces Apoptosis in Human Bladder Cancer Cells through Regulating AKT/mTOR Signaling Pathway. J. Mol. Histol. 2015, 46, 157–171. [Google Scholar] [CrossRef]
- Li, S.; Ibaragi, S.; Hu, G.F. Angiogenin as a Molecular Target for the Treatment of Prostate Cancer. Curr. Cancer Ther. Rev. 2011, 7, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, N.; Wang, L.; Kishimoto, K.; Tsuji, T.; Hu, G.F. A Therapeutic Target for Prostate Cancer Based on Angiogenin-Stimulated Angiogenesis and Cancer Cell Proliferation. Proc. Natl. Acad. Sci. USA 2006, 103, 14519–14524. [Google Scholar] [CrossRef] [Green Version]
- Opitz, B.; Schroder, N.W.; Spreitzer, I.; Michelsen, K.S.; Kirschning, C.J.; Hallatschek, W.; Zahringer, U.; Hartung, T.; Gobel, U.B.; Schumann, R.R. Toll-Like Receptor-2 Mediates Treponema Glycolipid and Lipoteichoic Acid-Induced NF-kappaB Translocation. J. Biol. Chem. 2001, 276, 22041–22047. [Google Scholar] [CrossRef] [Green Version]
- Takeda, K.; Akira, S. Toll-Like Receptors. Curr. Protoc. Immunol. 2015, 109. [Google Scholar] [CrossRef]
- Su, S.B.; Silver, P.B.; Grajewski, R.S.; Agarwal, R.K.; Tang, J.; Chan, C.C.; Caspi, R.R. Essential Role of the MyD88 Pathway, but Nonessential Roles of TLRs 2, 4, and 9, in the Adjuvant Effect Promoting Th1-Mediated Autoimmunity. J. Immunol. 2005, 175, 6303–6310. [Google Scholar] [CrossRef] [Green Version]
- Vogel, S.N.; Fenton, M. Toll-Like Receptor 4 Signalling: New Perspectives on a Complex Signal-Transduction Problem. Biochem. Soc. Trans. 2003, 31, 664–668. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.J.; Gardner, P.J.; Copland, D.A.; Liyanage, S.E.; Gonzalez-Cordero, A.; Kleine Holthaus, S.M.; Luhmann, U.F.; Smith, A.J.; Ali, R.R.; Dick, A.D. Multimodal Analysis of Ocular Inflammation Using the Endotoxin-Induced Uveitis Mouse Model. Dis. Model. Mech. 2016, 9, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, K.J.; Hwang, E.; Min, K.M.; Jee, J.G.; Lee, C.K.; Hwang, K.Y.; Jeon, Y.H.; Chang, S.I.; Cheong, H.K. The Dual Binding Site of Angiogenin and Its Inhibition Mechanism: The Crystal Structure of the Rat Angiogenin-Heparin Complex. Chem. Commun. (Camb.) 2014, 50, 12966–12969. [Google Scholar] [CrossRef] [PubMed]
- Hatzi, E.; Bassaglia, Y.; Badet, J. Internalization and Processing of Human Angiogenin by Cultured Aortic Smooth Muscle celLs. Biochem. Biophys. Res. Commun. 2000, 267, 719–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viatour, P.; Merville, M.P.; Bours, V.; Chariot, A. Phosphorylation of NF-kappaB and IkappaB Proteins: Implications in Cancer and Inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef]
- Niederberger, E.; Geisslinger, G. Proteomics and NF-kappaB: An Update. Expert Rev. Proteomics 2013, 10, 189–204. [Google Scholar] [CrossRef]
- Zhong, H.; SuYang, H.; Erdjument-Bromage, H.; Tempst, P.; Ghosh, S. The Transcriptional Activity of NF-kappaB is Regulated by the IkappaB-Associated PKAc Subunit through a Cyclic AMP-Independent Mechanism. Cell 1997, 89, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Bohuslav, J.; Chen, L.F.; Kwon, H.; Mu, Y.; Greene, W.C. p53 Induces NF-kappaB Activation by an IkappaB Kinase-Independent Mechanism Involving Phosphorylation of p65 by Ribosomal S6 Kinase 1. J. Biol. Chem. 2004, 279, 26115–26125. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB Signaling in Inflammation. Signal. Transduct. Target. Ther. 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Wang, D. Discrepancy between mRNA and Protein Abundance: Insight from Information Retrieval Process in Computers. Comput. Biol. Chem. 2008, 32, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Maier, T.; Guell, M.; Serrano, L. Correlation of mRNA and Protein in Complex Biological Samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [Green Version]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between Differentially Expressed mRNA and mRNA-Protein Correlations in a Xenograft Model System. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srisa-Art, M.; Kang, D.K.; Hong, J.; Park, H.; Leatherbarrow, R.J.; Edel, J.B.; Chang, S.I.; deMello, A.J. Analysis of Protein-Protein Interactions by Using Droplet-Based Microfluidics. ChemBioChem 2009, 10, 1605–1611. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Footpad Injection | Eye Drop | Treatment Group | Number of Animals |
|---|---|---|---|
| Saline | BSS | Cont | 5 |
| LPS | BSS | BSS | 5 |
| LPS | Mutant angiogenin | mANG | 5 |
| LPS | Wild angiogenin | ANG | 5 |
| Gene | Forward Sequence (5′-3’) | Reverse Sequence (5’-3’) | Product Size (bp) |
|---|---|---|---|
| IL-1β | CACCTTCTTTTCCTTCATCTTTG | GTCGTTGCTTGTCTCTCCTTGTA | 241 |
| IL-4 | GTCACTGACTGTAGAGAGCTATTG | CTGTCGTTACATCCGTGGATAC | 107 |
| IL-8 | AGACAGTGGCAGGGATTCAC | GAGTGTGGCTATGACTTCGGT | 95 |
| IL-10 | CTGCTATGTTGCCTGCTCTTA | GGGAAGTGGGTGCAGTTATT | 86 |
| TNF-α | GACCCTCACACTCAGATCATCTTC | TTGTCTTTGAGATCCATGCCATT | 147 |
| iNOS | TTGGGTCTTGTTAGCCTAGTC | TGTGCAGTCCCAGTGAGGAAC | 262 |
| TLR4 | GGCATCATCTTCATTGTCCTTG | AGCATTGTCCTCCCACTCG | 111 |
| Myd88 | AGAGTGGAGAGCAGTGTC | GGCAGTAGCAGATGAAGG | 109 |
| GAPDH | GCAAGGATACTGAGAGCAAG | GGATGGAATTGTGAGGGAGA | 98 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.; Kim, J.T.; Lee, S.J.; Kim, J.C. The Anti-Inflammatory Effects of Angiogenin in an Endotoxin Induced Uveitis in Rats. Int. J. Mol. Sci. 2020, 21, 413. https://doi.org/10.3390/ijms21020413
Park J, Kim JT, Lee SJ, Kim JC. The Anti-Inflammatory Effects of Angiogenin in an Endotoxin Induced Uveitis in Rats. International Journal of Molecular Sciences. 2020; 21(2):413. https://doi.org/10.3390/ijms21020413
Chicago/Turabian StylePark, Jihae, Jee Taek Kim, Soo Jin Lee, and Jae Chan Kim. 2020. "The Anti-Inflammatory Effects of Angiogenin in an Endotoxin Induced Uveitis in Rats" International Journal of Molecular Sciences 21, no. 2: 413. https://doi.org/10.3390/ijms21020413
APA StylePark, J., Kim, J. T., Lee, S. J., & Kim, J. C. (2020). The Anti-Inflammatory Effects of Angiogenin in an Endotoxin Induced Uveitis in Rats. International Journal of Molecular Sciences, 21(2), 413. https://doi.org/10.3390/ijms21020413