Epimers of Vitamin D: A Review



{kind=link}
{kind=link}
{kind=link}
Abstract
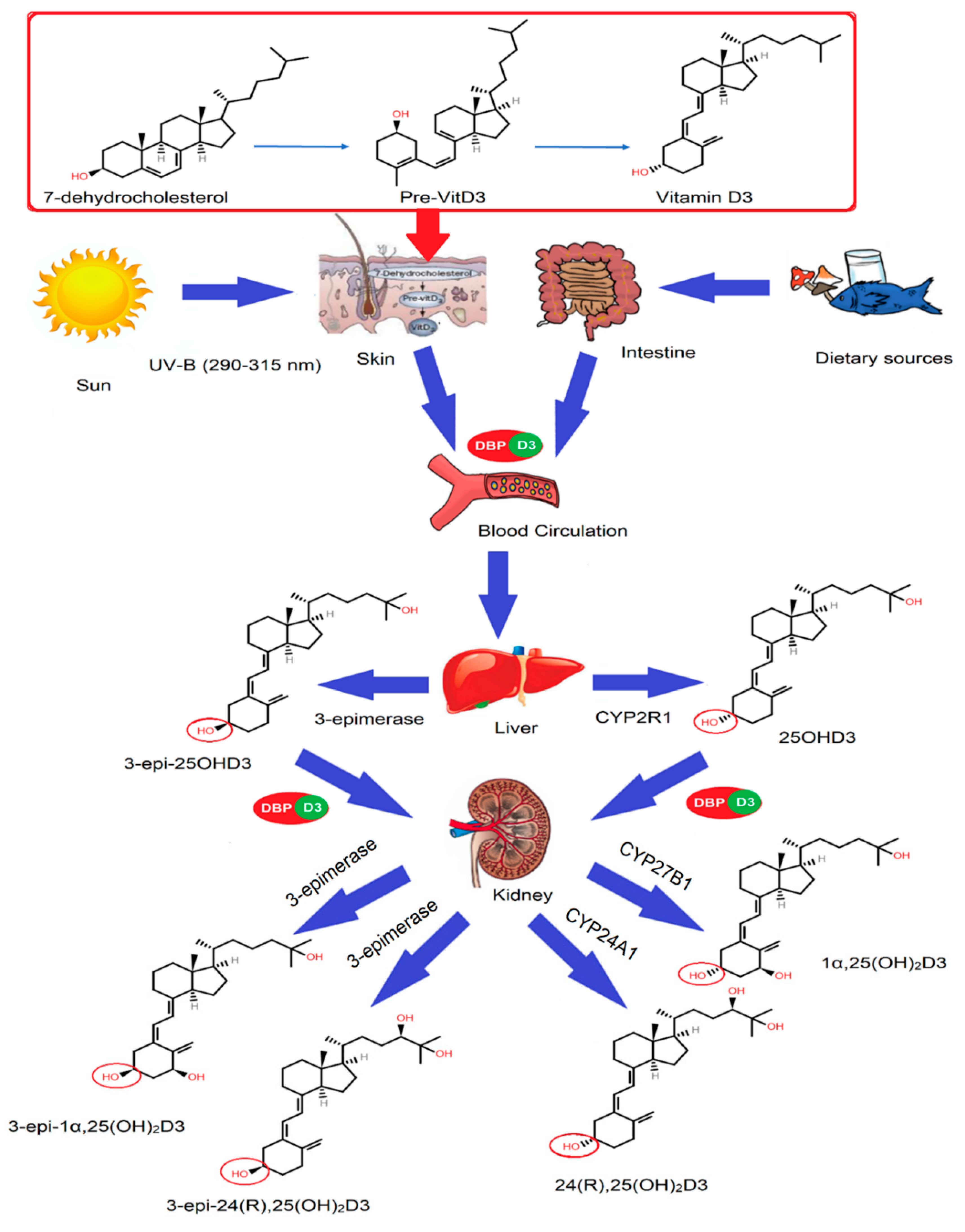
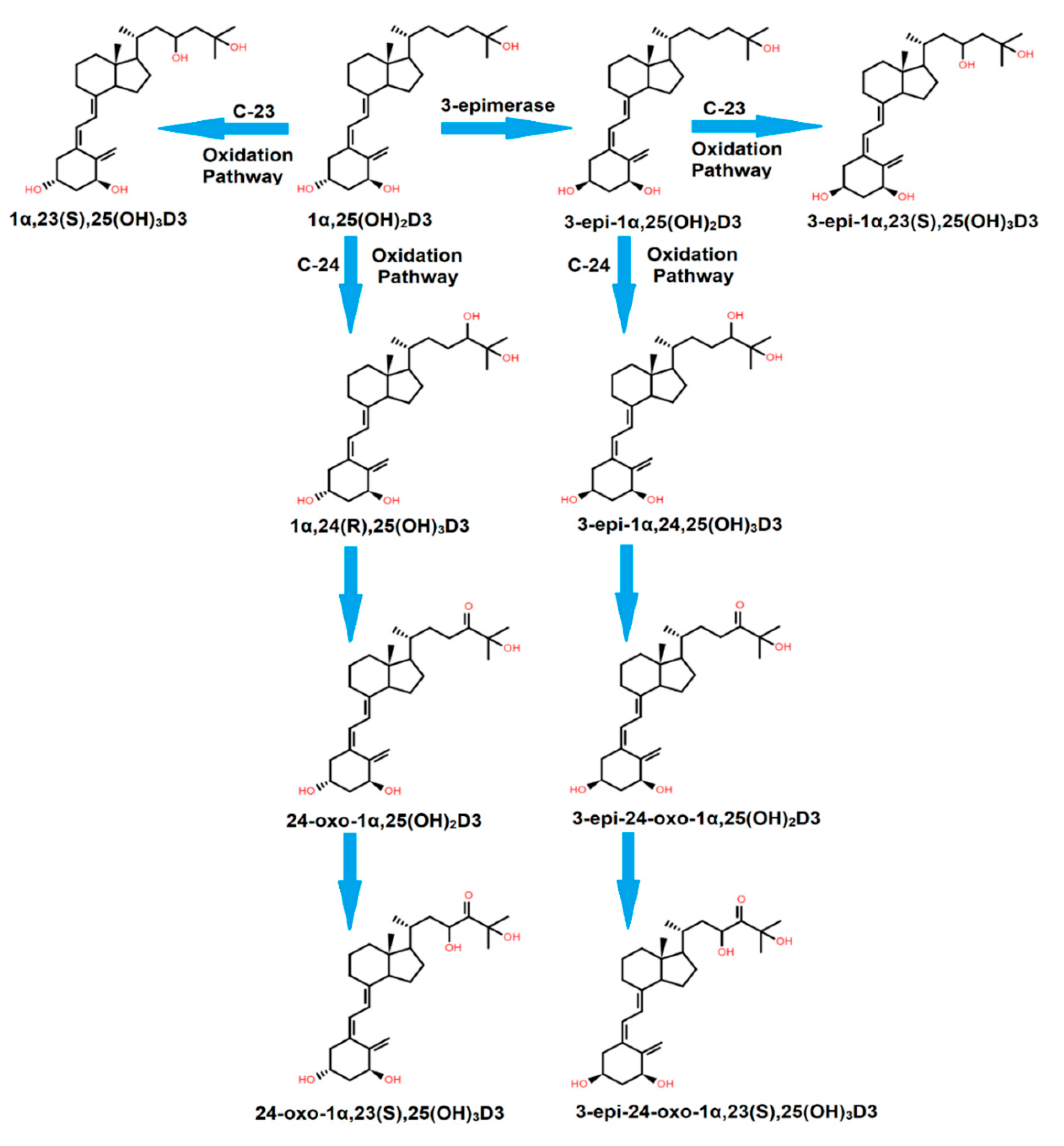
:- All major vitamin D forms can undergo epimerization at the C3 position, leading to the formation of C3-epimers that can overestimate vitamin D status in routine laboratory tests.
- Higher levels of C3-epimers are observed in mothers and newborns.
- Oral supplementation of vitamin D can cause an increased production of epimers in mice.
- LC-MS/MS can be used to separate C3-epimers from other vitamin D metabolites.
1. Introduction
2. C3-Epimerization of Vitamin D
3. Role of Epimers in Calcium, Phosphorus and PTH Homeostasis
4. Potency of Epimerization in Microsomal Fractions
5. The Role of DBP, VDR, and Genetics in Epimerization
6. Vitamin D Epimer Levels in Humans and Mice
7. C-3 Epimer Levels in Newborn and Adults
8. Techniques for Measurement of Vitamin D Epimers
9. Quality Assurance in C-3 Epimers Determination
10. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Ghaly, S.; Bliuc, D.; Center, J.R.; Clarke, M.W.; Jones, A.P.; Trend, S.; Kermode, A.G.; Neale, R.E.; Hart, P.H. Vitamin D C3-epimer levels are proportionally higher with oral vitamin D supplementation compared to ultraviolet irradiation of skin in mice but not humans. J. Steroid Biochem. Mol. Biol. 2019, 186, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.J.; Taylor, R.L.; Reddy, G.S.; Grebe, S.K.G. C-3 epimers can account for a significant proportion of total circulating 25-hydroxyvitamin D in infants, complicating accurate measurement and interpretation of vitamin D status. J. Clin. Endocrinol. Metab. 2006, 91, 3055–3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mydtskov, N.D.; Lykkedegn, S.; Fruekilde, P.B.N.; Nielsen, J.; Barington, T.; Christesen, H.T. S-25-hydroxyvitamin D and C3-epimers in pregnancy and infancy: An Odense Child Cohort study. Clin. Biochem. 2017, 50, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Torugsa, S.; Nimitphong, H.; Warodomwichit, D.; Chailurkit, L.; Srijaruskul, K.; Chanprasertyothin, S.; Ongphiphadhanakul, B. The genetic determinants of circulating C3-epimers of 25-hydroxyvitamin D. J. Clin. Transl. Endocrinol. 2018, 12, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Shah, I.; Al-Dabbagh, B.; Gariballa, S.; Al-Menhali, A.; Muhammad, N.; Yasin, J.; Ashraf, S.S. Application of a new vitamin D blood test on the Emirati population. J. Steroid Biochem. Mol. Biol. 2018, 180, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Shah, I.; James, R.; Barker, J.; Petroczi, A.; Naughton, D.P. Misleading measures in Vitamin D analysis: A novel LC-MS/MS assay to account for epimers and isobars. Nutr. J. 2011, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Shah, I.; Akhtar, M.K.; Hisaindee, S.; Rauf, M.A.; Sadig, M.; Ashraf, S.S. Clinical diagnostic tools for vitamin D assessment. J. Steroid Biochem. Mol. Biol. 2018, 180, 105–117. [Google Scholar] [CrossRef]
- Institute of Medicine (US) Committee to Review Dietary Reference Intakes for Vitamin D and Calcium. Dietary Reference Intakes for Calcium and Vitamin D; Ross, A.C., Taylor, C.L., Yaktine, A.L., Del Valle, H.B., Eds.; The National Academies Collection: Reports Funded by National Institutes of Health; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Giovannucci, E. Chapter 58—Methods of Evaluating Population Studies of Vitamin D: Strengths and Weaknesses. In Vitamin D, 4th ed.; Feldman, D., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 3–14. ISBN 978-0-12-809963-6. [Google Scholar]
- Holick, M.F.; Chen, T.C. Vitamin D deficiency: A worldwide problem with health consequences. Am. J. Clin. Nutr. 2008, 87, 1080S–1086S. [Google Scholar] [CrossRef] [Green Version]
- Clemens, T.L.; Henderson, S.L.; Adams, J.S.; Holick, M.F. Increased Skin Pigment Reduces the Capacity of Skin to Synthesise Vitamin D3. Lancet 1982, 319, 74–76. [Google Scholar] [CrossRef]
- Sedrani, S.H. Low 25-hydroxyvitamin D and normal serum calcium concentrations in Saudi Arabia: Riyadh region. Ann. Nutr. Metab. 1984, 28, 181–185. [Google Scholar] [CrossRef]
- El-Hajj Fuleihan, G.; Nabulsi, M.; Choucair, M.; Salamoun, M.; Hajj Shahine, C.; Kizirian, A.; Tannous, R. Hypovitaminosis D in healthy schoolchildren. Pediatrics 2001, 107, E53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holick, M.F.; Matsuoka, L.Y.; Wortsman, J. Age, vitamin D, and solar ultraviolet. Lancet 1989, 2, 1104–1105. [Google Scholar] [CrossRef]
- Holick, M.F. Resurrection of vitamin D deficiency and rickets. J. Clin. Investig. 2006, 116, 2062–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff-Ferrari, H.A.; Giovannucci, E.; Willett, W.C.; Dietrich, T.; Dawson-Hughes, B. Estimation of optimal serum concentrations of 25-hydroxyvitamin D for multiple health outcomes. Am. J. Clin. Nutr. 2006, 84, 18–28. [Google Scholar] [CrossRef]
- Chiu, K.C.; Chu, A.; Go, V.L.W.; Saad, M.F. Hypovitaminosis D is associated with insulin resistance and beta cell dysfunction. Am. J. Clin. Nutr. 2004, 79, 820–825. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Naughton, D.P. Vitamin D in health and disease: Current perspectives. Nutr. J. 2010, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Bailey, D.; Veljkovic, K.; Yazdanpanah, M.; Adeli, K. Analytical measurement and clinical relevance of vitamin D3 C3-epimer. Clin. Biochem. 2013, 46, 190–196. [Google Scholar] [CrossRef]
- Holick, M.F.; MacLaughlin, J.A.; Clark, M.B.; Holick, S.A.; Potts, J.T.; Anderson, R.R.; Blank, I.H.; Parrish, J.A.; Elias, P. Photosynthesis of previtamin D3 in human skin and the physiologic consequences. Science 1980, 210, 203–205. [Google Scholar] [CrossRef]
- Holick, M.F.; Tian, X.Q.; Allen, M. Evolutionary importance for the membrane enhancement of the production of vitamin D3 in the skin of poikilothermic animals. Proc. Natl. Acad. Sci. USA 1995, 92, 3124–3126. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, T.; Izumi, S.; Ide, H.; Ohyama, Y. Identification of a novel rat microsomal vitamin D3 25-hydroxylase. J. Biol. Chem. 2004, 279, 22848–22856. [Google Scholar] [CrossRef] [Green Version]
- Tuckey, R.C.; Cheng, C.Y.S.; Slominski, A.T. The serum vitamin D metabolome: What we know and what is still to discover. J. Steroid Biochem. Mol. Biol. 2019, 186, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Schuster, I. Cytochromes P450 are essential players in the vitamin D signaling system. Biochim. Biophys. Acta (BBA) Proteins Proteomics 2011, 1814, 186–199. [Google Scholar] [CrossRef]
- Reddy, G.S.; Muralidharan, K.R.; Okamura, W.H.; Tserng, K.Y.; McLane, J.A. Metabolism of 1alpha,25-dihydroxyvitamin D(3) and its C-3 epimer 1alpha,25-dihydroxy-3-epi-vitamin D(3) in neonatal human keratinocytes. Steroids 2001, 66, 441–450. [Google Scholar] [CrossRef]
- Kamao, M.; Tatematsu, S.; Hatakeyama, S.; Sakaki, T.; Sawada, N.; Inouye, K.; Ozono, K.; Kubodera, N.; Reddy, G.S.; Okano, T. C-3 epimerization of vitamin D3 metabolites and further metabolism of C-3 epimers: 25-hydroxyvitamin D3 is metabolized to 3-epi-25-hydroxyvitamin D3 and subsequently metabolized through C-1alpha or C-24 hydroxylation. J. Biol. Chem. 2004, 279, 15897–15907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischof, M.G.; Siu-Caldera, M.L.; Weiskopf, A.; Vouros, P.; Cross, H.S.; Peterlik, M.; Reddy, G.S. Differentiation-related pathways of 1 alpha,25-dihydroxycholecalciferol metabolism in human colon adenocarcinoma-derived Caco-2 cells: Production of 1 alpha,25-dihydroxy-3epi-cholecalciferol. Exp. Cell Res. 1998, 241, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Siu-Caldera, M.-L.; Sekimoto, H.; Weiskopf, A.; Vouros, P.; Muralidharan, K.R.; Okamura, W.H.; Bishop, J.; Norman, A.W.; Uskoković, M.R.; Schuster, I.; et al. Production of 1α,25-Dihydroxy-3-epi-vitamin D3 in two rat osteosarcoma cell lines (UMR 106 and ROS 17/2.8): Existence of the C-3 epimerization pathway in ROS 17/2.8 cells in which the C-24 oxidation pathway is not expressed. Bone 1999, 24, 457–463. [Google Scholar] [CrossRef]
- Brown, A.J.; Ritter, C.; Slatopolsky, E.; Muralidharan, K.R.; Okamura, W.H.; Reddy, G.S. 1Alpha,25-dihydroxy-3-epi-vitamin D3, a natural metabolite of 1alpha,25-dihydroxyvitamin D3, is a potent suppressor of parathyroid hormone secretion. J. Cell. Biochem. 1999, 73, 106–113. [Google Scholar] [CrossRef]
- Sekimoto, H.; Siu-Caldera, M.-L.; Weiskopf, A.; Vouros, P.; Muralidharan, K.R.; Okamura, W.H.; Uskokovic, M.R.; Reddy, G.S. 1α,25-Dihydroxy-3-epi-vitamin D3: In vivo metabolite of 1α,25-dihydroxyvitamin D3 in rats. FEBS Lett. 1999, 448, 278–282. [Google Scholar] [CrossRef] [Green Version]
- Kamao, M.; Hatakeyama, S.; Sakaki, T.; Sawada, N.; Inouye, K.; Kubodera, N.; Reddy, G.S.; Okano, T. Measurement and characterization of C-3 epimerization activity toward vitamin D3. Arch. Biochem. Biophys. 2005, 436, 196–205. [Google Scholar] [CrossRef]
- Kamao, M.; Tatematsu, S.; Sawada, N.; Sakaki, T.; Hatakeyama, S.; Kubodera, N.; Okano, T. Cell specificity and properties of the C-3 epimerization of Vitamin D3 metabolites. J. Steroid Biochem. Mol. Biol. 2004, 89, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Rehan, V.K.; Torday, J.S.; Peleg, S.; Gennaro, L.; Vouros, P.; Padbury, J.; Rao, D.S.; Reddy, G.S. 1Alpha,25-dihydroxy-3-epi-vitamin D3, a natural metabolite of 1alpha,25-dihydroxy vitamin D3: Production and biological activity studies in pulmonary alveolar type II cells. Mol. Genet. Metab. 2002, 76, 46–56. [Google Scholar] [CrossRef]
- Bianchini, C. The Generation of C-3α Epimer of 25-hydroxyvitamin D in Adult Sprague Dawley Rats and Older Adult Humans. Ph.D. Thesis, McGill University, Montréal, QC, Canada, April 2014. [Google Scholar]
- Fleet, J.C.; Bradley, J.; Reddy, G.S.; Ray, R.; Wood, R.J. 1 alpha,25-(OH)2-vitamin D3 analogs with minimal in vivo calcemic activity can stimulate significant transepithelial calcium transport and mRNA expression in vitro. Arch. Biochem. Biophys. 1996, 329, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Kadiyala, S.; Nagaba, S.; Takeuchi, K.; Yukihiro, S.; Qiu, W.; Eyes, S.T.; Uskokovic, M.R.; Posner, G.H.; Reddy, G.S.; Guggino, S.E. Metabolites and analogs of 1alpha,25-dihydroxyvitamin D(3): Evaluation of actions in bone. Steroids 2001, 66, 347–355. [Google Scholar] [CrossRef]
- Astecker, N.; Reddy, G.S.; Herzig, G.; Vorisek, G.; Schuster, I. 1alpha,25-Dihydroxy-3-epi-vitamin D3 a physiological metabolite of 1alpha,25-dihydroxyvitamin D3: Its production and metabolism in primary human keratinocytes. Mol. Cell. Endocrinol. 2000, 170, 91–101. [Google Scholar] [CrossRef]
- Rhieu, S.Y.; Annalora, A.J.; Wang, G.; Flarakos, C.C.; Gathungu, R.M.; Vouros, P.; Sigüeiro, R.; Mouriño, A.; Schuster, I.; Palmore, G.T.R.; et al. Metabolic stability of 3-epi-1α,25-dihydroxyvitamin D3 over 1 α 25-dihydroxyvitamin D3: Metabolism and molecular docking studies using rat CYP24A1. J. Cell. Biochem. 2013, 114, 2293–2305. [Google Scholar] [CrossRef]
- Kusudo, T.; Sakaki, T.; Abe, D.; Fujishima, T.; Kittaka, A.; Takayama, H.; Hatakeyama, S.; Ohta, M.; Inouye, K. Metabolism of A-ring diastereomers of 1alpha,25-dihydroxyvitamin D3 by CYP24A1. Biochem. Biophys. Res. Commun. 2004, 321, 774–782. [Google Scholar] [CrossRef]
- Phinney, K.W.; Bedner, M.; Tai, S.S.-C.; Vamathevan, V.V.; Sander, L.C.; Sharpless, K.E.; Wise, S.A.; Yen, J.H.; Schleicher, R.L.; Chaudhary-Webb, M.; et al. Development and certification of a standard reference material for vitamin D metabolites in human serum. Anal. Chem. 2012, 84, 956–962. [Google Scholar] [CrossRef] [Green Version]
- Yetley, E.A.; Pfeiffer, C.M.; Schleicher, R.L.; Phinney, K.W.; Lacher, D.A.; Christakos, S.; Eckfeldt, J.H.; Fleet, J.C.; Howard, G.; Hoofnagle, A.N.; et al. NHANES monitoring of serum 25-hydroxyvitamin D: A roundtable summary. J. Nutr. 2010, 140, 2030S–2045S. [Google Scholar]
- Higashi, T.; Sakajiri, K.; Shimada, K. Analysis of C-3 epimerization in (24R)-24,25-dihydroxyvitamin D3 catalyzed by hydroxysteroid dehydrogenase. J. Pharm Biomed. Anal. 2004, 36, 429–436. [Google Scholar] [CrossRef]
- Penning, T.M.; Sharp, R.B.; Krieger, N.R. Purification and properties of 3 alpha-hydroxysteroid dehydrogenase from rat brain cytosol. Inhibition by nonsteroidal anti-inflammatory drugs and progestins. J. Biol. Chem. 1985, 260, 15266–15272. [Google Scholar]
- Hara, A.; Inoue, Y.; Nakagawa, M.; Naganeo, F.; Sawada, H. Purification and characterization of NADP+-dependent 3 alpha-hydroxysteroid dehydrogenase from mouse liver cytosol. J. Biochem. 1988, 103, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.C.; Kochakian, C.D. The induction, purification and characterization of 17 beta-hydroxy-C19-steroid dehydrogenase of the female guinea pig kidney. J. Steroid Biochem. 1979, 10, 187–193. [Google Scholar] [CrossRef]
- Bikle, D. Vitamin D: Production, Metabolism, and Mechanisms of Action. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., et al., Eds.; MDText.com, Inc.: Dartmouth, MA, USA, 2000. [Google Scholar]
- Mandarino, N.R.; Júnior, F.d.C.M.; Salgado, J.V.L.; Lages, J.S.; Salgado Filho, N. Is Vitamin D Deficiency a New Risk Factor for Cardiovascular Disease? Open Cardiovasc. Med. J. 2015, 9, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, F.; Sigüeiro, R.; Sato, Y.; Araujo, C.; Schuster, I.; Antony, P.; Peluso, J.; Muller, C.; Mouriño, A.; Moras, D.; et al. 1α,25(OH)2-3-epi-vitamin D3, a natural physiological metabolite of vitamin D3: Its synthesis, biological activity and crystal structure with its receptor. PLoS ONE 2011, 6, e18124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ooms, N.; van Daal, H.; Beijers, A.M.; Gerrits, G.P.J.M.; Semmekrot, B.A.; van den Ouweland, J.M.W. Time-course analysis of 3-epi-25-hydroxyvitamin D3 shows markedly elevated levels in early life, particularly from vitamin D supplementation in preterm infants. Pediatr. Res. 2016, 79, 647–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdanpanah, M.; Bailey, D.; Walsh, W.; Wan, B.; Adeli, K. Analytical measurement of serum 25-OH-vitamin D₃, 25-OH-vitamin D₂ and their C3-epimers by LC-MS/MS in infant and pediatric specimens. Clin. Biochem. 2013, 46, 1264–1271. [Google Scholar] [CrossRef]
- Teegarden, M.D.; Campbell, A.R.; Cooperstone, J.L.; Tober, K.L.; Schwartz, S.J.; Oberyszyn, T.M. 25-Hydroxyvitamin D3 and its C-3 epimer are elevated in the skin and serum of Skh-1 mice supplemented with dietary vitamin D3. Mol. Nutr. Food Res. 2017, 61, 1700293. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.-K.; Li, W.; Postlethwaite, A.; Tieu, E.W.; Tang, E.K.Y.; Tuckey, R.C. Detection of novel CYP11A1-derived secosteroids in the human epidermis and serum and pig adrenal gland. Sci. Rep. 2015, 5, 14875. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.T.; Li, W.; Kim, T.-K.; Semak, I.; Wang, J.; Zjawiony, J.K.; Tuckey, R.C. Novel activities of CYP11A1 and their potential physiological significance. J. Steroid Biochem. Mol. Biol. 2015, 151, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.T.; Kim, T.-K.; Shehabi, H.Z.; Semak, I.; Tang, E.K.Y.; Nguyen, M.N.; Benson, H.A.E.; Korik, E.; Janjetovic, Z.; Chen, J.; et al. In vivo evidence for a novel pathway of vitamin D3 metabolism initiated by P450scc and modified by CYP27B1. FASEB J. 2012, 26, 3901–3915. [Google Scholar] [CrossRef] [Green Version]
- Luca, M.D.; D’Anna, F.; Bondanza, S.; Franzi, A.T.; Cancedda, R. Human epithelial cells induce human melanocyte growth in vitro but only skin keratinocytes regulate its proper differentiation in the absence of dermis. J. Cell Biol. 1988, 107, 1919–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabierowski, S.E.; Fukunaga-Kalabis, M.; Li, L.; Herlyn, M. Dermis-derived stem cells: A source of epidermal melanocytes and melanoma? Pigment Cell Melanoma Res. 2011, 24, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV Light Touches the Brain and Endocrine System Through Skin, and Why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Ouweland, J.M.W.; Beijers, A.M.; van Daal, H. Overestimation of 25-hydroxyvitamin D3 by increased ionisation efficiency of 3-epi-25-hydroxyvitamin D3 in LC–MS/MS methods not separating both metabolites as determined by an LC–MS/MS method for separate quantification of 25-hydroxyvitamin D3, 3-epi-25-hydroxyvitamin D3 and 25-hydroxyvitamin D2 in human serum. J. Chromatogr. B 2014, 967, 195–202. [Google Scholar]
- Aghajafari, F.; Field, C.J.; Rabi, D.; Kaplan, B.J.; Maggiore, J.A.; O’Beirne, M.; Hanley, D.A.; Eliasziw, M.; Dewey, D.; Ross, S. Plasma 3-Epi-25-Hydroxycholecalciferol Can Alter the Assessment of Vitamin D Status Using the Current Reference Ranges for Pregnant Women and Their Newborns. J. Nutr. 2016, 146, 70–75. [Google Scholar]
- Park, H.; Wood, M.R.; Malysheva, O.V.; Jones, S.; Mehta, S.; Brannon, P.M.; Caudill, M.A. Placental vitamin D metabolism and its associations with circulating vitamin D metabolites in pregnant women. Am. J. Clin. Nutr. 2017, 106, 1439–1448. [Google Scholar] [CrossRef] [Green Version]
- Baecher, S.; Leinenbach, A.; Wright, J.A.; Pongratz, S.; Kobold, U.; Thiele, R. Simultaneous quantification of four vitamin D metabolites in human serum using high performance liquid chromatography tandem mass spectrometry for vitamin D profiling. Clin. Biochem. 2012, 45, 1491–1496. [Google Scholar] [CrossRef]
- Keevil, B. Does the presence of 3-epi-25OHD3 affect the routine measurement of vitamin D using liquid chromatography tandem mass spectrometry? Clin. Chem. Lab. Med. 2011, 50, 181–183. [Google Scholar] [CrossRef]
- Chailurkit, L.; Aekplakorn, W.; Ongphiphadhanakul, B. Serum C3 epimer of 25-hydroxyvitamin D and its determinants in adults: A national health examination survey in Thais. Osteoporos. Int. 2015, 26, 2339–2344. [Google Scholar] [CrossRef]
- Strathmann, F.G.; Sadilkova, K.; Laha, T.J.; LeSourd, S.E.; Bornhorst, J.A.; Hoofnagle, A.N.; Jack, R. 3-epi-25 hydroxyvitamin D concentrations are not correlated with age in a cohort of infants and adults. Clin. Chim. Acta 2012, 413, 203–206. [Google Scholar] [CrossRef] [Green Version]
- Van den Ouweland, J.M.W.; Beijers, A.M.; van Daal, H. Fast separation of 25-hydroxyvitamin D3 from 3-epi-25-hydroxyvitamin D3 in human serum by liquid chromatography-tandem mass spectrometry: Variable prevalence of 3-epi-25-hydroxyvitamin D3 in infants, children, and adults. Clin. Chem. 2011, 57, 1618–1619. [Google Scholar] [CrossRef] [Green Version]
- Schleicher, R.L.; Encisco, S.E.; Chaudhary-Webb, M.; Paliakov, E.; McCoy, L.F.; Pfeiffer, C.M. Isotope dilution ultra performance liquid chromatography-tandem mass spectrometry method for simultaneous measurement of 25-hydroxyvitamin D2, 25-hydroxyvitamin D3 and 3-epi-25-hydroxyvitamin D3 in human serum. Clin. Chim. Acta 2011, 412, 1594–1599. [Google Scholar] [CrossRef]
- Jenkinson, C.; Taylor, A.; Storbeck, K.-H.; Hewison, M. Analysis of multiple vitamin D metabolites by ultra-performance supercritical fluid chromatography-tandem mass spectrometry (UPSFC-MS/MS). J. Chromatogr. B 2018, 1087, 43–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rola, R.; Kowalski, K.; Bieńkowski, T.; Kołodyńska-Goworek, A.; Studzińska, S. Development of a method for multiple vitamin D metabolite measurements by liquid chromatography coupled with tandem mass spectrometry in dried blood spots. Analyst 2019, 144, 299–309. [Google Scholar] [CrossRef]
- Hollis, B.W. Editorial: The determination of circulating 25-hydroxyvitamin D: No easy task. J. Clin. Endocrinol. Metab. 2004, 89, 3149–3151. [Google Scholar] [CrossRef] [Green Version]
- Horst, R.L.; Reinhardt, T.A.; Beitz, D.C.; Littledike, E.T. A sensitive competitive protein binding assay for vitamin D in plasma. Steroids 1981, 37, 581–591. [Google Scholar] [CrossRef]
- Jones, G. Assay of vitamins D2 and D3, and 25-hydroxyvitamins D2 and D3 in human plasma by high-performance liquid chromatography. Clin. Chem. 1978, 24, 287–298. [Google Scholar]
- Terry, A.H.; Sandrock, T.; Meikle, A.W. Measurement of 25-hydroxyvitamin D by the Nichols ADVANTAGE, DiaSorin LIAISON, DiaSorin RIA, and liquid chromatography-tandem mass spectrometry. Clin. Chem. 2005, 51, 1565–1566. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Schoenmakers, I.; Jones, K.; Koulman, A.; Prentice, A.; Volmer, D.A. Quantitative determination of vitamin D metabolites in plasma using UHPLC-MS/MS. Anal. Bioanal. Chem. 2010, 398, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Lind, C.; Chen, J.; Byrjalsen, I. Enzyme immunoassay for measuring 25-hydroxyvitamin D3 in serum. Clin. Chem. 1997, 43, 943–949. [Google Scholar]
- Lensmeyer, G.L.; Wiebe, D.A.; Binkley, N.; Drezner, M.K. HPLC method for 25-hydroxyvitamin D measurement: Comparison with contemporary assays. Clin. Chem. 2006, 52, 1120–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Rogers, K.; Wardle, R.; El-Khoury, J.M. High-throughput measurement of 25-hydroxyvitamin D by LC–MS/MS with separation of the C3-epimer interference for pediatric populations. Clin. Chim. Acta 2016, 454, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Gallo, S.; Comeau, K.; Agellon, S.; Vanstone, C.; Sharma, A.; Jones, G.; L’abbé, M.; Khamessan, A.; Weiler, H.; Rodd, C. Methodological issues in assessing plasma 25-hydroxyvitamin D concentration in newborn infants. Bone 2014, 61, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.D.; Jones, J.C.; Shannon, J.; Williams, E.L.; Jones, G.; Kaufmann, M.; Sempos, C. 25-Hydroxyvitamin D assays: Potential interference from other circulating vitamin D metabolites. J. Steroid Biochem. Mol. Biol. 2016, 164, 134–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couchman, L.; Benton, C.M.; Moniz, C.F. Variability in the analysis of 25-hydroxyvitamin D by liquid chromatography-tandem mass spectrometry: The devil is in the detail. Clin. Chim. Acta 2012, 413, 1239–1243. [Google Scholar] [CrossRef]
- Hanson, C.; Anderson-Berry, A.; Lyden, E.; Kaufmann, M.; Wu, A.; Elliott, E.; Lee, J.-I.; Jones, G. Dynamics of Vitamin D Metabolism in Maternal-Fetal Dyads. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 486–490. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Zohily, B.; Al-Menhali, A.; Gariballa, S.; Haq, A.; Shah, I. Epimers of Vitamin D: A Review. Int. J. Mol. Sci. 2020, 21, 470. https://doi.org/10.3390/ijms21020470
Al-Zohily B, Al-Menhali A, Gariballa S, Haq A, Shah I. Epimers of Vitamin D: A Review. International Journal of Molecular Sciences. 2020; 21(2):470. https://doi.org/10.3390/ijms21020470
Chicago/Turabian StyleAl-Zohily, Bashar, Asma Al-Menhali, Salah Gariballa, Afrozul Haq, and Iltaf Shah. 2020. "Epimers of Vitamin D: A Review" International Journal of Molecular Sciences 21, no. 2: 470. https://doi.org/10.3390/ijms21020470
APA StyleAl-Zohily, B., Al-Menhali, A., Gariballa, S., Haq, A., & Shah, I. (2020). Epimers of Vitamin D: A Review. International Journal of Molecular Sciences, 21(2), 470. https://doi.org/10.3390/ijms21020470