Small RNA and mRNA Sequencing Reveal the Roles of microRNAs Involved in Pomegranate Female Sterility


Abstract
:1. Introduction
2. Results
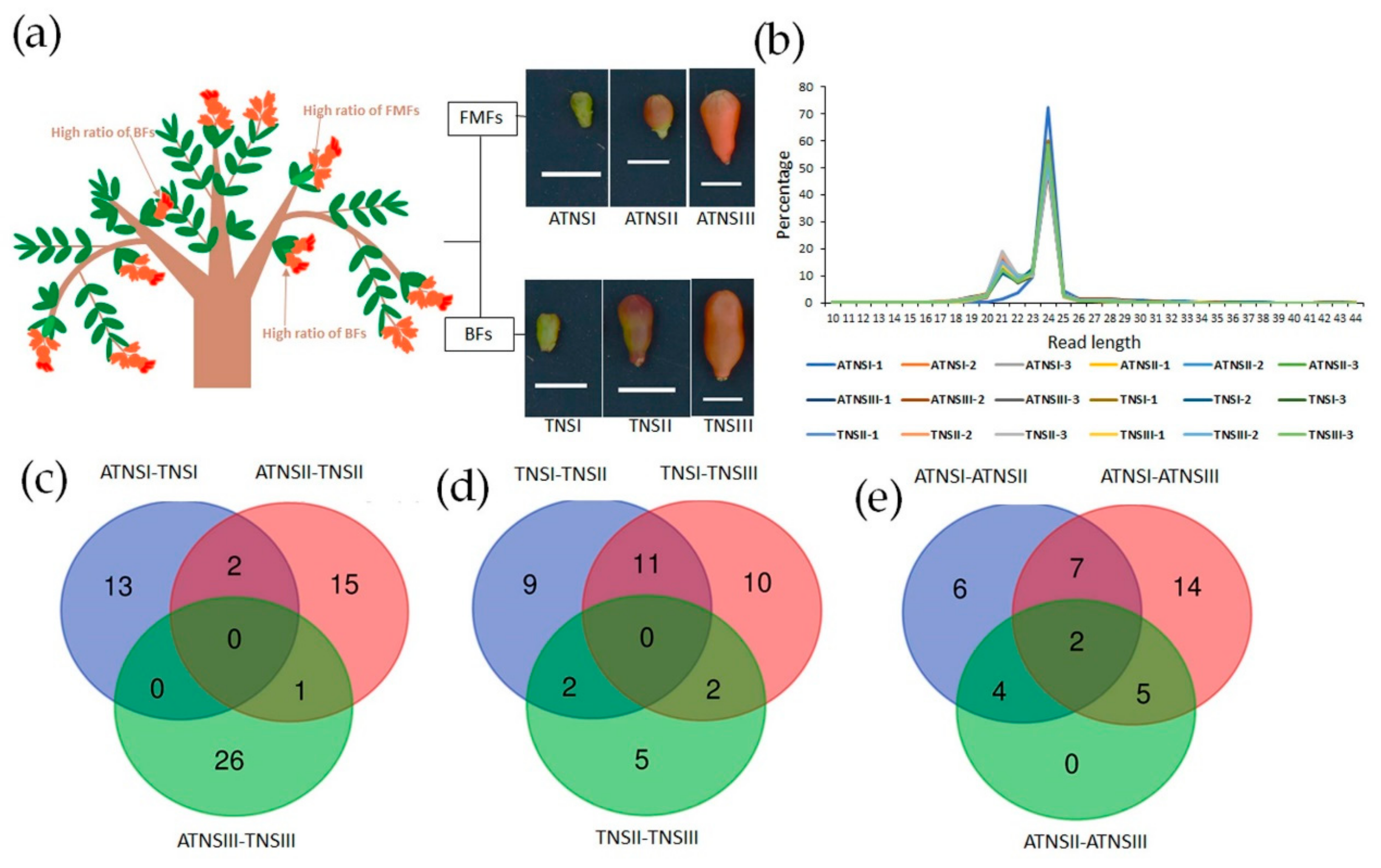
2.1. Deep Sequencing of Small RNAs of Pomegranate Fertile and Sterile Pistils
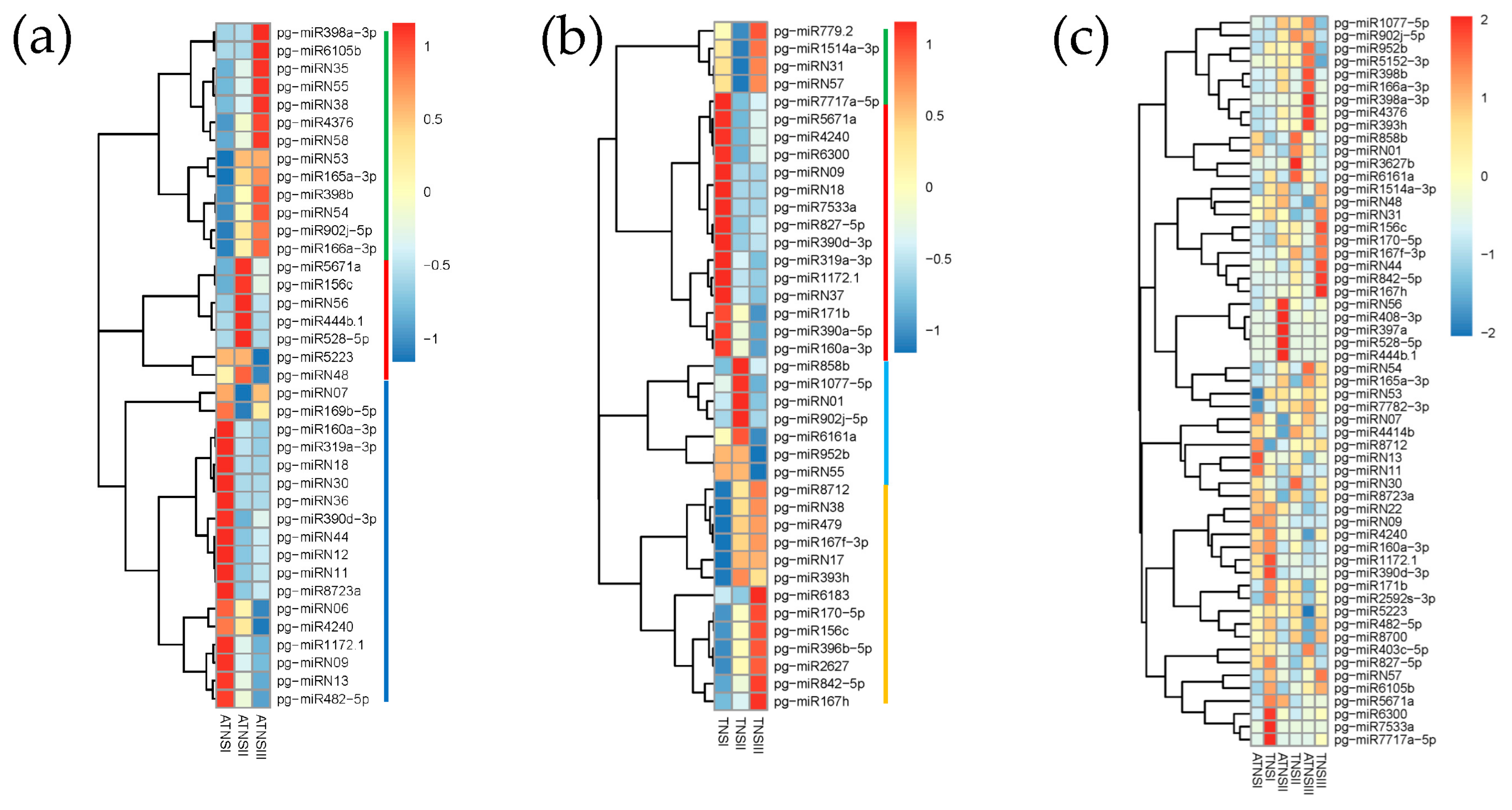
2.2. Identifying miRNAs Involved in Pomegranate Female Sterility
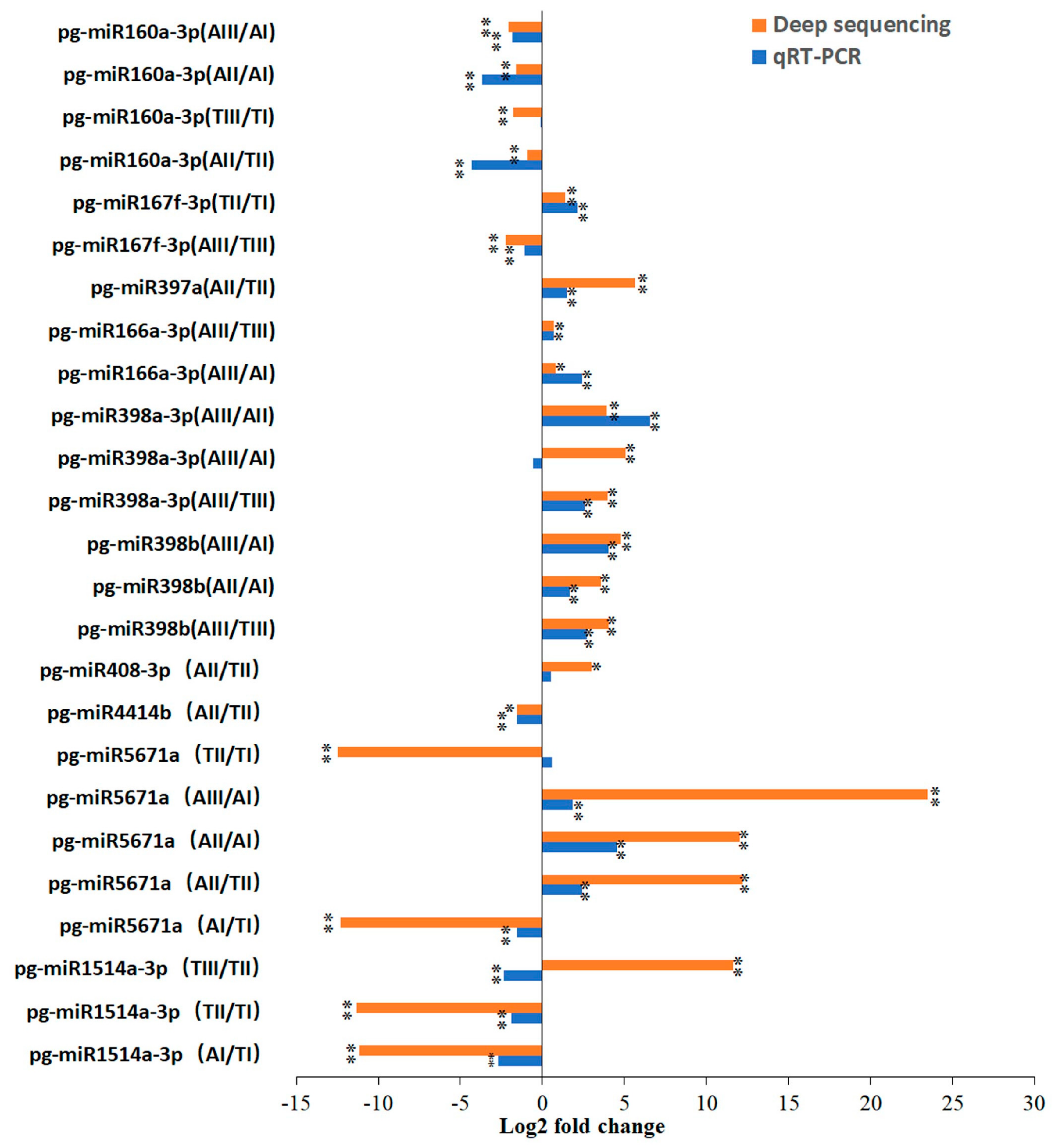
2.3. Validation of Expression Patterns of miRNAs
2.4. Identification and Analysis of miRNA Targets
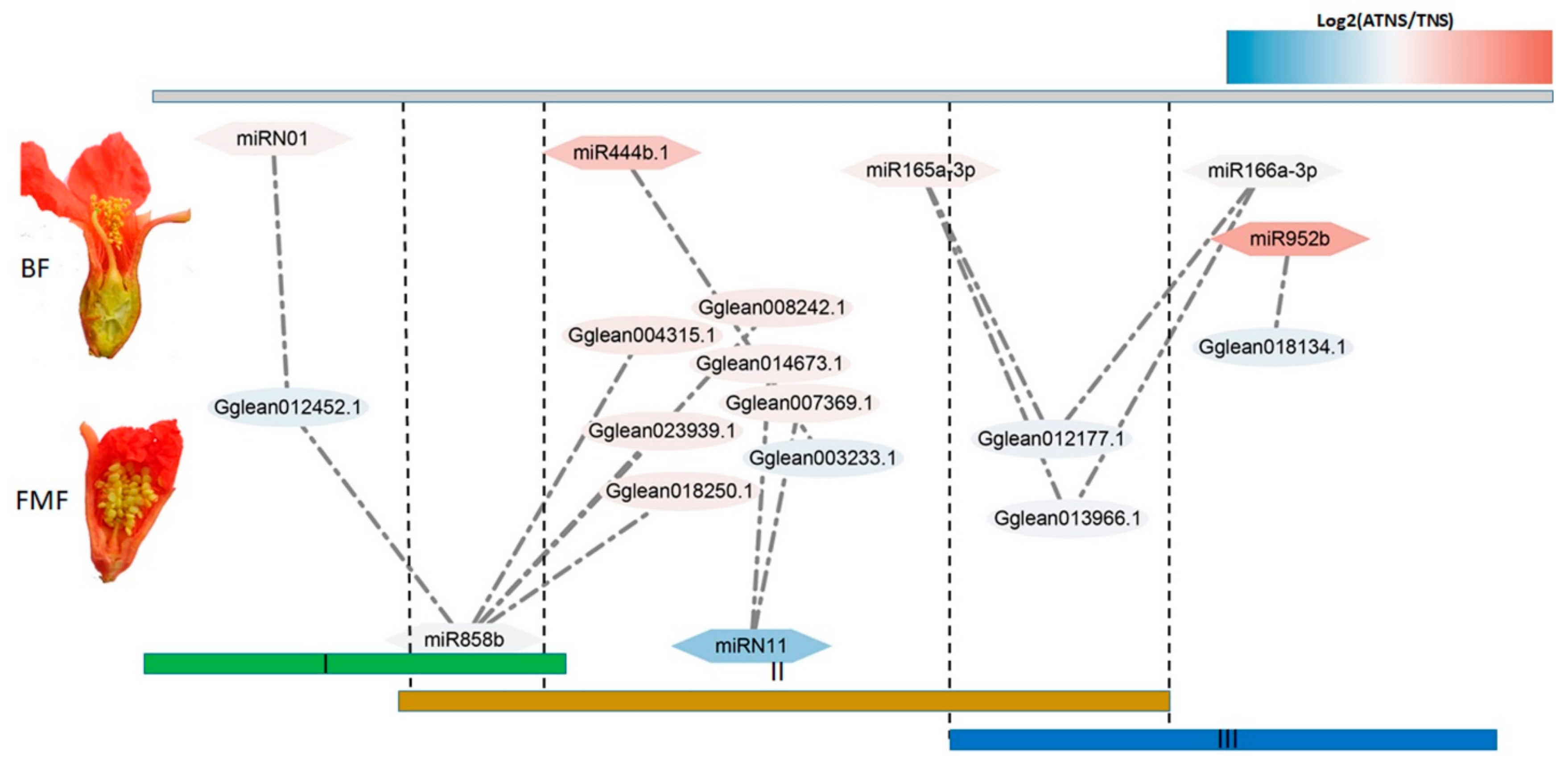
2.5. MicroRNA–mRNA Interaction Identification
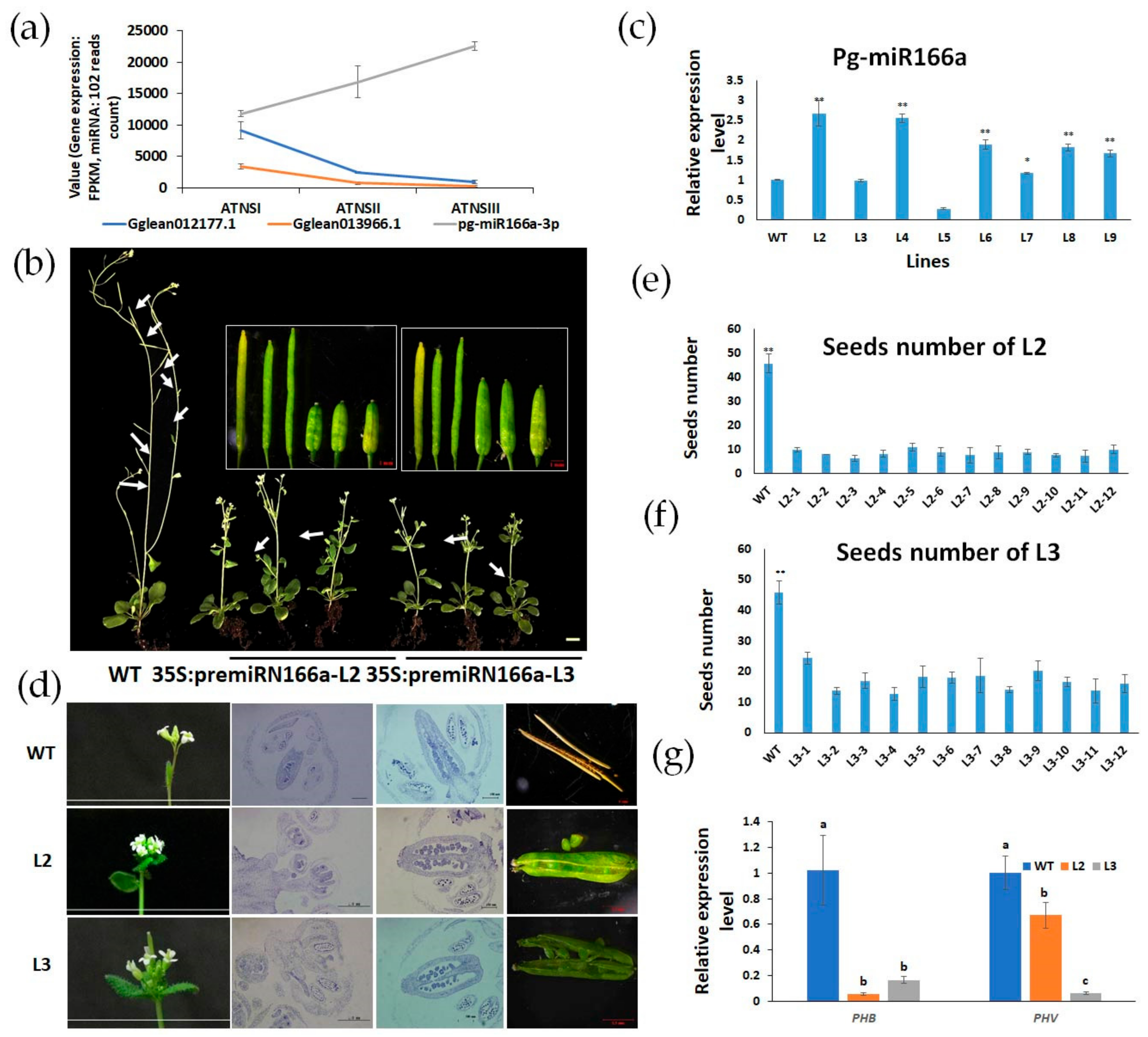
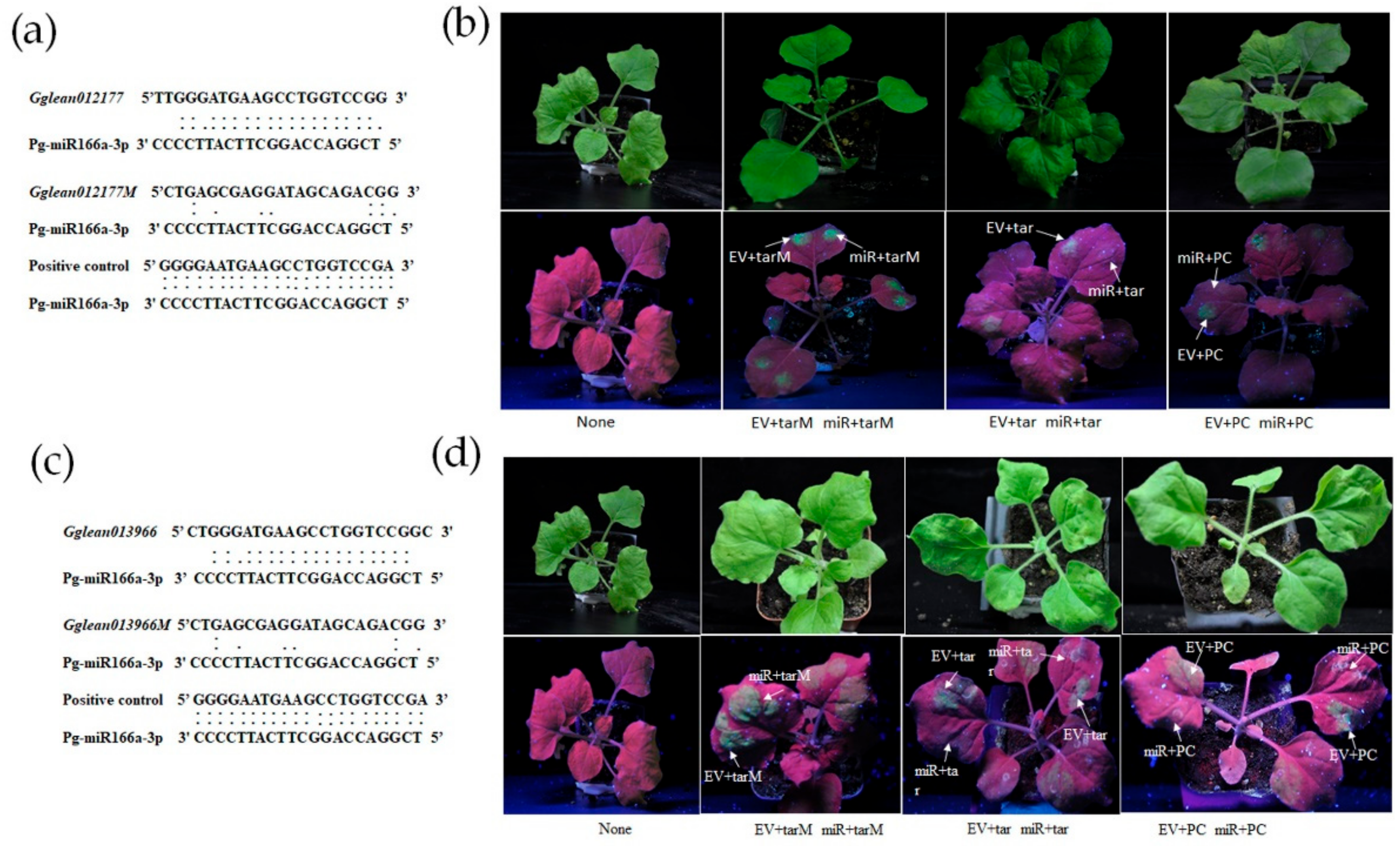
2.6. Pg-miR166a-3p Functions via Target HD-Zip Transcription Factor
2.7. Newly Identified miRNAs in Pomegranate
3. Discussion
3.1. Potential Roles of miRNAs in Pomegranate Female Sterility
3.2. MicroRNA Targets Regulating Reproduction Development Involved in Female Sterility
4. Materials and Methods
4.1. Sample Collection
4.2. RNA Isolation, Library Construction, and Sequencing
4.3. Bioinformatic Analysis
4.4. MicroRNA Expression Profiles and Comparison between FMFs and BFs
4.5. MicroRNA-target Prediction and Association Analysis with Transcription Analysis Result
4.6. MicroRNA Function Annotation
4.7. Validation of miRNA Expression in Pomegranate Female Pistils by Stem-Loop qRT-PCR
4.8. Nicotiana Benthamiana Transient Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pereira, T.N.S.; Lersten, N.R.; Palmer, R.G. Genetic and cytological analyses of a partial-female-sterile mutant (PS1) in soybean (Glycine max; Leguminosae). Am. J. Bot. 1997, 84, 781–791. [Google Scholar] [CrossRef]
- Tang, J.J.; Chen, X.; Hu, Q.D.; Kato, M.; Shimizu, K.; Yokoo, M. A comparatively histological observation on the mega gametophytic abortion of female-sterile rice FS-1 and its maternal parent Fujisaka 5. Acta Biol. Exp. Sinica. 2002, 35, 313–318. [Google Scholar] [PubMed]
- Chen, X.J.; Qi, C.K.; Zhang, J.F.; Pu, H.M.; Gao, J.Q.; Fu, S.Z. A primary study on biological characteristics of female sterile mutant FS-M1 of rapeseed (Brassica napus L.). Chin. J. Oil Crop Sci. 2003, 25, 12–15. [Google Scholar]
- Chakrabarty, S. Female sterility and carpel development in plants. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Champaign, IL, USA, 2017. Available online: http://hdl.handle.net/2142/98088 (accessed on 18 December 2019).
- Aviram, M.; Rosenblat, M.; Gaitini, D.; Samy, N.; Aaron, H.; Leslie, D.; Nina, V.; Dita, P.; Judith, A.; Harley, L.; et al. Pomegranate juice consumption for 3 years by patients with carotid artery stenosis reduces common carotid intima-media thickness, blood pressure and LDL oxidation. Clin. Nutr. 2004, 23, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Pooja, S.; Sarah, M.C.; Farrukh, A. Pomegranate for prevention and treatment of cancer: An update. Molecules 2017, 22, 177. [Google Scholar] [CrossRef] [Green Version]
- Holland, D.; Hatib, K.; Bar-Yaakov, I. Pomegranate: Botany, horticulture, breeding. Hort. Rev. 2009, 35, 127–191. [Google Scholar]
- Wetzstein, H.Y.; Ravid, N.; Wilkins, E.; Martinelli, A.P. A morphological and histological characterization of bisexual and male flower types in pomegranate. J. Am. Soc. Horti. Sci. 2011, 136, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.N.; Li, H.X.; Niu, J.; Xue, H.; Liu, B.B.; Wang, Q.; Zhang, F.H.; Zhao, D.G.; Cao, S.Y. Growth characteristics and morphology differences between bisexual flowers and functionally male flowers of ‘Tunisiruanzi’ pomegranate. J. Fruit Sci. 2017, 34, 80–87. [Google Scholar]
- Chen, L.N.; Zhang, J.; Li, H.X.; Niu, J.; Xue, H.; Liu, B.B.; Wang, Q.; Luo, X.; Zhang, F.H.; Zhao, D.G.; et al. Transcriptomic analysis reveals candidate genes for female sterility in pomegranate flowers. Front. Plant Sci. 2017, 8, 1430. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Jones, D.C.; Wang, Q.L.; Sun, R.R.; Zhang, B.H. Small RNA sequencing identifies miRNA roles in ovule and fibre development. Plant Biotechnol. J. 2015, 13, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X. A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development. Science 2004, 303, 2022–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, L.; Dunoyer, P.; Jay, F.; Arnold, B.; Dharmasiri, N.; Estelle, M.; Voinnet, O.; Jones, J.D.G. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science 2006, 312, 436–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunkar, R.; Zhu, J.K. Novel and stress-regulated micro-RNAs and other small RNAs from Arabidopsis. Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.W.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.M.; Xue, W.; Dong, C.J.; Jin, L.G.; Bian, S.M.; Wang, C.; Wu, X.Y.; Liu, J.Y. A comparative miRNAome analysis reveals seven fiber initiation-related and 36 novel miRNAs in developing cotton ovules. Mol. Plant 2012, 5, 889–900. [Google Scholar] [CrossRef] [Green Version]
- Reiser, L.; Fischer, R.L. The ovule and the embryo sac. Plant Cell. 1993, 5, 1291–1301. [Google Scholar] [CrossRef]
- Elliott, R.C.; Betzner, A.S.; Huttner, E.; Oakes, M.P.; Tucker, W.Q.; Gerentes, D.; Perez, P.; Smyth, D.R. Aintegumenta, an apetala2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth. Plant Cell 1996, 8, 155–168. [Google Scholar]
- Jin, Y.G. Botany, 2nd ed.; Science Press: Beijing, China, 2010. [Google Scholar]
- Wu, Y.F.; Reed, G.W.; Tian, C.Q. Arabidopsis microRNA167 controls patterns of ARF6 and ARF8 expression, and regulates both female and male reproduction. Development 2006, 133, 4211–4218. [Google Scholar] [CrossRef] [Green Version]
- Sieber, P.; Petrascheck, M.; Barberis, A.; Schneitz, K. Organ polarity in Arabidopsis. NOZZLE physically interacts with members of the YABBY family. Plant Physiol. 2004, 135, 2172–2185. [Google Scholar] [PubMed] [Green Version]
- Hashimoto, K.; Miyashima, S.; Sato-Nara, K.; Yamada, T.; Nakajima, K. Functionally diversified members of the MIR165/6 gene family regulate ovule morphogenesis in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 1017–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.F.; Ning, K.; Che, G.; Yan, S.S.; Han, L.J.; Gu, R.; Li, Z.; Weng, Y.Q.; Zhang, X.L. CsSPL functions as an adaptor between HD-ZIP III and CsWUS transcription factors regulating anther and ovule development in cucumber. Plant J. 2018, 94, 535–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Tu, L.L.; Wang, L.C.; Hu, H.Y.; Xu, J.; Zhang, X.L. microRNA 157-targeted SPL genes regulate floral organ size and ovule production in cotton. BMC Plant Biol. 2017, 17, 7. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.F.F.; Silva, E.M.; Mariana, D.S.A.; Azevedo, M.S.; Guivin, M.A.; Ramiro, D.A.; Figueiredo, C.R.; Carrer, R.; Peres, L.E.P.; Nogueira, F.T.S. microRNA156-targeted SPL/SBP box transcription factors regulate tomato ovary and fruit development. Plant J. 2014, 78, 604–618. [Google Scholar] [CrossRef]
- Kanupriya, C.; Radhika, V.; Ravishankar, K.V. Mining of miRNAs in pomegranate (Punica granatum L.) by pyrosequencing of part of the genome. J. Pomol. Horti. Sci. 2013, 88, 735–742. [Google Scholar]
- Saminathan, T.; Bodunrin, A.; Singh, N.V.; Devarajan, R.; Nimmakayala, P.; Jeff, M.; Aeadhya, M.; Reddy, U.K. Genome-wide identification of microRNAs in pomegranate (Punica granatum L.) by high-throughput sequencing. BMC Plant Biol. 2016, 16, 122. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Cao, D.; Zhang, J.F.; Chen, L.; Xia, X.C.; Li, H.X.; Zhao, D.G.; Zhang, F.H.; Xue, H.; Chen, L.N.; et al. Integrated microRNA and mRNA expression profiling reveals a complex network regulating pomegranate (Punica granatum L.) seed hardness. Sci. Rep. 2018, 8, 9292. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Li, H.X.; Wu, Z.K.; Yao, W.; Zhao, P.; Cao, D.; Yu, H.Y.; Li, K.D.; Poudel, K.; Zhao, D.G.; et al. The pomegranate (Punica granatum L.) draft genome dissects genetic divergence between soft- and hard-seeded cultivars. Plant Biotechnol. J. 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Yamada, T.; Sasaki, Y.; Hashimoto, K.; Nakajima, K.; Gasser, C.S. CORONA, PHABULOSA and PHAVOLUTA collaborate with BELL1 to confine WUSCHEL expression to the nucellus in Arabidopsis ovules. Development 2016, 143, 422–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Yang, L.; Yu, M.; Wang, J.B. Identification and expression analysis of microRNAs during ovule development in rice (Oryza sativa) by deep sequencing. Plant Cell Rep. 2017, 36, 1815–1827. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Pang, M.; Nah, G.; Shi, X.L.; Ye, W.X.; Stelly, D.M.; Chen, Z.J. miR828 and miR858 regulate homoeologous MYB2 gene functions in Arabidopsis trichome and cotton fibre development. Nat. Commun. 2014, 5, 3050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramona, S.; Susanne, D.; Cornelia, G.; Gerd, H.; Benedikt, A.; Tom, S.; Sylvestre, M.; Yoshihiro, O.; Hiroshi, E.; Ivan, F.; et al. Tomato MYB21 acts in ovules to mediate jasmonate-regulated fertility. Plant Cell 2019, 31, 1043–1062. [Google Scholar]
- Luo, M.; Dennis, E.S.; Berger, F.; Peacock, W.J.; Chaudhury, A. MINISEED3 (MINI3), a WRKY family gene, and HAIKU2 (IKU2), a leucine-rich repeat (LRR) KINASE gene, are regulators of seed size in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 17531–17536. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Palma, J.D.; Oane, R.; Gamuyao, R.; Luo, M.; Chaudhury, A.; Herve, P.; Xue, Q.Z.; Bennett, J. OsTDL1A binds to the LRR domain of rice receptor kinase MSP1, and is required to limit sporocyst numbers. Plant J. 2010, 54, 375–387. [Google Scholar] [CrossRef] [Green Version]
- Li, X.F.; Li, Y.J.; An, Y.H.; Xiong, L.J.; Shao, X.U.; Wang, Y.; Sun, Y. AKINβ1 is involved in the regulation of nitrogen metabolism and sugar signaling in Arabidopsis. J. Integr. Plant Biol. 2009, 51, 513–520. [Google Scholar] [CrossRef]
- Sawicki, M.; Jacquens, L.; Baillieul, F.; Clement, C.; Vaillant-Gaveau, N.; Jacquard, C. Distinct regulation in inflorescence carbohydrate metabolism according to grapevine cultivars during floral development. Physiol. Plantarum. 2015, 154, 447–467. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jung, J.H.; Reyes, J.L.; Kim, Y.S.; Kim, S.Y.; Chung, K.S.; Kim, J.; Lee, M.; Lee, Y.; Kim, V.N.; et al. microRNA-directed cleavage of ATHB15 mRNA regulates vascular development in Arabidopsis inflorescence stems. Plant J. 2005, 42, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Chae, K.; Lord, E.M. Pollen tube growth and guidance: Roles of small, secreted proteins. Ann. Bot. 2011, 108, 627–636. [Google Scholar] [CrossRef] [Green Version]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J.L. Radial patterning of Arabidopsis shoots by class III HD-ZIP and KANADI genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef] [Green Version]
- McConnell, J.R.; Barton, K. Leaf polarity and meristem formation in Arabidopsis. Development 1998, 125, 2935–2942. [Google Scholar] [PubMed]
- Kelley, D.R.; Skinner, D.J.; Gasser, C.S. Roles of polarity determinants in ovule development. Plant J. 2009, 57, 1054–1064. [Google Scholar] [CrossRef] [Green Version]
- Kiefer, E.; Heller, W.; Ernst, D. A simple and efficient protocol for isolation of functional RNA from plant tissues rich in secondary metabolites. Plant Mol. Biol. Rep. 2000, 18, 33–39. [Google Scholar] [CrossRef]
- Pearson, W.R.; Wood, T.; Zhang, Z.; Miller, W. Comparison of DNA sequences with protein sequences. Genomics 1997, 46, 24–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package ‘corrplot’. Statistician 2017, 56, 316–324. [Google Scholar]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Meth. 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedlander, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.W.; Ryu, M.Y.; Viczián, A.; Jung, H.J.; Kim, G.M.; Arce, A.L.; Achkar, N.P.; Manavella, P.; Dolde, U.; Wenkel, S. Light triggers the miRNA-biogenetic inconsistency for de-etiolated seedling survivability in Arabidopsis thaliana. Mol. Plant 2019. [Google Scholar] [CrossRef]
- Perdikari, A.; Leparc, G.G.; Balaz, M.; Pires, N.D.; Lidell, M.E.; Sun, W.; Fernandez-Albert, F.; Müller, S.; Akchiche, N.; Dong, H.; et al. BATLAS: Deconvoluting Brown Adipose Tissue. Cell Rep. 2018, 25, 784–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaselli, S.; Galeano, F.; Alon, S.; Raho, S.; Galardi, S.; Polito, V.A.; Presutti, C.; Vincenti, S.; Eisenberg, E.; Locatelli, F.; et al. Modulation of microRNA editing, expression and processing by ADAR2 deaminase in glioblastoma. Genome Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Bo, X.; Wang, S. TargetFinder: A software for antisense oligonucleotide target site selection based on MAST and secondary structures of target mRNA. Bioinformatics 2005, 21, 1401–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.J.; Ma, Y.K.; Chen, T.; Wang, M.; Wang, X.J. PsRobot: A web-based plant small RNA meta-analysis toolbox. Nucleic Acids Res. 2012, 40, W22–W28. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Grocock, R.J.; Dongen, S.V.; Bateman, A.; Enright, A.J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.Y.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2016, 1, 2019–2025. [Google Scholar] [CrossRef]
- Liu, Y.L.; Wang, L.; Chen, D.J.; Wu, X.M.; Huang, D.; Chen, L.L.; Li, L.; Deng, X.X.; Xu, Q. Genome-wide comparison of microRNAs and their targeted transcripts among leaf, flower and fruit of sweet orange. BMC Genom. 2014, 15, 695. [Google Scholar] [CrossRef] [Green Version]
- Long, J.M.; Liu, Z.; Wu, X.M.; Fang, Y.N.; Jia, H.H.; Xie, Z.Z.; Deng, X.X.; Guo, W.W. Genome-scale mRNA and small RNA transcriptomic insights into initiation of citrus apomixis. J. Exp. Bot. 2016, 67, 5743–5756. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA ID | p-Value | Log2 (ATNS/TNS) | Regulation | Target Gene ID | p-Value | Log2 (ATNS/TNS) | Regulation |
|---|---|---|---|---|---|---|---|
| ATNSI/TNSI | |||||||
| miR858b | 0.0005004 | 0.921592594 | UP | Gglean012452.1 | 0.0052191 | −2.680132231 | DOWN |
| miRN01 | 0.0000002 | 1.432745067 | UP | Gglean012452.1 | 0.0052191 | −2.680132231 | DOWN |
| ATNSII/TNSII | |||||||
| miRN11 | 0.0093121 | −13.15241036 | DOWN | Gglean007369.1 | 0.0136615 | 1.678181538 | UP |
| miRN11 | 0.0093121 | −13.15241036 | DOWN | Gglean014673.1 | 0.0043412 | 1.779818526 | UP |
| miR165a-3p | 0.0150903 | 1.861219672 | UP | Gglean013966.1 | 0.0000209 | −1.188541887 | DOWN |
| miR165a-3p | 0.0150903 | 1.861219672 | UP | Gglean012177.1 | 0.0001133 | −1.099146097 | DOWN |
| miR858b | 0.0005004 | −0.863522677 | DOWN | Gglean008242.1 | 0.000037 | 2.754373731 | UP |
| miR858b | 0.0005004 | −0.863522677 | DOWN | Gglean018250.1 | 0.000000251 | 2.372360824 | UP |
| miR858b | 0.0005004 | −0.863522677 | DOWN | Gglean023939.1 | 0.0020669 | 2.157568169 | UP |
| miR858b | 0.0005004 | −0.863522677 | DOWN | Gglean004315.1 | 0.00000026 | 2.769310866 | UP |
| miR444b.1 | 0.0482444 | 8.687029035 | UP | Gglean003233.1 | 0.0001883 | −2.440374727 | DOWN |
| ATNSIII/TNSIII | |||||||
| miR952b | 0.000000000000000000000509 | 14.53397128 | UP | Gglean018134.1 | 0.0004515 | −2.379136194 | DOWN |
| miR166a-3p | 0.00000000000000000491 | 0.718106582 | UP | Gglean012177.1 | 0.0000015 | −1.614832343 | DOWN |
| miR166a-3p | 0.00000000000000000491 | 0.718106582 | UP | Gglean013966.1 | 0.0077827 | −1.215622716 | DOWN |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Luo, X.; Yang, X.; Jing, D.; Xia, X.; Li, H.; Poudel, K.; Cao, S. Small RNA and mRNA Sequencing Reveal the Roles of microRNAs Involved in Pomegranate Female Sterility. Int. J. Mol. Sci. 2020, 21, 558. https://doi.org/10.3390/ijms21020558
Chen L, Luo X, Yang X, Jing D, Xia X, Li H, Poudel K, Cao S. Small RNA and mRNA Sequencing Reveal the Roles of microRNAs Involved in Pomegranate Female Sterility. International Journal of Molecular Sciences. 2020; 21(2):558. https://doi.org/10.3390/ijms21020558
Chicago/Turabian StyleChen, Lina, Xiang Luo, Xuanwen Yang, Dan Jing, Xiaocong Xia, Haoxian Li, Krishna Poudel, and Shangyin Cao. 2020. "Small RNA and mRNA Sequencing Reveal the Roles of microRNAs Involved in Pomegranate Female Sterility" International Journal of Molecular Sciences 21, no. 2: 558. https://doi.org/10.3390/ijms21020558
APA StyleChen, L., Luo, X., Yang, X., Jing, D., Xia, X., Li, H., Poudel, K., & Cao, S. (2020). Small RNA and mRNA Sequencing Reveal the Roles of microRNAs Involved in Pomegranate Female Sterility. International Journal of Molecular Sciences, 21(2), 558. https://doi.org/10.3390/ijms21020558