The Impact of IPTG Induction on Plasmid Stability and Heterologous Protein Expression by Escherichia coli Biofilms


Abstract
:1. Introduction
2. Results
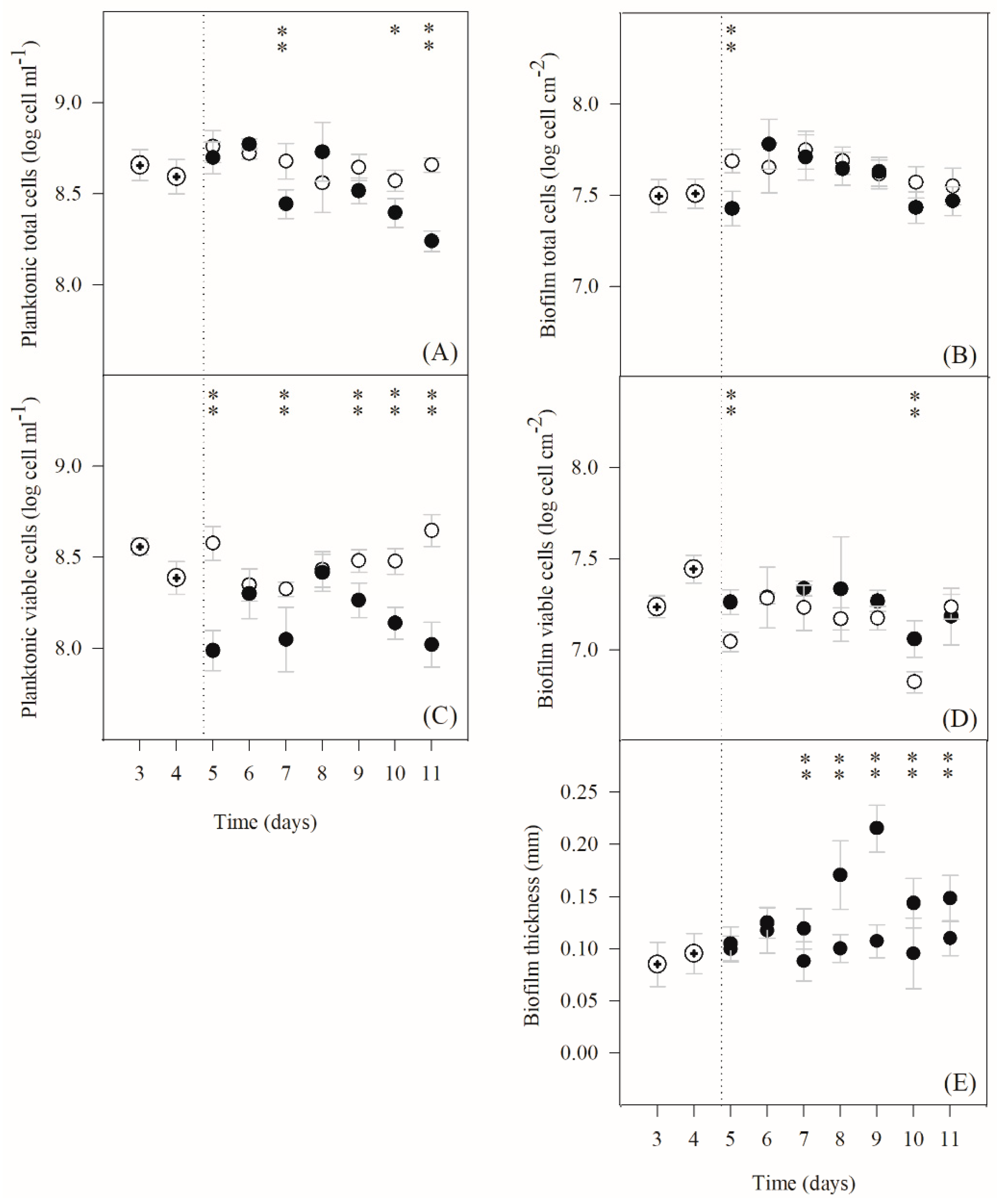
2.1. Effect of IPTG Induction on Planktonic and Biofilm Growth
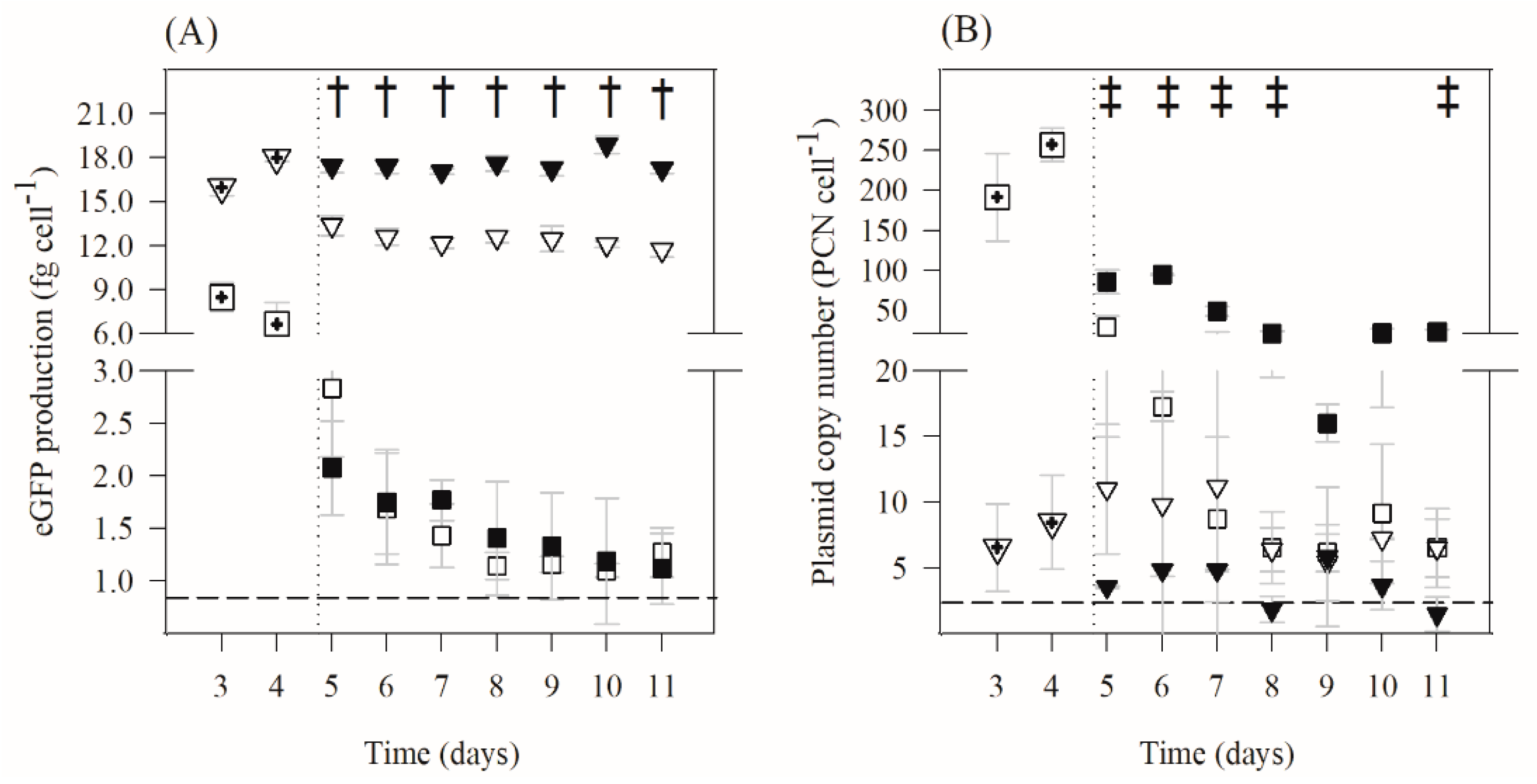
2.2. Determination of eGFP, PCN, mRNA and Total RNA
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Culture Medium
4.2. Biofilm Formation System
4.3. Estimation of IPTG Transfer Rate
4.4. Biofilm and Planktonic Analysis
4.5. Quantification of EPS
4.6. Quantification of eGFP Production
4.7. Quantification of PCN
4.7.1. Preparation of Plasmid DNA (pDNA) Standards
4.7.2. Real-Time PCR (RT-PCR)
4.8. Quantification of Total RNA
4.9. Quantification of eGFP Gene Transcription
4.10. Statistical Analysis
 ) and 95% (p values < 0.05;
) and 95% (p values < 0.05;  or
or  ). Paired t-test analysis (p values < 0.05) was also performed using Statgraphics software to compare biofilm and planktonic curves in Figure 2 (statistically significant differences between biofilm conditions were marked with †, while differences between planktonic conditions were indicated with ‡).
). Paired t-test analysis (p values < 0.05) was also performed using Statgraphics software to compare biofilm and planktonic curves in Figure 2 (statistically significant differences between biofilm conditions were marked with †, while differences between planktonic conditions were indicated with ‡).5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Area of the coupon (L2) | |
| Bulk IPTG concentration (M L−3) | |
| Hydraulic diameter (L) | |
| Dilution rate (T−1) | |
| Diffusion coefficient of IPTG in water (L2 T−1) | |
| Volumetric flow rate (L3 T−1) | |
| External mass transfer coefficient (L T−1) | |
| Mass transfer rate (M T−1) | |
| Reynolds number (-) | |
| Schmidt number (-) | |
| Sherwood number (-) | |
| Time (T) | |
| Total system volume (L3) | |
| Average residence time (T) |
References
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [Green Version]
- Mergulhão, F.J.; Summers, D.K.; Monteiro, G.A. Recombinant protein secretion in Escherichia coli. Biotechnol. Adv. 2005, 23, 177–202. [Google Scholar] [CrossRef] [PubMed]
- Mergulhão, F.J.; Monteiro, G.A. Secretion capacity limitations of the Sec pathway in Escherichia coli. J. Microbiol. Biotechnol. 2004, 14, 128–133. [Google Scholar]
- Sanchez-Garcia, L.; Martín, L.; Mangues, R.; Ferrer-Miralles, N.; Vázquez, E.; Villaverde, A. Recombinant pharmaceuticals from microbial cells: A 2015 update. Microb. Cell Fact. 2016, 15, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mergulhão, F.J.; Monteiro, G.A.; Cabral, J.M.S.; Taipa, M.A. Design of bacterial vector systems for the production of recombinant proteins in Escherichia coli. J. Microbiol. Biotechnol. 2004, 14, 1–14. [Google Scholar]
- Carrier, T.; Jones, K.L.; Keasling, J.D. mRNA stability and plasmid copy number effects on gene expression from an inducible promoter system. Biotechnol. Bioeng. 1998, 59, 666–672. [Google Scholar] [CrossRef]
- Bentley, W.E.; Mirjalili, N.; Andersen, D.C.; Davis, R.H.; Kompala, D.S. Plasmid-encoded protein: The principal factor in the “metabolic burden” associated with recombinant bacteria. Biotechnol. Bioeng. 1990, 35, 668–681. [Google Scholar] [CrossRef]
- Birnbaum, S.; Bailey, J.E. Plasmid presence changes the relative levels of many host cell proteins and ribosome components in recombinant Escherichia coli. Biotechnol. Bioeng. 1991, 37, 736–745. [Google Scholar] [CrossRef]
- Davies, D.G.; Geesey, G.G. Regulation of the alginate biosynthesis gene algC in Pseudomonas aeruginosa during biofilm development in continuous culture. Appl. Environ. Microbiol. 1995, 61, 860–867. [Google Scholar] [CrossRef] [Green Version]
- Cook, L.C.; Dunny, G.M. Effects of biofilm growth on plasmid copy number and expression of antibiotic resistance genes in Enterococcus faecalis. Antimicrob. Agents Chemother. 2013, 57, 1850–1856. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, H.A.; Niu, C.; Gilbert, E.S. Enhanced high copy number plasmid maintenance and heterologous protein production in an Escherichia coli biofilm. Biotechnol. Bioeng. 2007, 97, 439–446. [Google Scholar] [CrossRef] [PubMed]
- May, T.; Ito, A.; Okabe, S. Induction of multidrug resistance mechanism in Escherichia coli biofilms by interplay between tetracycline and ampicillin resistance genes. Antimicrob. Agents Chemother. 2009, 53, 4628–4639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donlan, R.M. Role of biofilms in antimicrobial resistance. ASAIO J. 2000, 46, S47–S52. [Google Scholar] [CrossRef]
- Huang, C.-T.; Peretti, S.W.; Bryers, J.D. Plasmid retention and gene expression in suspended and biofilm cultures of recombinant Escherichia coli DH5α(pMJR1750). Biotechnol. Bioeng. 1993, 41, 211–220. [Google Scholar] [CrossRef]
- Ma, H.; Katzenmeyer, K.N.; Bryers, J.D. Non-invasive in situ monitoring and quantification of TOL plasmid segregational loss within Pseudomonas putida biofilms. Biotechnol. Bioeng. 2013, 110, 2949–2958. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.S.; Burmølle, M.; Sørensen, S.J. A spatiotemporal view of plasmid loss in biofilms and planktonic cultures. Biotechnol. Bioeng. 2013, 110, 3071–3074. [Google Scholar] [CrossRef]
- Gomes, L.C.; Mergulhão, F.J. Heterologous protein production in Escherichia coli biofilms: A non-conventional form of high cell density cultivation. Process. Biochem. 2017, 57, 1–8. [Google Scholar] [CrossRef]
- Stoodley, P.; Warwood, B.K. Use of flow cells an annular reactors to study biofilms. In Biofilms in Medicine, Industry and Environmental Biotechnology: Characteristics, Analysis and Control; Lens, P., O’Flaherty, V., Moran, A.P., Stoodley, P., Mahony, T., Eds.; IWA Publishing: Cornwall, UK, 2003; pp. 197–213. [Google Scholar]
- Teodósio, J.S.; Simões, M.; Alves, M.A.; Melo, L.F.; Mergulhão, F.J. Setup and validation of flow cell systems for biofouling simulation in industrial settings. Sci. World J. 2012, 2012, 10. [Google Scholar] [CrossRef]
- Sharma, S.S.; Blattner, F.R.; Harcum, S.W. Recombinant protein production in an Escherichia coli reduced genome strain. Metab. Eng. 2007, 9, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Sandén, A.M.; Prytz, I.; Tubulekas, I.; Förberg, C.; Le, H.; Hektor, A.; Neubauer, P.; Pragai, Z.; Harwood, C.; Ward, A.; et al. Limiting factors in Escherichia coli fed-batch production of recombinant proteins. Biotechnol. Bioeng. 2003, 81, 158–166. [Google Scholar] [CrossRef]
- Schaepe, S.; Kuprijanov, A.; Simutis, R.; Lübbert, A. Avoiding overfeeding in high cell density fed-batch cultures of E. coli during the production of heterologous proteins. J. Biotechnol. 2014, 192, 146–153. [Google Scholar] [CrossRef]
- Bentley, W.E.; Davis, R.H.; Kompala, D.S. Dynamics of induced CAT expression in E. coli. Biotechnol. Bioeng. 1991, 38, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, P.; Hofmann, K.; Holst, O.; Mattiasson, B.; Kruschke, P. Maximizing the expression of a recombinant gene in Escherichia coli by manipulation of induction time using lactose as inducer. Appl. Microbiol. Biotechnol. 1992, 36, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.K.; Peretti, S.W. Effect of chemically-induced, cloned-gene expression on protein synthesis in E. coli. Biotechnol. Bioeng. 1991, 38, 397–412. [Google Scholar] [CrossRef]
- Werner, E.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Heydorn, A.; Molin, S.; Pitts, B.; Stewart, P.S. Stratified growth in Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 2004, 70, 6188–6196. [Google Scholar] [CrossRef] [Green Version]
- Stewart, P.S. Diffusion in biofilms. J. Bacteriol. 2003, 185, 1485–1491. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Castané, A.; Caminal, G.; López-Santín, J. Direct measurements of IPTG enable analysis of the induction behavior of E. coli in high cell density cultures. Microb. Cell Fact. 2012, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-T.; Peretti, S.W.; Bryers, J.D. Effects of inducer levels on a recombinant bacterial biofilm formation and gene expression. Biotechnol. Lett. 1994, 16, 903–908. [Google Scholar] [CrossRef]
- Teodósio, J.S.; Simões, M.; Melo, L.F.; Mergulhão, F.J. Flow cell hydrodynamics and their effects on E. coli biofilm formation under different nutrient conditions and turbulent flow. Biofouling 2011, 27, 1–11. [Google Scholar] [CrossRef]
- Lenz, A.P.; Williamson, K.S.; Pitts, B.; Stewart, P.S.; Franklin, M.J. Localized gene expression in Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 2008, 74, 4463–4471. [Google Scholar] [CrossRef] [Green Version]
- Bryers, J.D.; Huang, C.-T. Recombinant plasmid retention and expression in bacterial biofilm cultures. Water Sci. Technol. 1995, 31, 105–115. [Google Scholar] [CrossRef]
- Berg, J.; Tymoczko, J.; Stryer, L. Gene expression is the transformation of DNA information into functional molecules. In Biochemistry, 5th ed.; Berg, J., Tymoczko, J., Stryer, L., Eds.; W.H. Freeman and Company: New York, NY, USA, 2002. [Google Scholar]
- Dong, H.; Nilsson, L.; Kurland, C.G. Gratuitous overexpression of genes in Escherichia coli leads to growth inhibition and ribosome destruction. J. Bacteriol. 1995, 177, 1497–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Kuipers, G.; Niemiec, Ł.; Baumgarten, T.; Slotboom, D.J.; de Gier, J.-W.; Hjelm, A. High-level production of membrane proteins in E. coli BL21(DE3) by omitting the inducer IPTG. Microb. Cell Fact. 2015, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.L.; Kim, S.-W.; Keasling, J.D. Low-copy plasmids can perform as well as or better than high-copy plasmids for metabolic engineering of bacteria. Metab. Eng. 2000, 2, 328–338. [Google Scholar] [CrossRef]
- Lowder, M.; Unge, A.; Maraha, N.; Jansson, J.K.; Swiggett, J.; Oliver, J.D. Effect of starvation and the viable-but-nonculturable state on green fluorescent protein (GFP) fluorescence in GFP-tagged Pseudomonas fluorescens A506. Appl. Environ. Microbiol. 2000, 66, 3160–3165. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Dubey, A. Metabolic burden as reflected by maintenance coefficient of recombinant Escherichia coli overexpressing target gene. Biotechnol. Lett. 1995, 17, 1155–1160. [Google Scholar] [CrossRef]
- Diaz Ricci, J.C.; Hernández, M.E. Plasmid effects on Escherichia coli metabolism. Crit. Rev. Biotechnol. 2000, 20, 79–108. [Google Scholar] [CrossRef]
- Mergulhão, F.J.; Monteiro, G.A. Analysis of factors affecting the periplasmic production of recombinant proteins in Escherichia coli. J. Microbiol. Biotechnol. 2007, 17, 1236–1241. [Google Scholar]
- Gomes, L.C.; Mergulhão, F.J. Effects of antibiotic concentration and nutrient medium composition on Escherichia coli biofilm formation and green fluorescent protein expression. FEMS Microbiol. Lett. 2017, 364, fnx042. [Google Scholar] [CrossRef] [Green Version]
- Teodósio, J.S.; Simões, M.; Mergulhão, F.J. The influence of nonconjugative Escherichia coli plasmids on biofilm formation and resistance. J. Appl. Microbiol. 2012, 113, 373–382. [Google Scholar] [CrossRef]
- Fogler, H.S. Distributions of residence times for chemical reactors. In Elements of Chemical Reaction Engineering, 4th ed.; Fogler, H.S., Ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2006. [Google Scholar]
- Perry, R.H.; Green, D.W. Perry’s Chemical Engineers’ Handbook; McGraw-Hill: New York, NY, USA, 1997. [Google Scholar]
- Gomes, L.C.; Carvalho, D.; Briandet, R.; Mergulhão, F.J. Temporal variation of recombinant protein expression in Escherichia coli biofilms analysed at single-cell level. Process. Biochem. 2016, 51, 1155–1161. [Google Scholar] [CrossRef] [Green Version]
- Gomes, L.C.; Silva, L.N.; Simões, M.; Melo, L.F.; Mergulhão, F.J. Escherichia coli adhesion, biofilm development and antibiotic susceptibility on biomedical materials. J. Biomed. Mater. Res. A 2015, 103, 1414–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Simões, M.; Pereira, M.O.; Sillankorva, S.; Azeredo, J.; Vieira, M.J. The effect of hydrodynamic conditions on the phenotype of Pseudomonas fluorescens biofilms. Biofouling 2007, 23, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carapuça, E.; Azzoni, A.R.; Prazeres, D.M.; Monteiro, G.A.; Mergulhão, F.J. Time-course determination of plasmid content in eukaryotic and prokaryotic cells using real-time PCR. Mol. Biotechnol. 2007, 37, 120–126. [Google Scholar] [CrossRef]
- Lee, C.; Kim, J.; Shin, S.G.; Hwang, S. Absolute and relative QPCR quantification of plasmid copy number in Escherichia coli. J. Biotechnol. 2006, 123, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A method for the absolute quantification of cDNA using real-time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef]
 )). The averages ± SDs (indicated as bars) are illustrated. Statistical analysis corresponding to each time point is represented with
)). The averages ± SDs (indicated as bars) are illustrated. Statistical analysis corresponding to each time point is represented with  for a confidence level greater than 90% (p < 0.1) and with
for a confidence level greater than 90% (p < 0.1) and with  for a confidence level greater than 95% (p < 0.05).
)). The averages ± SDs (indicated as bars) are illustrated. Statistical analysis corresponding to each time point is represented with for a confidence level greater than 90% (p < 0.1) and with for a confidence level greater than 95% (p < 0.05).
for a confidence level greater than 95% (p < 0.05).
)). The averages ± SDs (indicated as bars) are illustrated. Statistical analysis corresponding to each time point is represented with for a confidence level greater than 90% (p < 0.1) and with for a confidence level greater than 95% (p < 0.05).
 ) and planktonic (
) and planktonic (  ) cells). The averages ± SDs (indicated as bars) are illustrated. Statistical analysis for a confidence level greater than 95% (p < 0.05) is pointed as † when induced biofilm cells are different from the non-induced biofilm cells and as ‡ when induced planktonic cells are different from the non-induced planktonic cells.
) and planktonic ( ) cells). The averages ± SDs (indicated as bars) are illustrated. Statistical analysis for a confidence level greater than 95% (p < 0.05) is pointed as † when induced biofilm cells are different from the non-induced biofilm cells and as ‡ when induced planktonic cells are different from the non-induced planktonic cells.
) cells). The averages ± SDs (indicated as bars) are illustrated. Statistical analysis for a confidence level greater than 95% (p < 0.05) is pointed as † when induced biofilm cells are different from the non-induced biofilm cells and as ‡ when induced planktonic cells are different from the non-induced planktonic cells.
) and planktonic ( ) cells). The averages ± SDs (indicated as bars) are illustrated. Statistical analysis for a confidence level greater than 95% (p < 0.05) is pointed as † when induced biofilm cells are different from the non-induced biofilm cells and as ‡ when induced planktonic cells are different from the non-induced planktonic cells.
 for a confidence level greater than 90% (p < 0.1) and by
for a confidence level greater than 90% (p < 0.1) and by  for a confidence level greater than 95% (p < 0.05).
for a confidence level greater than 90% (p < 0.1) and by for a confidence level greater than 95% (p < 0.05).
for a confidence level greater than 95% (p < 0.05).
for a confidence level greater than 90% (p < 0.1) and by for a confidence level greater than 95% (p < 0.05).

{kind=link}
{kind=link}
{kind=link}
| Biofilm Characteristics | Induced | Non-Induced |
|---|---|---|
| Biofilm mass (mgbiofilm cm−2) | 4.00 ± 1.17 | 4.50 ± 0.870 |
| Log cellular density (cells·cm−2) | 7.83 ± 0.460 | 7.76 ± 0.460 |
| Matrix proteins (mg g−1biofilm) | 8.60 ± 1.80 | 9.40 ± 1.70 |
| Matrix polysaccharides (mg g−1biofilm) | 249 ± 41.0 | 231 ± 35.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, L.; Monteiro, G.; Mergulhão, F. The Impact of IPTG Induction on Plasmid Stability and Heterologous Protein Expression by Escherichia coli Biofilms. Int. J. Mol. Sci. 2020, 21, 576. https://doi.org/10.3390/ijms21020576
Gomes L, Monteiro G, Mergulhão F. The Impact of IPTG Induction on Plasmid Stability and Heterologous Protein Expression by Escherichia coli Biofilms. International Journal of Molecular Sciences. 2020; 21(2):576. https://doi.org/10.3390/ijms21020576
Chicago/Turabian StyleGomes, Luciana, Gabriel Monteiro, and Filipe Mergulhão. 2020. "The Impact of IPTG Induction on Plasmid Stability and Heterologous Protein Expression by Escherichia coli Biofilms" International Journal of Molecular Sciences 21, no. 2: 576. https://doi.org/10.3390/ijms21020576
APA StyleGomes, L., Monteiro, G., & Mergulhão, F. (2020). The Impact of IPTG Induction on Plasmid Stability and Heterologous Protein Expression by Escherichia coli Biofilms. International Journal of Molecular Sciences, 21(2), 576. https://doi.org/10.3390/ijms21020576