Comprehensive Analyses of Cytochrome P450 Monooxygenases and Secondary Metabolite Biosynthetic Gene Clusters in Cyanobacteria

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
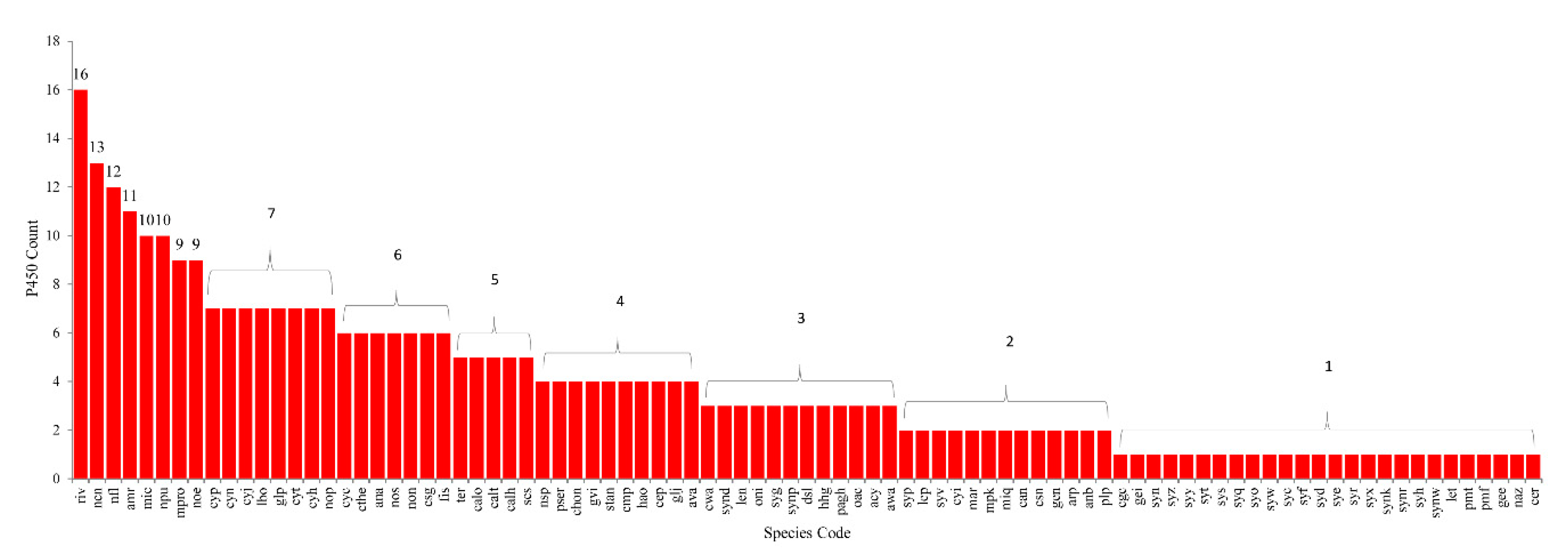
2.1. Cyanobacterial Species Have Lowest Number of P450s
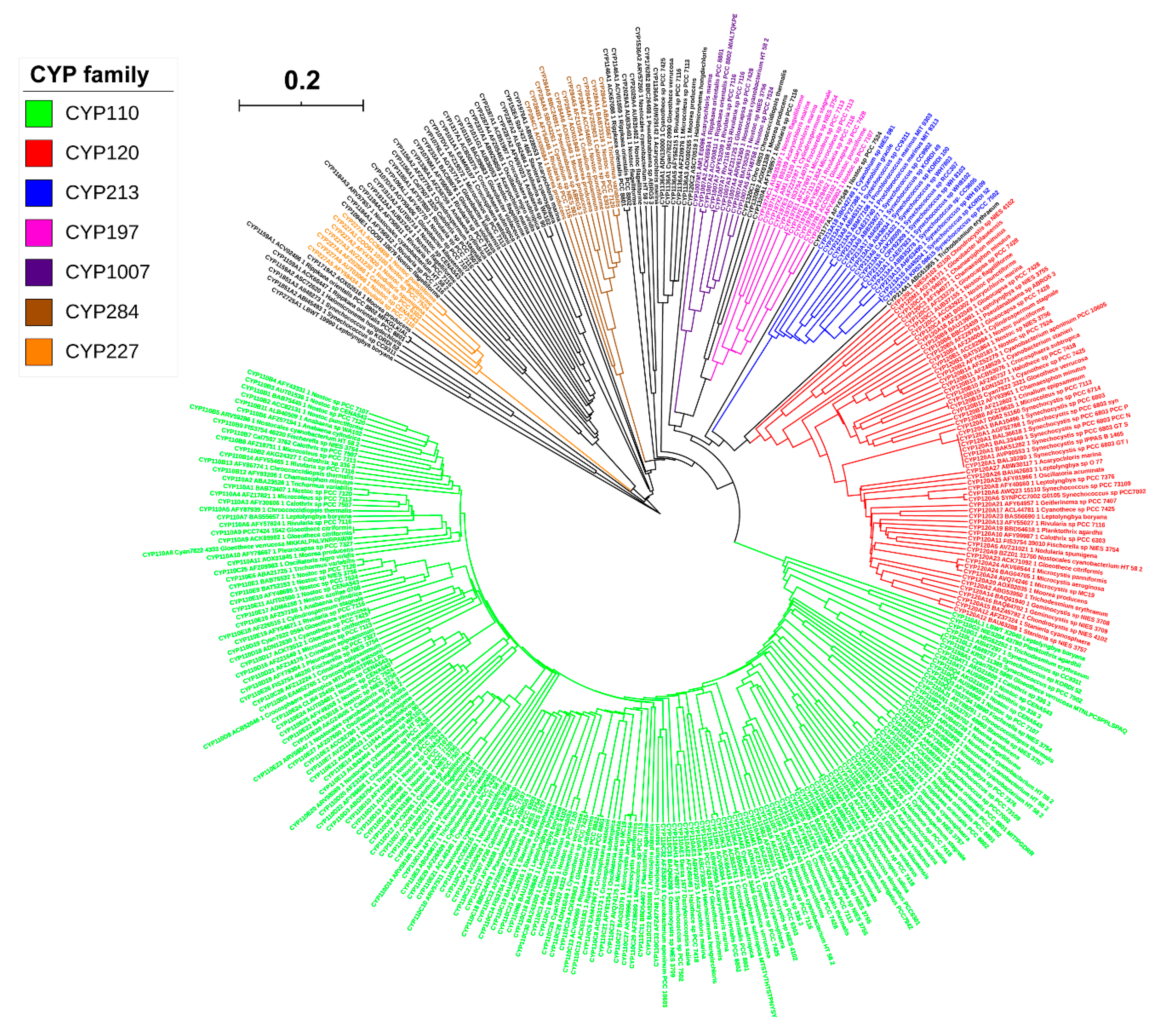
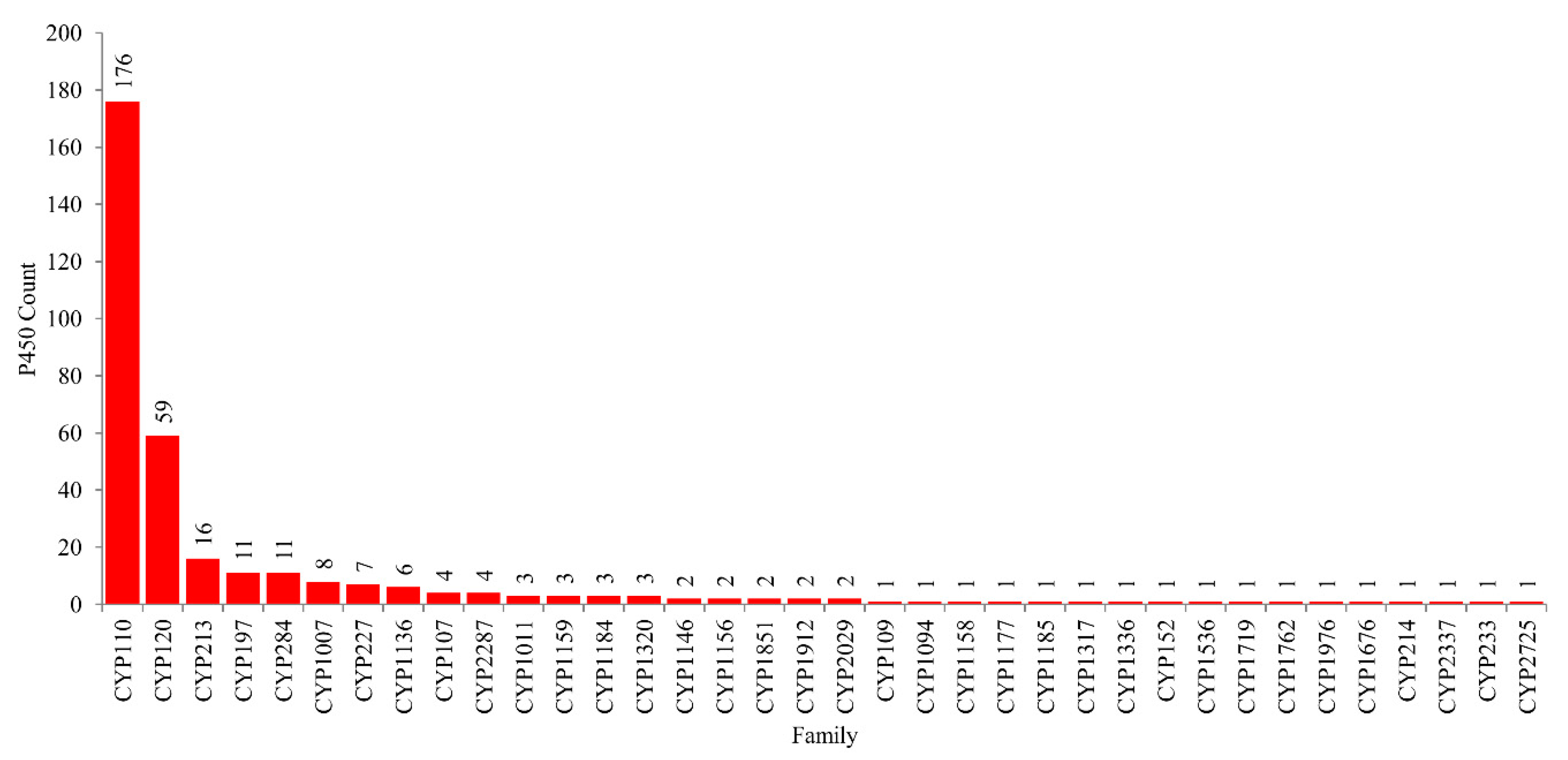
2.2. CYP110 is the Dominant P450 Family in Cyanobacterial Species
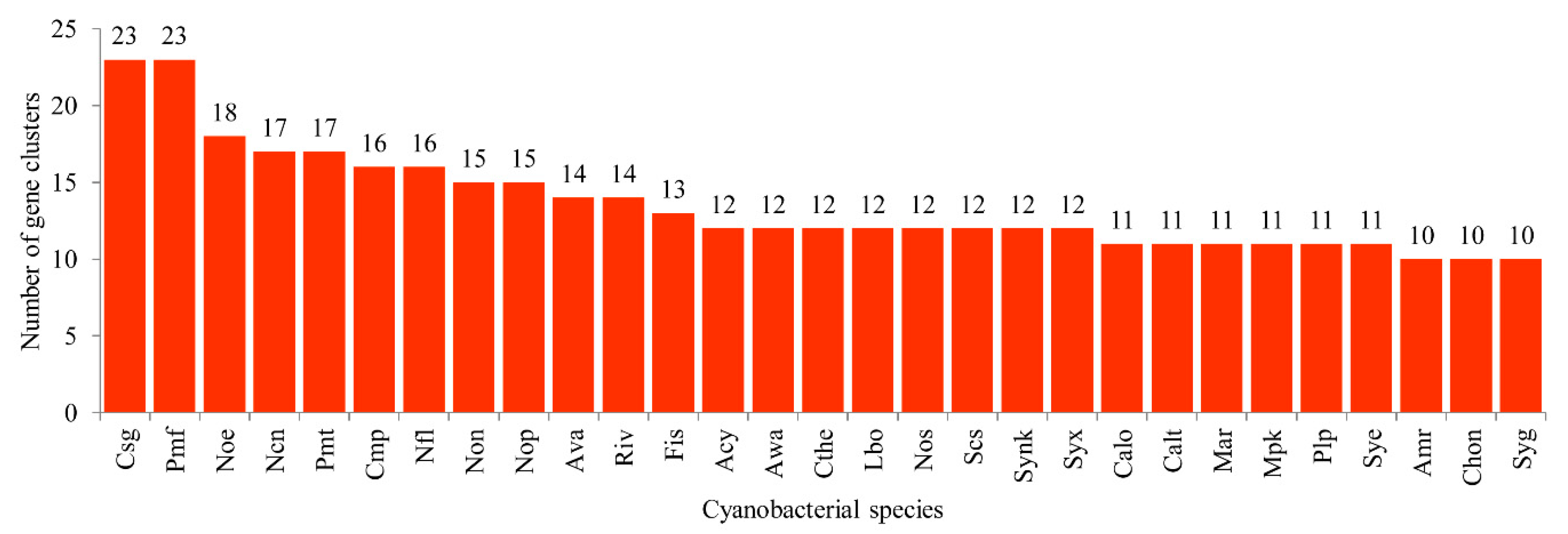
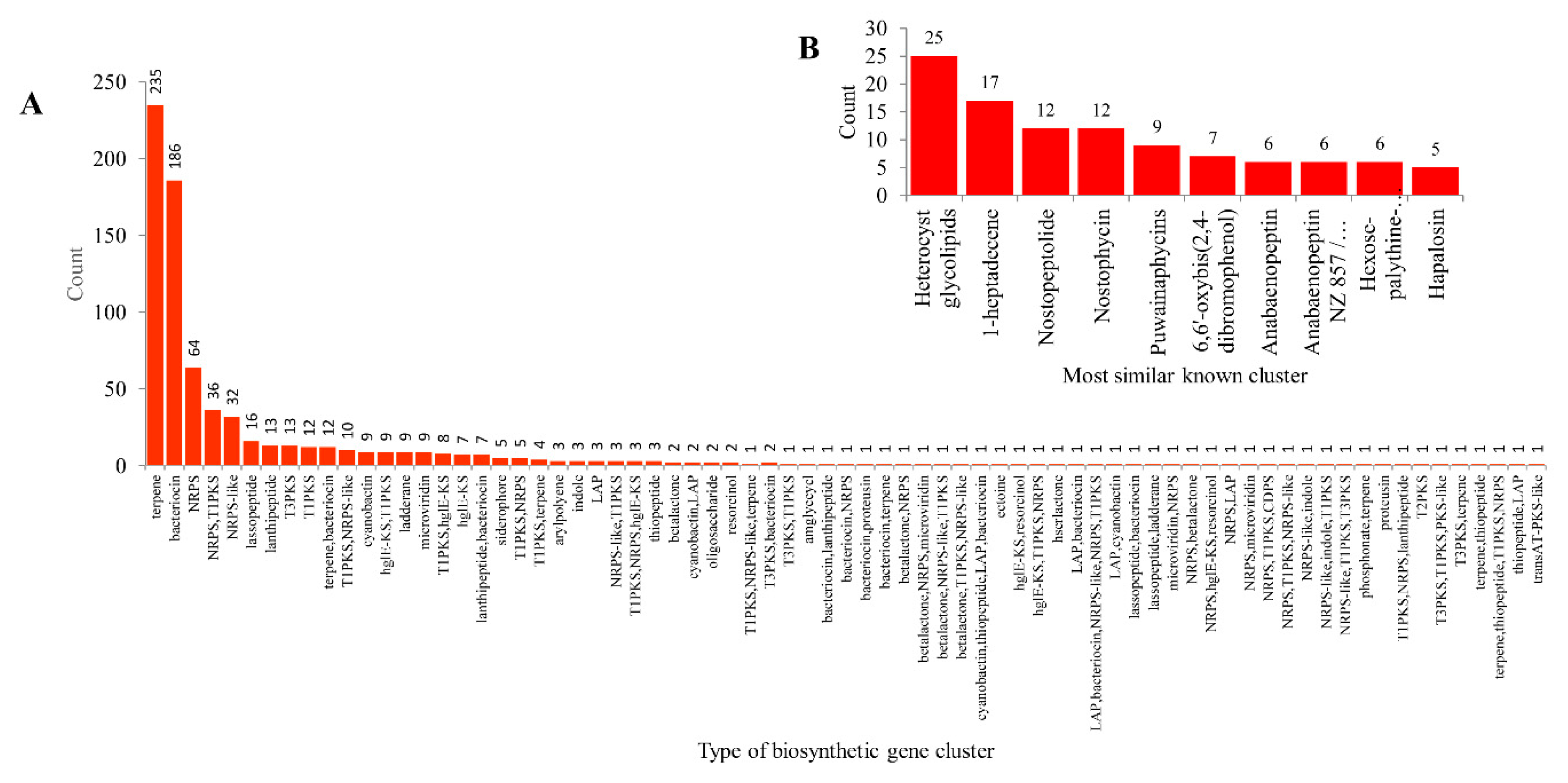
2.3. Cyanobacterial Species Have Lowest Secondary Metabolite Biosynthetic Gene Clusters
2.4. Cyanobacterial Species Has Highest Gene Cluster Diversity Percentage Compared to Bacillus and Mycobacterial Species
2.5. Few Cyanobacterial Species P450s Found to be Part of Secondary Metabolite Biosynthetic Gene Clusters
2.6. Cyanobacterial Species P450s Functions and Features Resemblance to Eukaryotic P450s
3. Materials and Methods
3.1. Species and Databases
3.2. Genome Data Mining and Annotation of P450s
3.3. Phylogenetic Tree Construction of Cyanobacterial Species P450s
3.4. Generation of P450 Profile Heat-Maps
3.5. Secondary Metabolite Biosynthetic Gene Clusters Analysis
3.6. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bekker, A.; Holland, H.; Wang, P.-L.; Rumble Iii, D.; Stein, H.; Hannah, J.; Coetzee, L.; Beukes, N. Dating the rise of atmospheric oxygen. Nature 2004, 427, 117–120. [Google Scholar] [CrossRef]
- Kump, L.R. The rise of atmospheric oxygen. Nature 2008, 451, 277–278. [Google Scholar] [CrossRef]
- Shih, P.M. Cyanobacterial evolution: Fresh insight into ancient questions. Curr. Biol. 2015, 25, R192–R193. [Google Scholar] [CrossRef] [Green Version]
- Flores, F.G. The Cyanobacteria: Molecular Biology, Genomics, and Evolution; Horizon Scientific Press: Heatherset, UK, 2008. [Google Scholar]
- Buick, R. The antiquity of oxygenic photosynthesis: Evidence from stromatolites in sulphate-deficient Archaean lakes. Science 1992, 255, 74–77. [Google Scholar] [CrossRef]
- Raven, J.A.; Allen, J.F. Genomics and chloroplast evolution: What did cyanobacteria do for plants? Genome Biol. 2003, 4, 209. [Google Scholar] [CrossRef] [Green Version]
- Willis, K.; McElwain, J. The Evolution of Plants; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Gaysina, L.A.; Saraf, A.; Singh, P. Cyanobacteria in Diverse Habitats. In Cyanobacteria; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–28. [Google Scholar]
- Dvořák, P.; Casamatta, D.A.; Hašler, P.; Jahodářová, E.; Norwich, A.R.; Poulíčková, A. Diversity of the cyanobacteria. In Modern Topics in the Phototrophic Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2017; pp. 3–46. [Google Scholar]
- Kultschar, B.; Llewellyn, C. Secondary Metabolites in Cyanobacteria. Second. Metab. Sources Appl. 2018, 23. [Google Scholar] [CrossRef] [Green Version]
- Robert, F.O.; Pandhal, J.; Wright, P.C. Exploiting cyanobacterial P450 pathways. Curr. Opin. Microbiol. 2010, 13, 301–306. [Google Scholar] [CrossRef]
- Cohen, S.E.; Golden, S.S. Circadian rhythms in cyanobacteria. Microbiol. Mol. Biol. Rev. 2015, 79, 373–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, P.E.; Leister, D. Cyanobacteria as an experimental platform for modifying bacterial and plant photosynthesis. Front. Bioeng. Biotechnol. 2014, 2, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bothe, H.; Schmitz, O.; Yates, M.G.; Newton, W.E. Nitrogen fixation and hydrogen metabolism in cyanobacteria. Microbiol. Mol. Biol. Rev. 2010, 74, 529–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Merino, M.; Singh, A.K.; Ducat, D.C. New applications of synthetic biology tools for cyanobacterial metabolic engineering. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Wilde, A.; Dienst, D. Tools for genetic manipulation of cyanobacteria. In Bioenergetic Processes of Cyanobacteria; Springer: Berlin/Heidelberg, Germany, 2011; pp. 685–703. [Google Scholar]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Swingley, W.D.; Chen, M.; Cheung, P.C.; Conrad, A.L.; Dejesa, L.C.; Hao, J.; Honchak, B.M.; Karbach, L.E.; Kurdoglu, A.; Lahiri, S.; et al. Niche adaptation and genome expansion in the chlorophyll d-producing cyanobacterium Acaryochloris marina. Proc. Natl. Acad. Sci. USA 2008, 105, 2005–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, N.M.; Mueller, R.S.; Shepardson, J.W.; Landry, Z.C.; Morre, J.T.; Maier, C.S.; Hardy, F.J.; Dreher, T.W. Structural and functional analysis of the finished genome of the recently isolated toxic Anabaena sp. WA102. BMC Genom. 2016, 17, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, T.; Tanaka, A.; Sato, S.; Kotani, H.; Sazuka, T.; Miyajima, N.; Sugiura, M.; Tabata, S. Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. I. Sequence features in the 1 Mb region from map positions 64% to 92% of the genome. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 1995, 2, 153–166, 191–198. [Google Scholar]
- Sugita, C.; Ogata, K.; Shikata, M.; Jikuya, H.; Takano, J.; Furumichi, M.; Kanehisa, M.; Omata, T.; Sugiura, M.; Sugita, M. Complete nucleotide sequence of the freshwater unicellular cyanobacterium Synechococcus elongatus PCC 6301 chromosome: Gene content and organization. Photosynth. Res. 2007, 93, 55–67. [Google Scholar] [CrossRef]
- Palenik, B.; Brahamsha, B.; Larimer, F.W.; Land, M.; Hauser, L.; Chain, P.; Lamerdin, J.; Regala, W.; Allen, E.E.; McCarren, J.; et al. The genome of a motile marine Synechococcus. Nature 2003, 424, 1037–1042. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Kaneko, T.; Sato, S.; Ikeuchi, M.; Katoh, H.; Sasamoto, S.; Watanabe, A.; Iriguchi, M.; Kawashima, K.; Kimura, T.; et al. Complete genome structure of the thermophilic cyanobacterium Thermosynechococcus elongatus BP-1. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2002, 9, 123–130. [Google Scholar]
- Yamaguchi, H.; Shimura, Y.; Suzuki, S.; Yamagishi, T.; Tatarazako, N.; Kawachi, M. Complete Genome Sequence of Cyanobium sp. NIES-981, a Marine Strain Potentially Useful for Ecotoxicological Bioassays. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Walsby, A.; Van Rijn, J.; Cohen, Y. The biology of a new gas-vacuolate cyanobacterium, Dactylococcopsis salina sp. nov., in Solar Lake. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1983, 217, 417–447. [Google Scholar]
- Kučera, P.; Uher, B.; Komárek, O. Epiphytic cyanophytes Xenococcus kerneri and Chamaesiphon minutus on the freshwater red alga Paralemanea catenata (Rhodophyta). Biologia 2006, 61, 11–13. [Google Scholar] [CrossRef]
- Hirose, Y.; Fujisawa, T.; Ohtsubo, Y.; Katayama, M.; Misawa, N.; Wakazuki, S.; Shimura, Y.; Nakamura, Y.; Kawachi, M.; Yoshikawa, H.; et al. Complete Genome Sequence of Cyanobacterium Leptolyngbya sp. NIES-3755. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimura, Y.; Hirose, Y.; Misawa, N.; Osana, Y.; Katoh, H.; Yamaguchi, H.; Kawachi, M. Comparison of the terrestrial cyanobacterium Leptolyngbya sp. NIES-2104 and the freshwater Leptolyngbya boryana PCC 6306 genomes. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2015, 22, 403–412. [Google Scholar]
- Bruno, L.; Billi, D.; Bellezza, S.; Albertano, P. Cytomorphological and genetic characterization of troglobitic Leptolyngbya strains isolated from Roman hypogea. Appl. Environ. Microbiol. 2009, 75, 608–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Hernandez-Prieto, M.A.; Loughlin, P.C.; Li, Y.; Willows, R.D. Genome and proteome of the chlorophyll f-producing cyanobacterium Halomicronema hongdechloris: Adaptative proteomic shifts under different light conditions. BMC Genom. 2019, 20, 207. [Google Scholar] [CrossRef]
- Chen, M.; Schliep, M.; Willows, R.D.; Cai, Z.L.; Neilan, B.A.; Scheer, H. A red-shifted chlorophyll. Science 2010, 329, 1318–1319. [Google Scholar] [CrossRef]
- Tajima, N.; Kanesaki, Y.; Sato, S.; Yoshikawa, H.; Maruyama, F.; Kurokawa, K.; Ohta, H.; Nishizawa, T.; Asayama, M.; Sato, N. Complete Genome Sequence of the Nonheterocystous Cyanobacterium Pseudanabaena sp. ABRG5-3. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Dufresne, A.; Salanoubat, M.; Partensky, F.; Artiguenave, F.; Axmann, I.M.; Barbe, V.; Duprat, S.; Galperin, M.Y.; Koonin, E.V.; Le Gall, F.; et al. Genome sequence of the cyanobacterium Prochlorococcus marinus SS120, a nearly minimal oxyphototrophic genome. Proc. Natl. Acad. Sci. USA 2003, 100, 10020–10025. [Google Scholar] [CrossRef] [Green Version]
- Hirose, Y.; Katayama, M.; Ohtsubo, Y.; Misawa, N.; Iioka, E.; Suda, W.; Oshima, K.; Hanaoka, M.; Tanaka, K.; Eki, T.; et al. Complete Genome Sequence of Cyanobacterium Geminocystis sp. Strain NIES-3709, Which Harbors a Phycoerythrin-Rich Phycobilisome. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, T.; Nakajima, N.; Okamoto, S.; Suzuki, I.; Tanabe, Y.; Tamaoki, M.; Nakamura, Y.; Kasai, F.; Watanabe, A.; Kawashima, K.; et al. Complete genomic structure of the bloom-forming toxic cyanobacterium Microcystis aeruginosa NIES-843. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2007, 14, 247–256. [Google Scholar] [CrossRef]
- Mobberley, J.M.; Romine, M.F.; Cole, J.K.; Maezato, Y.; Lindemann, S.R.; Nelson, W.C. Draft Genome Sequence of Cyanobacterium sp. Strain HL-69, Isolated from a Benthic Microbial Mat from a Magnesium Sulfate-Dominated Hypersaline Lake. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bench, S.R.; Heller, P.; Frank, I.; Arciniega, M.; Shilova, I.N.; Zehr, J.P. Whole genome comparison of six Crocosphaera watsonii strains with differing phenotypes. J. Phycol. 2013, 49, 786–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, E.A.; Liberton, M.; Stockel, J.; Loh, T.; Elvitigala, T.; Wang, C.; Wollam, A.; Fulton, R.S.; Clifton, S.W.; Jacobs, J.M.; et al. The genome of Cyanothece 51142, a unicellular diazotrophic cyanobacterium important in the marine nitrogen cycle. Proc. Natl. Acad. Sci. USA 2008, 105, 15094–15099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capone, D.G.; Burns, J.A.; Montoya, J.P.; Subramaniam, A.; Mahaffey, C.; Gunderson, T.; Michaels, A.F.; Carpenter, E.J. Nitrogen fixation by Trichodesmium spp.: An important source of new nitrogen to the tropical and subtropical North Atlantic Ocean. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Fujisawa, T.; Narikawa, R.; Okamoto, S.; Ehira, S.; Yoshimura, H.; Suzuki, I.; Masuda, T.; Mochimaru, M.; Takaichi, S.; Awai, K.; et al. Genomic structure of an economically important cyanobacterium, Arthrospira (Spirulina) platensis NIES-39. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2010, 17, 85–103. [Google Scholar] [CrossRef] [Green Version]
- Churro, C.; Azevedo, J.; Vasconcelos, V.; Silva, A. Detection of a Planktothrix agardhii Bloom in Portuguese Marine Coastal Waters. Toxins 2017, 9, 391. [Google Scholar] [CrossRef] [Green Version]
- Leao, T.; Castelao, G.; Korobeynikov, A.; Monroe, E.A.; Podell, S.; Glukhov, E.; Allen, E.E.; Gerwick, W.H.; Gerwick, L. Comparative genomics uncovers the prolific and distinctive metabolic potential of the cyanobacterial genus Moorea. Proc. Natl. Acad. Sci. USA 2017, 114, 3198–3203. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Kaneko, T.; Sato, S.; Mimuro, M.; Miyashita, H.; Tsuchiya, T.; Sasamoto, S.; Watanabe, A.; Kawashima, K.; Kishida, Y.; et al. Complete genome structure of Gloeobacter violaceus PCC 7421, a cyanobacterium that lacks thylakoids. Dna Res. Int. J. Rapid Publ. Rep. Genes Genomes 2003, 10, 137–145. [Google Scholar] [CrossRef]
- Kaneko, T.; Nakamura, Y.; Wolk, C.P.; Kuritz, T.; Sasamoto, S.; Watanabe, A.; Iriguchi, M.; Ishikawa, A.; Kawashima, K.; Kimura, T.; et al. Complete genomic sequence of the filamentous nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2001, 8, 205–213, 227–253. [Google Scholar]
- Ekman, M.; Picossi, S.; Campbell, E.L.; Meeks, J.C.; Flores, E. A Nostoc punctiforme sugar transporter necessary to establish a Cyanobacterium-plant symbiosis. Plant Physiol. 2013, 161, 1984–1992. [Google Scholar] [CrossRef] [Green Version]
- Ran, L.; Larsson, J.; Vigil-Stenman, T.; Nylander, J.A.; Ininbergs, K.; Zheng, W.W.; Lapidus, A.; Lowry, S.; Haselkorn, R.; Bergman, B. Genome erosion in a nitrogen-fixing vertically transmitted endosymbiotic multicellular cyanobacterium. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Wang, H.; Sivonen, K.; Rouhiainen, L.; Fewer, D.P.; Lyra, C.; Rantala-Ylinen, A.; Vestola, J.; Jokela, J.; Rantasarkka, K.; Li, Z.; et al. Genome-derived insights into the biology of the hepatotoxic bloom-forming cyanobacterium Anabaena sp. strain 90. BMC Genom. 2012, 13, 613. [Google Scholar] [CrossRef] [Green Version]
- Isojarvi, J.; Shunmugam, S.; Sivonen, K.; Allahverdiyeva, Y.; Aro, E.M.; Battchikova, N. Draft genome sequence of Calothrix strain 336/3, a novel h2-producing cyanobacterium isolated from a finnish lake. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Hirose, Y.; Fujisawa, T.; Ohtsubo, Y.; Katayama, M.; Misawa, N.; Wakazuki, S.; Shimura, Y.; Nakamura, Y.; Kawachi, M.; Yoshikawa, H.; et al. Complete genome sequence of cyanobacterium Fischerella sp. NIES-3754, providing thermoresistant optogenetic tools. J. Biotechnol. 2016, 220, 45–46. [Google Scholar] [CrossRef]
- Teikari, J.E.; Hou, S.; Wahlsten, M.; Hess, W.R.; Sivonen, K. Comparative Genomics of the Baltic Sea Toxic Cyanobacteria Nodularia spumigena UHCC 0039 and Its Response to Varying Salinity. Front. Microbiol. 2018, 9, 356. [Google Scholar] [CrossRef] [Green Version]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Wieland Brown, L.C.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Podust, L.M.; Sherman, D.H. Diversity of P450 enzymes in the biosynthesis of natural products. Nat. Prod. Rep. 2012, 29, 1251–1266. [Google Scholar] [CrossRef] [Green Version]
- Greule, A.; Stok, J.E.; De Voss, J.J.; Cryle, M.J. Unrivalled diversity: The many roles and reactions of bacterial cytochromes P450 in secondary metabolism. Nat. Prod. Rep. 2018, 35, 757–791. [Google Scholar] [CrossRef] [Green Version]
- Mthethwa, B.; Chen, W.; Ngwenya, M.; Kappo, A.; Syed, P.; Karpoormath, R.; Yu, J.-H.; Nelson, D.; Syed, K. Comparative analyses of cytochrome P450s and those associated with secondary metabolism in Bacillus species. Int. J. Mol. Sci. 2018, 19, 3623. [Google Scholar] [CrossRef] [Green Version]
- Senate, L.M.; Tjatji, M.P.; Pillay, K.; Chen, W.; Zondo, N.M.; Syed, P.R.; Mnguni, F.C.; Chiliza, Z.E.; Bamal, H.D.; Karpoormath, R. Similarities, variations, and evolution of cytochrome P450s in Streptomyces versus Mycobacterium. Sci. Rep. 2019, 9, 3962. [Google Scholar] [CrossRef] [Green Version]
- Syed, P.R.; Chen, W.; Nelson, D.R.; Kappo, A.P.; Yu, J.-H.; Karpoormath, R.; Syed, K. Cytochrome P450 Monooxygenase CYP139 Family Involved in the Synthesis of Secondary Metabolites in 824 Mycobacterial Species. Int. J. Mol. Sci. 2019, 20, 2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agger, S.A.; Lopez-Gallego, F.; Hoye, T.R.; Schmidt-Dannert, C. Identification of sesquiterpene synthases from Nostoc punctiforme PCC 73102 and Nostoc sp. strain PCC 7120. J. Bacteriol. 2008, 190, 6084–6096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhnel, K.; Ke, N.; Cryle, M.J.; Sligar, S.G.; Schuler, M.A.; Schlichting, I. Crystal structures of substrate-free and retinoic acid-bound cyanobacterial cytochrome P450 CYP120A1. Biochemistry 2008, 47, 6552–6559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvez, M.; Qhanya, L.B.; Mthakathi, N.T.; Kgosiemang, I.K.R.; Bamal, H.D.; Pagadala, N.S.; Xie, T.; Yang, H.; Chen, H.; Theron, C.W. Molecular evolutionary dynamics of cytochrome P450 monooxygenases across kingdoms: Special focus on mycobacterial P450s. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R. Cytochrome P450 nomenclature. Methods Mol. Biol. (Clifton N. J.) 1998, 107, 15–24. [Google Scholar]
- Nelson, D.R. Cytochrome P450 nomenclature, 2004. Methods Mol. Biol. (Clifton N. J.) 2006, 320, 1–10. [Google Scholar]
- Nelson, D.R. The cytochrome p450 homepage. Hum. Genom. 2009, 4, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Feyereisen, R. Insect CYP genes and P450 enzymes. In Insect Molecular Biology and Biochemistry; Elsevier: Amsterdam, The Netherlands, 2012; pp. 236–316. [Google Scholar]
- De Lima Procópio, R.E.; da Silva, I.R.; Martins, M.K.; de Azevedo, J.L.; de Araújo, J.M. Antibiotics produced by Streptomyces. Braz. J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Alder, A.; Bigler, P.; Werck-Reichhart, D.; Al-Babili, S. In vitro characterization of Synechocystis CYP120A1 revealed the first nonanimal retinoic acid hydroxylase. FEBS J. 2009, 276, 5416–5431. [Google Scholar] [CrossRef]
- Harada, H.; Shindo, K.; Iki, K.; Teraoka, A.; Okamoto, S.; Yu, F.; Hattan, J.; Utsumi, R.; Misawa, N. Efficient functional analysis system for cyanobacterial or plant cytochromes P450 involved in sesquiterpene biosynthesis. Appl. Microbiol. Biotechnol. 2011, 90, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Otomatsu, T.; Shindo, K.; Kitamura, E.; Sandmann, G.; Harada, H.; Misawa, N. Biocatalytic synthesis of flavones and hydroxyl-small molecules by recombinant Escherichia coli cells expressing the cyanobacterial CYP110E1 gene. Microb. Cell Factories 2012, 11, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, S.; Fjetland, C.R.; Lammers, P.J. Alkane-induced expression, substrate binding profile, and immunolocalization of a cytochrome P450 encoded on the nifD excision element of Anabaena 7120. BMC Microbiol 2005, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.; Werck-Reichhart, D. A P450-centric view of plant evolution. Plant J. 2011, 66, 194–211. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; DellaPenna, D. Progress in understanding the origin and functions of carotenoid hydroxylases in plants. Arch. Biochem. Biophys. 2004, 430, 22–29. [Google Scholar] [CrossRef]
- Kim, J.; Smith, J.J.; Tian, L.; DellaPenna, D. The evolution and function of carotenoid hydroxylases in Arabidopsis. Plant Cell Physiol. 2009, 50, 463–479. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2018, 47, D590–D595. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2016, 45, D200–D203. [Google Scholar] [CrossRef]
- Katoh, K.; Kuma, K.-i.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Boc, A.; Diallo, A.B.; Makarenkov, V. T-REX: A web server for inferring, validating and visualizing phylogenetic trees and networks. Nucleic Acids Res. 2012, 40, W573–W579. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2011, 40, D302–D305. [Google Scholar] [CrossRef] [PubMed]
- Ngwenya, M.; Chen, W.; Basson, A.; Shandu, J.; Yu, J.-H.; Nelson, D.; Syed, K. Blooming of unusual cytochrome P450s by tandem duplication in the pathogenic fungus Conidiobolus coronatus. Int. J. Mol. Sci. 2018, 19, 1711. [Google Scholar] [CrossRef] [Green Version]
- Akapo, O.O.; Padayachee, T.; Chen, W.; Kappo, A.P.; Yu, J.-H.; Nelson, D.R.; Syed, K. Distribution and Diversity of Cytochrome P450 Monooxygenases in the Fungal Class Tremellomycetes. Int. J. Mol. Sci. 2019, 20, 2889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, A.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Well Known for | Reference(s) |
|---|---|---|
| Acaryochloris marina | Species were isolated from the marine environment and can produce chlorophyll d as primary photosynthetic pigment that is able to use far-red light for photosynthesis. | [18] |
| Anabaena sp. WA102 | Filamentous nitrogen-fixing cyanobacterium that often form blooms in eutrophic water bodies and able to produce a range of neurotoxic secondary metabolites. | [19] |
| Synechocystis sp. PCC 6803 | Species found in fresh water and capable of both phototrophic and heterotrophic growth; owing to this ability, it was one of the most highly studied cyanobacterium for these characteristics. This species lost its nitrogen-fixing ability. | [20] |
| Synechococcus elongatus PCC 6301 | Unicellular, rod-shaped, fresh-water living, obligate photoautotrophic organism that has long been used as a model organism for photosynthesis research. | [21] |
| Synechococcus sp. WH8102 | Widely found in marine water across the world. It is well known for its oligotrophic nature, as it can utilize nitrogen and phosphorus sources. It also developed strategies to conserve limited iron stores by using nickel and cobalt in some enzymes. Species belonging to the genus Synchococcus are considered generalist compared to Prochlorococcus species, as they are nutritionally versatile and adapted to different ecological niches. These species developed a unique type of swimming motility, as they propel in the absence of any demonstrable external organelle. | [22] |
| Thermosynechococcus elongates | This species is unique among cyanobacterial species, as it grows in hot springs and has an optimal growth temperature of 55 °C. | [23] |
| Cyanobium sp. NIES-981 | This species is used for standard inhibition tests for toxicants in water, as it fulfills the criteria provided by the Organization for Economic Co-operation and Development test guidelines. | [24] |
| Dactylococcopsis salina | Gas-vacuolate cyanobacterium isolated from Solar Lake, a stratified heliothermal saline pool in Sinai. | [25] |
| Chamaesiphon minutus | It is an epiphyte of fresh water red alga Paralemanea catenata (Rhodophyta). | [26] |
| Leptolyngbya sp. NIES-3755 | Species belonging to this genus are found in various environments ranging from soil and fresh water to hypogean sites. This species was isolated from the soil at the Toyohashi University of Technology, Japan. | [27,28,29] |
| Halomicronema hongdechloris | It is the first cyanobacterium to be identified that produces chlorophyll f and is isolated from a stromatolite in the World Heritage site of Shark Bay, Western Australia. | [30,31] |
| Pseudanabaena sp. ABRG5-3 | It is a semifilamentous, non-heterocystous cyanobacterium isolated from a pond in Japan. | [32] |
| Prochlorococcus marinus subsp. marinus CCMP1375 | Among species of the Prochlorococcus genus, this cyanobacterium is extreme as it can grow at very low light levels in the ocean. Species belonging to this genus are the smallest known oxygen-evolving autotrophs and dominate the tropical and subtropical oceanic phytoplankton community. Species in this genus are adapted to different light levels in the ocean. | [33] |
| Geminocystis sp. NIES-3709 | Fresh water living cyanobacterium capable of accumulating large amounts of phycoerythrin, light-harvesting antenna proteins, compared to Geminocystis sp. NIES-3708. | [34] |
| Microcystis aeruginosa | Species belonging to this genus are the most representative of toxic bloom-forming cyanobacteria in eutrophic waters. M. aeruginosa is well-known for its toxicity by producing various toxic small polypeptides, including microcystin and cyanopeptolin. | [35] |
| Cyanobacterium sp. Strain HL-69 | It is isolated from the magnesium sulfate-dominated hypersaline Hot Lake in northern Washington. | [36] |
| Crocosphaera watsonii | Nitrogen-fixing cyanobacterium found in oligotrophic oceans adapted to iron and phosphorus limitation. | [37] |
| Crocosphaera subtropica | Unicellular cyanobacteria capable of fixing atmospheric dinitrogen (diazotroph) in marine environments, like filamentous cyanobacterial species. | [38] |
| Trichodesmium erythraeum | Filamentous cyanobacterium known as the primary producer and supplier of new nitrogen through its ability to fix atmospheric dinitrogen (diazotroph) in tropical and subtropical oceans. | [39] |
| Arthrospira (Spirulina) platensis | Economically important cyanobacterium, an important source of nutrition and medicinal value. This species is consumed as a source of protein around the world. | [40] |
| Planktothrix agardhii | Cyanobacterium forming bloom in eutrophic water and capable of producing toxins. | [41] |
| Moorea producens | Prolific secondary metabolite producing filamentous tropical marine cyanobacterium. One-fifth of its genome is devoted to the production of secondary metabolites. | [42] |
| Gloeobacter violaceus | Ancient cyanobacterium that lacks thylakoid membranes. | [43] |
| Nostoc sp. PCC 7120 | Filamentous cyanobacterium capable of fixing atmospheric dinitrogen (diazotroph). | [44] |
| Nostoc punctiforme | A facultative heterotroph symbiotic cyanobacterium capable of establishing symbiosis with Anthoceros punctatus. | [45] |
| Nostoc azollae 0708 | A nitrogen-fixing endosymbiont of water fern Azolla filiculoides Lam. | [46] |
| Anabaena sp. strain 90 | Hepatotoxic bloom-forming cyanobacterium with 5% of its genome devoted to synthesis of small peptides that are toxic to animals. | [47] |
| Calothrix strain 336/3 | Industrially relevant cyanobacterium capable of producing higher levels of hydrogen (biofuel) compared to N. punctiforme PCC 73102 and Nostoc (Anabaena) sp. strain PCC 7120. | [48] |
| Fischerella sp. NIES-3754 | Cyanobacterium isolated from hot spring in Japan with potential to have thermoresistant optogenetic tools. | [49] |
| Nodularia spumigena UHCC 0039 | Cyanobacterium responsible for Baltic sea brackish water cyanobacterial blooms producing toxins. | [50] |
| Cyanobacterial Species | Bacillus Species | Mycobacterial Species | Streptomyces Species | |
|---|---|---|---|---|
| Total No. of Species Analyzed | 114 | 128 | 60 | 48 |
| No. of P450s | 341 | 507 | 1784 | 1625 |
| No. of Families | 36 | 13 | 77 | 144 |
| No. of Subfamilies | 79 | 28 | 132 | 377 |
| Dominant P450 family | CYP110 | CYP107 | CYP125 | CYP107 |
| No. of BGCs * | 770 | 1098 | 898 | 1461 |
| Types of BGCs | 73 | 33 | 18 | 159 |
| No. of P450s Part of BGCs | 27 | 112 | 204 | 554 |
| Average No. of P450s | 3 | 4 | 30 | 34 |
| P450 Diversity Percentage | 0.09 | 0.02 | 0.07 | 0.18 |
| Average No. of BGCs | 7 | 9 | 15 | 30 |
| Gene Cluster Diversity Percentage | 0.08 | 0.02 | 0.03 | 0.23 |
| Percentage of P450s Part of BGCs | 8 | 22 | 11 | 34 |
| Reference | This work | [54] | [55,59] | [55] |
| P450 Names | Type of Clusters | Most Similar Known Cluster | Similarity |
|---|---|---|---|
| CYP213A8 | T3PKS | Xenocyloins | 25% |
| CYP213A5 | bacteriocin | ||
| CYP213A6 | T3PKS | Colicin V | 2% |
| CYP110AH1 | NRPS | Anabaenopeptin NZ 857/nostamide A | 100% |
| CYP213A3 | bacteriocin | ||
| CYP120C2 | T2PKS | Ambiguine | 6% |
| CYP110K6 | NRPS | ||
| CYP120A21 | bacteriocin | ||
| CYP110Q3 | NRPS, T1PKS | Hapalosin | 40% |
| CYP110C17 | terpene | ||
| CYP110C29 | NRPS, T1PKS | Nostophycin | 27% |
| CYP1011G1 | NRPS, T1PKS | Crocacin | 23% |
| CYP110AP1 | terpene | ||
| CYP110AT1 | NRPS, T1PKS | Hapalosin | 40% |
| CYP110Q4 | NRPS, T1PKS | Hapalosin | 40% |
| CYP110C21 | NRPS-like | Anacyclamide | 14% |
| CYP197E3 | NRPS, T1PKS | Cryptophycin | 37% |
| CYP110AG1 | terpene | Hectochlorin | 25% |
| CYP110E29 | terpene, thiopeptide, T1PKS, NRPS | Nostophycin | 27% |
| CYP110E18 | terpene, thiopeptide, T1PKS, NRPS | Nostophycin | 27% |
| CYP110C21 | terpene | ||
| CYP110Q4 | NRPS, T1PKS | Puwainaphycins | 40% |
| CYP110AT1 | NRPS, T1PKS | Puwainaphycins | 40% |
| CYP120A13 | ladderane | ||
| CYP1185A1 | lassopeptide, bacteriocin | ||
| CYP110Q2 | NRPS, T1PKS | Hapalosin | 40% |
| CYP110C14 | terpene | 6,6′-oxybis(2,4-dibromophenol) | 14% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khumalo, M.J.; Nzuza, N.; Padayachee, T.; Chen, W.; Yu, J.-H.; Nelson, D.R.; Syed, K. Comprehensive Analyses of Cytochrome P450 Monooxygenases and Secondary Metabolite Biosynthetic Gene Clusters in Cyanobacteria. Int. J. Mol. Sci. 2020, 21, 656. https://doi.org/10.3390/ijms21020656
Khumalo MJ, Nzuza N, Padayachee T, Chen W, Yu J-H, Nelson DR, Syed K. Comprehensive Analyses of Cytochrome P450 Monooxygenases and Secondary Metabolite Biosynthetic Gene Clusters in Cyanobacteria. International Journal of Molecular Sciences. 2020; 21(2):656. https://doi.org/10.3390/ijms21020656
Chicago/Turabian StyleKhumalo, Makhosazana Jabulile, Nomfundo Nzuza, Tiara Padayachee, Wanping Chen, Jae-Hyuk Yu, David R. Nelson, and Khajamohiddin Syed. 2020. "Comprehensive Analyses of Cytochrome P450 Monooxygenases and Secondary Metabolite Biosynthetic Gene Clusters in Cyanobacteria" International Journal of Molecular Sciences 21, no. 2: 656. https://doi.org/10.3390/ijms21020656
APA StyleKhumalo, M. J., Nzuza, N., Padayachee, T., Chen, W., Yu, J. -H., Nelson, D. R., & Syed, K. (2020). Comprehensive Analyses of Cytochrome P450 Monooxygenases and Secondary Metabolite Biosynthetic Gene Clusters in Cyanobacteria. International Journal of Molecular Sciences, 21(2), 656. https://doi.org/10.3390/ijms21020656